丙酸鉻的功能及其在水產飼料中的應用
2023-01-03 03:53:08劉亮李寧任麗娜
飼料博覽 2022年6期
曹 杰,劉亮,李寧,任麗娜
(建明(中國)科技有限公司,廣東 珠海 519040)
鉻(Cr)作為一種金屬元素,其原子量為51.996,密度為7.20 g·cm-3,熔點為(1 857±20)℃,沸點為672℃。在動物體內存在的主要形式為Cr、Cr2+、Cr3+、Cr6+。價態越高,其氧化性能就越高,在空氣中Cr2+很不穩定,會迅速發生氧化轉化為Cr3+;Cr6+能夠與氧結合形成具有很強氧化性的鉻酸鹽或者重鉻酸鹽;在酸性溶液中很容易發生還原反應,轉化為Cr3+;Cr3+是所有鉻元素中最穩定的一種氧化態,也是生物體中最常見的一種鉻元素存在形式。1954年發現,大鼠補鉻能夠增強其肝臟合成脂肪酸以及膽固醇的能力,研究者隨后提出鉻可能是動物體必需的一種微量元素。隨后開展試驗分離出葡萄糖耐受因子(GTF),表明其中含有鉻,從而證明鉻是影響胰島素發揮功能所必需的元素之一。GTF能夠促進受體和胰島素之間的結合,從而促進胰島素對動物體內糖(碳水化合物)的代謝能力,同時還證明鉻是胰島素能夠促進脂肪代謝所必需的物質[1]。
添加在飼料中的鉻有兩種形式:無機鉻和有機鉻,鉻的吸收與其價態及化學形式有很大關系,無機鉻在動物體內的吸收率非常有限,為0.4%~3%;有機鉻的吸收效率更高,可達到25%~30%,并且有機鉻的生物學效應更高。研究表明丙酸鉻的吸收效率比其他的有機鉻更高[2],這是由于有機微量元素吸收率的高低取決于其配體的種類和螯合程度,如圖1,美國食品藥品監督管理局(FDA)認定的丙酸鉻分子式為[Cr3(O)(CH3CH2CO2)6(H2O)3]CH3CH2CO2,分子量為737.534,這種由3個鉻跟7個丙酸螯合形成復雜螯合結構的丙酸鉻,其分子量大、螯合率高,可避免復雜環境和微生物的影響,并且減少了與其他微量元素之間的拮抗和干擾,在小腸中釋放出來后到血液中發揮作用,不會在機體中富集。
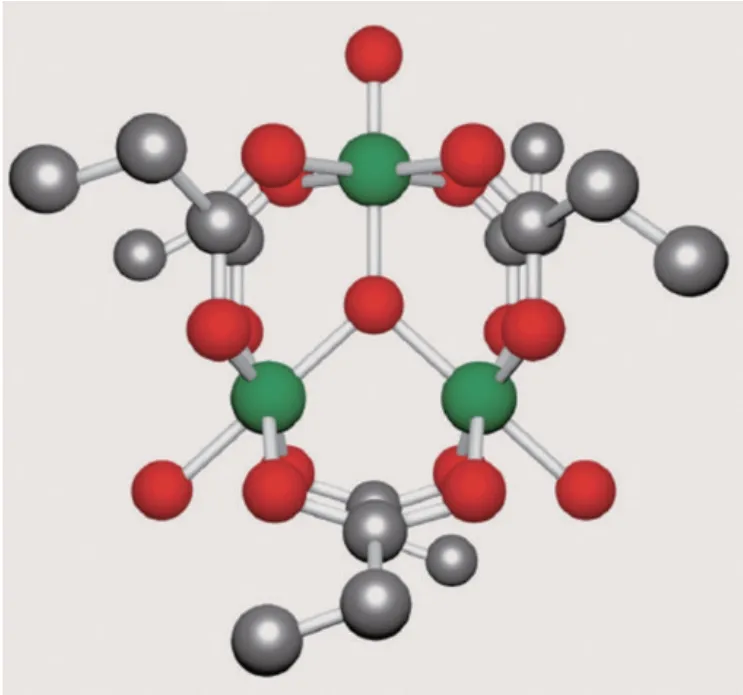
圖1 丙酸鉻獨特的分子式結構
1 丙酸鉻的生物學功能
丙酸鉻的生物學功能是能夠通過Cr3+構成GTF來增強胰島素的功能,從而對動物體的糖脂代謝、蛋白質及核酸代謝產生影響[3]。
1.1 丙酸鉻對糖代謝的影響
丙酸鉻是組成GTF的重要成分,能夠促進受體和胰島素之間的結合,而且作為低分子量鉻結合物(LMWCr)中的Cr3+可擴增胰島素的信號,增強胰島素的功能,進而促進葡萄糖向細胞膜的輸送、轉運效率,降低動物的血糖濃度[4]。J.Heimbach等[5]研究指出,鉻對于胰島素功能的改善可能有兩種途徑:(1)激活受體細胞膜與胰島素間的二硫鍵,進而增強特異變體與胰島素之間的結合力;(2)鉻的存在使細胞膜表面的胰島素受體數量增多,因此組織對胰島素的敏感度增強,提高組織細胞攝取葡萄糖的效率,脂肪組織吸收的葡萄糖效率增加,減少體內脂肪分解[6]。
