紅花變豆菜葉綠體基因組組裝與序列特征分析研究
2022-11-18 02:46:06任偉超張美琦劉秀波
中草藥 2022年22期
關鍵詞:植物
王 震,柳 馳,任偉超,張美琦,劉秀波*,馬 偉, 3, 4*
? 藥材與資源 ?
紅花變豆菜葉綠體基因組組裝與序列特征分析研究
王 震1,柳 馳2#,任偉超1,張美琦1,劉秀波1*,馬 偉1, 3, 4*
1. 黑龍江中醫藥大學藥學院,黑龍江 哈爾濱 150040 2. 開姆尼茨工業大學信息學院,德國 開姆尼茨 09111 3. 江蘇康緣藥業股份有限公司,江蘇 連云港 222001 4. 中藥制藥過程新技術國家重點實驗室,江蘇 連云港 222001
以紅花變豆菜新鮮葉片為試驗材料,通過高通量測序手段,對其葉綠體基因組進行組裝和序列特征分析,為進一步開展變豆菜屬植物的進化和遺傳發育提供指導方法。實驗流程按照Illumina公司提供的標準操作流程執行,包括樣品質量檢測、文庫構建、文庫質量檢測和文庫測序等流程,使用已報道的變豆菜的葉綠體基因組作為參考序列,得到紅花變豆菜完整的葉綠體基因組序列并對其進行組裝、特征序列分析和進化分析。紅花變豆菜葉綠體基因組具有典型的環狀四分體結構,長度155 700 bp,包括1個大的單拷貝區(large single copy,LSC,85 979 bp),1個小的單拷貝區(small single copy,SSC,17 053 bp),1對相反的序列(inverted repeats sequence,IR,26 333 bp);共注釋到130個基因,其中蛋白編碼的基因86個,tRNA基因36個,rRNA基因8個,AT含量占葉綠體基因組的61.83%;共檢測到1個正向長重復序列(forward repeat sequence),3個回文長重復序列(palindromic repeat sequence);此外,還檢測到了168個簡單重復序列(simple sequence repeats,SSR)位點;與蛋白編碼基因對應的密碼子偏好使用A/T堿基,編碼異亮氨酸的密碼子(I)使用次數最高,編碼半胱氨酸的密碼子(C)使用次數最低;最后與7種已報道的傘形科植物葉綠體基因組和2個外類群物種通過最大似然法(maximum likelihood,ML)構建系統發育樹,發現紅花變豆菜與變豆菜和直刺變豆菜親緣關系最為密切。紅花變豆菜的葉綠體基因組數據為傘形科植物研究提供了更為詳細完善的資料,為該物種葉綠體基因工程和系統進化分析提供參考依據。
紅花變豆菜;葉綠體基因組;高通量測序;系統進化;最大似然法
紅花變豆菜Fr. Schmidt是傘形科(Apiaceae)變豆菜屬多年生草本植物,是一種可食用的山野菜,具有一定的藥用價值,變豆菜屬全世界約40余種[1]。其生長在山間林下,陰濕及腐殖質較多的地方,海拔200~470 m,主要產地在我國的東北地區,另外,蒙古、朝鮮、日本北部也有分布[2]。變豆菜屬植物在中國約有18種,具有一定的經濟價值和藥用價值[3],薄片變豆菜Hance又叫血經草,具有治療跌打損傷和風寒感冒等功效[4],天藍變豆菜Franch.、直刺變豆菜S. Moor和川滇變豆菜Wolff ex Kretsch均為有名的地方藥材,具有活血化瘀和止咳化痰的藥效,同屬的紅花變豆菜也可能會具有相應的藥用價值[5]。變豆菜屬植物主要基于傳統的形態學進行鑒定,如從花粉形態[6]和果實微形態出發進行分類,但是部分品種只從形態學鑒定很難區分,導致其傳統分類發展緩慢,因此需要尋找新的方法來對其鑒定手段進行補充。
葉綠體普遍存在于陸地植物、藻類和部分原生生物當中,是綠色植物進行光合作用的細胞器[7],功能包括產生色素、合成糖和某些氨基酸等[8]。被子植物中,葉綠體基因組相對保守,絕大多數為雙鏈環狀結構,包括1個小的單拷貝區(small single copy,SSC)、1個大的單拷貝區(large single copy,LSC)以及2個編碼相同,方向相反的序列(inverted repeats sequence,IR)IRa和IRb[9]。相對于核基因組,葉綠體基因組具有結構保守、堿基變異速率適中、易于測序等多種優勢,已廣泛應用于各植物類群的系統進化研究[10]。基于本課題組前期對紅花變豆菜葉綠體基因組的研究[11],對其序列數據進一步挖掘分析,對組裝后的葉綠體基因組結構、長重復序列分析、簡單重復序列(simple sequence repeat,SSR)位點、密碼子偏好性以及與其他傘形科植物使用最大似然法(maximum likelihood,ML)構建系統發育樹,分析近緣物種親緣關系,為紅花變豆菜的鑒定、系統發育研究提供新的方法。
1 材料與方法
1.1 材料
新鮮的紅花變豆菜樣本于2020年8月20日采集自中國黑龍江省伊春市(N47°81′08′′,E128°90′97′′)。樣品保存在黑龍江中醫藥大學,樣品標本號為YCL20190507007,由黑龍江中醫藥大學馬偉研究員鑒定為傘形科變豆菜屬植物紅花變豆菜Fr. Schmidt。將采集的紅花變豆菜新鮮幼嫩的葉片,通過液氮速凍,并進行研磨,于?80 ℃冰箱保存備用。
1.2 方法
1.2.1 DNA提取 使用CTAB法提取紅花變豆菜的全基因組DNA。
1.2.2 測序 總DNA樣品經武漢貝納科技服務有限公司檢測合格后,用機械打斷的方法(超聲波)將DNA片段化,然后對片段化的DNA進行片段純化、末端修復、3′端加A、連接測序接頭,使用瓊脂糖凝膠電泳進行片段大小選擇,進行PCR擴增形成測序文(NEBNext?Ultra? DNA Library Prep Kit for Illumina?),建好的文庫先進行文庫質檢,質檢合格的文庫用Illumina NovaSeq進行測序,對測序得到的序列,通過SOAPnuke(version:1.3.0)軟件進行低質量數據過濾,過濾標準為去除N堿基含量超過5%的reads、去除低質量(質量值≤5)堿基數目達到50%的reads和去除有adapter污染的reads,最終得到有效數據(clean reads)。
1.2.3 葉綠體基因組組裝 使用SPAdes(version:3.13.0;參數:?k 127)[12]軟件進行基因