體外分子試驗結果表明,細胞內存在的Cr3+能提升胰島素受體上酪氨酸殘基磷酸化水平,增強胰島素信號傳遞到下游的效果[7];蛋白免疫印跡技術研究發現Cr3+也能夠加速葡萄糖轉運蛋白4(GLUT4)的轉膜速率,增加機體對葡萄糖的攝入[8];熒光定量技術研究發現鉻能夠增強細胞胰島素受體IR、葡萄糖轉運基因GLUT等的表達水平,能夠加快血糖在體內的轉移效率[9]。也有報道發現丙酸鉻通過增強糖原合成酶的活性來提高糖原合成。將丙酸鉻外源加入到胰島素含量低的犢牛日糧中,能夠使犢牛的血糖代謝速率維持穩定,這也證明了丙酸鉻在犢牛上能提高其血糖代謝速率[10]。
1.2 丙酸鉻對脂肪代謝的影響
在脂肪的體內代謝過程中,丙酸鉻起的主要功能是調節膽固醇平衡,從而影響動物肝臟中膽固醇和脂肪的清除和合成[11]。促進脂肪的重新分配,降低體內總膽固醇以及血清三酰甘油的含量,提高動物體內血清中高密度脂蛋白膽固醇的含量,從而降低脂肪在體內的沉積[12]。丙酸鉻能夠消除在主動脈上已經沉積的膽固醇,同時還有助于減少沉積的膽固醇數量,因此有助于預防動脈粥樣硬化的發生,增強動物體脂肪組織的脂肪分解代謝并顯著抑制脂肪合成代謝[13]。
研究表明,丙酸鉻影響動物脂肪代謝的因素主要包括:(1)增強胰島素功能和活性,調節和影響脂類代謝,從而使高血脂狀況得到緩解;(2)增強肝臟內皮細胞酶(HEL)以及血漿卵磷脂膽固醇-酰基轉移酶(LCAT)的活性,增強骨骼肌和心臟脂肪等組織上脂蛋白酶(LPL)的功能,促進動物體生成更多高密度脂蛋白(HDL);(3)通過對體內膽固醇代謝的影響以及調節各種脂蛋白的比例和含量,進而對脂肪蓄積進行調節和改善[14]。Wang M.Q.等[15]向豬日糧中加入200 μg·kg-1的丙酸鉻復合制劑,發現豬血清脂肪酶活性和HDL的濃度顯著提高,并且血清中的膽固醇含量和三酰甘油含量顯著降低。
1.3 丙酸鉻對蛋白質及核酸代謝的影響
胰島素功能被丙酸鉻增強以后,氨基酸進入胞內的效率同時被促進,進而增強蛋白質轉錄和合成。此外,丙酸鉻有助于血液中的胰島素運轉到周圍組織,肌肉細胞增強其對胰島素的內化作用,加快葡萄糖吸收,促進組織貯存更多的氨基酸(如蛋氨酸、絲氨酸以及甘氨酸),加強肝臟細胞利用上述氨基酸合成蛋白質的能力[16]。研究發現,雞補充丙酸鉻能提高其日糧中蛋白質利用率,同時降低氮的排泄率[17]。
研究證明,丙酸鉻影響蛋白質代謝的可能機制有:(1)啟動并激活受體細胞膜和胰島素間的二硫鍵連接,促進葡萄糖和氨基酸等營養物質進入胞內,在生長激素和類胰島素樣生長因子等多因子的作用下促進動物體內的蛋白質合成代謝;(2)丙酸鉻進入體內后與細胞染色體核仁的DNA相結合,增加DNA啟動點數量,促進蛋白質的合成[18]。
核酸中鉻含量較高時,有助于核酸結構的完整性和穩定性維持。在鉻的保護下,RNA能免受熱變性,同時鉻主要在核仁中積累,能促進RNA合成后的后加工過程,再加工為rRNA[19]。
1.4 丙酸鉻對免疫功能的影響
通過改善和調節機體pH,丙酸鉻能維持血液中滲透壓的穩定,平衡血液中膽固醇水平,從而保證動物體的生理功能得以正常發揮。皮質醇是應激反應中腎上腺產生的一種激素,對體內抗體產生抑制作用,同時對淋巴細胞功能也有抑制,進而對免疫過程的多環節產生抑制。研究證實,補充丙酸鉻能提高動物應激時血清中免疫球蛋白水平,降低血清皮質醇濃度,提高抗體效價[20]。在山羊日糧中補充丙酸鉻0.5 mg·d-1,其淋巴細胞數和白細胞數等百分比增多,同時增快了外周血淋巴細胞的增殖速度[21]。運輸應激狀況下,丙酸鉻的添加可使公牛血液中中性粒細胞百分率和白細胞總數增加[22]。在虹鱒日糧中添加丙酸鉻使頭腎巨噬細胞的溶菌酶含量、吞噬能力和呼吸爆發活力得以提高[23]。
2 丙酸鉻在水產飼料中的應用
作為營養素,丙酸鉻最早在畜禽方面進行研究,而作為飼料添加劑,丙酸鉻當前的適用范圍包括豬、奶牛、家禽。研究表明在畜禽上使用丙酸鉻可促進其生長,尤其是促進糖類等營養物質的利用,同時能改善動物的體成分組成,并且提高動物的抗應激能力和增強免疫能力[14,24-27]。近年丙酸鉻在水產養殖上的研究也取得了一些成果,特別是在魚類上的研究。研究表明,丙酸鉻能改變糖類等營養物質的沉積和利用方向,對水產動物的免疫、繁殖、生長以及胴體品質產生影響,可降低應激反應,強化動物體的免疫功能,提高生長和生產性能。目前已證實丙酸鉻等有機鉻可以促進鯉魚[28]、草魚[29]、鯽魚[30]、羅非魚[31]、大黃魚[32]、武昌魚[33]、牙鲆[34]、團頭魴[35]、虹鱒[23]、三文魚[36]、河蟹[37]和凡納濱對蝦[38]等水產動物的生長和飼料利用率。
2.1 提升水產動物糖利用率
糖(碳水化合物)是飼料原料中最廉價的能量物質,如能使用糖代替部分飼料中的蛋白質,則有節約蛋白質的效果,從而可以降低飼料中蛋白質的含量,不僅節約配方成本,而且能對蛋白質和漁業資源形成有效保護,因為該舉措能降低養殖過程中氨氮、亞硝酸鹽的排放,減少對水體環境的污染。
但水產動物對糖的利用能力低,被認為是“葡萄糖不耐受”體質。當配方中糖含量過高時,有可能引發機體持續的高血糖,從而使其代謝、生理功能發生紊亂[39]。研究發現,Cr3+可促進水產動物對不同種類糖的利用效率,增強水產動物對葡萄糖的耐受性[40],能抑制糖異生并提高鯉魚對葡萄糖的利用效率[41]。在草魚飼料中添加不同濃度梯度的丙酸鉻,隨著丙酸鉻劑量的增加,草魚的蛋白質效率(PER)、特定生產率(SGR)也隨之升高,0.4 mg·kg-1劑量時PER和SGR達到最高值,此時的飼料系數卻是最小值;血清總膽固醇(TC)的含量隨丙酸鉻劑量的增加而降低(0.8 mg·kg-1添加水平時達到最低值);此外,0.8 mg·kg-1劑量時草魚的肥滿度顯著增高,內臟指數降低[42]。在三種不同種類的糖(葡萄糖、糊精和淀粉)作為糖源的鯽魚飼料中添加Cr3+,發現葡萄糖組的血糖濃度顯著降低,淀粉組的血糖濃度也受到一些影響[30]。
在用淀粉和葡萄糖作為糖源的羅非魚飼料中添加不同來源的鉻,發現Cr3+會抑制糖異生過程中葡萄糖-6-磷酸酶的活性,而糖酵解途徑中磷酸果糖激酶的活性在葡萄糖組中顯著增加。同時肝臟糖原的含量顯著增高,這表明Cr3+能促進動物更好地合成糖原,并延緩葡萄糖吸收高峰的來臨[43]。在含淀粉和葡萄糖的日糧中添加有機鉻,并連續8周投喂奧尼羅非魚魚種,結果表明,有機鉻組的特定生長率(SGR)、飼料效率(FCR)都顯著優于對照組,說明在羅非魚上使用有機鉻能顯著改善其對葡萄糖的利用[31]。在奧尼羅非魚日糧中添加不同濃度的煙酸鉻,然后灌服葡萄糖并檢測其血糖含量,結果煙酸鉻組血糖濃度的升高被顯著抑制了[44]。使用分子生物學技術手段研究添加酵母鉻(Cr-Y)、吡啶羧酸鉻(Cr-Pic)在尼羅羅非魚飼料中的作用,發現兩者均能降低血糖,影響體組織中營養物質構成,并有提高魚體生長的趨勢。此外,有機鉻能顯著增強尼羅羅非魚葡萄糖轉運基因GLUT的表達[45]。
2.2 促進水產動物生長
飼料中添加丙酸鉻可提高魚的生長速度,降低餌料系數。用含鉻的飼料喂養1齡鯉魚65 d發現鉻飼喂組的魚糞粗蛋白為12.79%,粗脂肪為2.01%,均低于對照組(對照組魚糞的粗蛋白和粗脂肪含量分別為15.60%和2.48%)。3月齡鯉魚飼喂30 d的研究結果與之類似,說明有機鉻可改善鯉魚對蛋白質、脂肪等營養物質的消化吸收,進而促進鯉魚的生長[28]。對飼喂高葡萄糖(35%)飼料的鯉魚,Cr3+在促進生長、提高飼料利用以及糖利用能力方面具有明顯的效果。添加吡啶羧酸鉻(CrPic)和蛋氨酸鉻(CrMet)能顯著提高鯉的增重率(WGR)、特定生長率(SGR)、飼料效率(FCR)以及蛋白質效率(PER);顯著增加全魚脂肪含量;顯著提高肌糖原含量;CrPic、CrMet顯著降低肝胰臟糖異生途徑磷酸烯醇式丙酮酸激酶(PEPCK)活性。同時CrMet組的肝糖原顯著提高;CrMet還顯著降低血清三酰甘油和總膽固醇含量;添加CrMet還顯著提高魚體胰島素、胰島素受體、生長激素含量;顯著降低血清葡萄糖和皮質醇含量;顯著提高肝胰臟糖酵解途徑丙酮酸激酶(PK)、己糖激酶(HK)活性[46]。在1齡鯉飼料中添加0.5 mg·kg-1鉻飼喂鯉魚65 d后,其相對生長率(RGR)比對照組高13.79%,飼料轉化率(FCR)改善21.83%[47]。
飼料中添加吡啶羧酸鉻可提高奧尼羅非魚的生長和飼料利用率[31]。在團頭魴的飼料中添加不同水平吡啶甲酸鉻表明:隨著添加量的增加,團頭魴的尾增重率(TWGR)、瞬間生長率(IGR)、絕對生長率(AGR)和相對生長率(RGR)等指標均增長,餌料系數(FCR)則顯著下降[35]。煙酸鉻、蛋氨酸鉻和甘氨酸鉻添加量為1.7 mg·kg-1均能提高奧尼羅非魚的長速并且改善飼料系數FCR[48]。凡納濱對蝦飼料中添加1.6 mg·kg-1的吡啶甲酸鉻能顯著提升其成活率SR、特定生長率SGR以及增重率WGR,而餌料系數FCR顯著降低[49]。
2.3 提高水產動物抗應激能力和免疫功能
羅非魚飼料中添加Cr3+后能減少血漿中皮質醇的濃度,緩解應激[50]。丙酸鉻能明顯提高羅非魚的抗應激能力,通過模擬羅非魚運輸過程中應激指標的變化,發現對照組血清肌酸激酶(CK)活性明顯提高,血糖濃度也明顯提高;但隨著外源添加丙酸鉻,各羅非魚試驗組的血糖濃度增幅均顯著低于對照組;模擬研究結束后,各組的肌酸激酶(CK)和血糖濃度的增量均呈下降趨勢,羅非魚對照組血清中肌酸激酶(CK)的活性比模擬運輸前增加了20.31 U·L-1,而添加200 μg·kg-1丙酸鉻的試驗組只增加了8.47 U·L-1,說明在應激條件下使用丙酸鉻能夠顯著改善機體的抗應激力[51]。在鯉魚飼料中添加酵母鉻,顯著提高了鯉魚運輸后的成活率[52]。李洪霞等[53]發現添加有機鉻可以顯著提升羅非魚的生長,改善飼料利用率(FCR)和抗冷應激能力。在受熱應激刺激下的建鯉飼料中添加有機鉻,增強了建鯉的抗熱應激能力,并且熱應激狀況下建鯉的PER、WGR以及肌肉粗蛋白含量均有提高[54]。在虹鱒飼料中按照1 540、2 340和4 110 μg·kg-1的濃度添加酵母鉻,發現虹鱒幼魚血清中溶菌酶含量以及呼吸爆發水平顯著提高,并且吞噬細胞的吞噬作用得到增強[23]。適量添加鉻可顯著提高凡納濱對蝦血清中酚氧化物酶、超氧化物歧化酶、堿性磷酸酶、谷丙轉氨酶和谷草轉氨酶的活性[49]。
吉富羅非魚飼糧中添加有機鉻0~0.8 mg·kg-1時,隨著添加濃度的提升,肝臟的堿性磷酸酶活性顯著增加;而當有機鉻的添加量提升到1.2~10.0 mg·kg-1時,肝臟的堿性磷酸酶活性卻顯著下降;并且外源添加鉻的量為0.8 mg·kg-1時肝臟溶菌酶活性顯著上升;外源鉻添加濃度為1.2 mg·kg-1時堿性磷酸酶活性顯著上升[55]。SRBC作為抗原接種奧尼羅非魚,并在其日糧中添加Cr3+發現,添加鉻試驗組的羅非魚的抗SRBC抗體效價在初次和二次免疫后都顯著上升,而且能維持較長時間[56]。
3 丙酸鉻對水體環境的影響
魚體中的鉻主要以Cr3+和Cr6+形式存在,Cr3+能加強糖代謝,促進動物生長,提高免疫力和抗應激能力。Cr3+對水體環境不會造成影響,但過量添加Cr3+對水生動物生長卻有負作用。
據報道,Cr3+的促生長劑量在鯉魚飼料中的應用濃度應低于2 mg·kg-1。當使用量為2 mg·kg-1時,30 d時鯉魚的生長速度減緩,40 d時甚至會發生負生長;當使用量為5 mg·kg-1時,生長速度減緩和負生長的情況分別縮減至20 d和30 d時發生[47]。水中總鉻濃度為24 μg·L-1和54 μg·L-1時,大鱗大馬哈幼魚(Oncorhynchus tshawytscha)暴露105 d,其成活率、生長速度都未受影響,繼續將水中總鉻濃度提高到120 μg·L-1以上,大鱗大馬哈幼魚的成活率和生長速度都明顯下降。腎臟是鉻作用的靶器官,魚的腎臟暴露在高濃度的鉻環境中,肉眼和顯微鏡下能夠觀察到病變的發生(如腎小管內壁細胞壞死),脂質過氧化產物上升。血液中DNA的變異也與脾腎的病理變化有關。這些研究表明鉻積累并最終影響脂質過氧化途徑,導致DNA損傷(表現為染色體變化),最終使組織損傷和細胞死亡[57]。無機鉻作為普遍存在于水體中的一種污染物,是導致我國地下水和土壤受污染的重要原因。其中Cr6+具有潛在的致癌性,會對水體環境造成污染。水體中Cr6+排放量最多的是生產含鉻(Ⅵ)廢物的企業,通過管道將含鉻(Ⅵ)廢水排放到水環境中是導致鉻殘留和重金屬超標的主要原因。在我國,由含鉻(Ⅵ)廢物排放造成的地下水和土壤受污染的比例高達20%。排放物中含有Cr6+廢物的企業數量眾多,這些含鉻(Ⅵ)排放物會影響海洋、河口、江河以及湖泊中重金屬鉻殘留,而且在水生生物中也存在鉻殘留現象。
細胞內大分子與滲入細胞后的Cr6+相互結合,使遺傳密碼改變,從而使細胞發生突變和癌變,具有潛在的致癌性[58]。研究表明,Cr6+危害金魚的主要原因是對肝細胞產生毒性,損害肝細胞,該毒性的強弱依賴鉻濃度刺激下產生活性氧的多少[59]。Cr6+對虹鱒的油脂過氧化反應、細胞形態、超氧化物歧化酶(SOD)活性、金屬硫蛋白(MT)水平和生長速度等都有影響。機體中肝臟組織對適應Cr6+毒性的方面起重要的作用,鰓卻對Cr6+的毒性比較敏感[60]。Cr6+劑量較低時對草魚有誘導效應,能夠短暫的、不同程度地影響肝臟谷胱甘肽過氧化物酶(GSH-Px)和超氧化物歧化酶(SOD)等抗氧化物質的活性;隨著Cr6+劑量的增大,GSHPx和SOD的活性變低,動物體抗氧化系統功能受到損傷[61]。研究表明,動物體內Cr6+轉化為Cr3+發揮毒理作用是通過在機體內形成活性氧,并引發油脂發生過氧化反應,同時增加抗氧化酶系的活性,進而激活氧化脅迫應答的適應性反應[62]。
對象山港重點養殖區內重金屬含量的調查表明,水體中總鉻的含量很少,達到一類海水水質標準,但是主港區和鄰近分區水體的鉻含量較高。同時發現東海10個重點河口港灣中象山港的重金屬數值是偏高的[63]。東海隘頑灣海域的4個養殖區中,水體表層沉積物的總鉻含量平均為82.97 μg·L-1,區間值為64.46~105.88 μg·g-1,各區的含鉻量差異顯著[64]。武漢墨水湖中鉻濃度為1.71 μg·L-1,湖中鰱魚的鉻含量為0.353 μg·g-1,但湖底沉積物的重金屬污染嚴重,總鉻濃度為213 mg·kg-1,是42 mg·kg-1的環境背景和生態效應臨界值(TEL值)的5倍,是160 mg·kg-1生態效應必然濃度值(PEL值)的1.3倍[65]。
4 總結
鉻作為人和動物必需微量元素,也能促進水產動物的生長,改善免疫功能和提升抗應激能力。但肉食性魚類(鱖鱸等)對淀粉等糖類耐受性差,淀粉等糖類使用量過高時會影響TOR通路,從而引起厭食、肝臟脂肪蓄積和生長的減緩等。而丙酸鉻能提高糖利用率,增強組織對葡萄糖的吸收和轉運效率;同時能夠增強脂肪分解并顯著地抑制脂肪合成,從而降低脂肪在機體中的沉積。這些研究對提高糖類在水產動物特別是肉食性魚類飼料配方中的占比、降低魚粉用量具有重要的啟發和價值。但需挖掘其作用機制,關注其在不同品種和不同水體中的使用方式和安全濃度。
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:10
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
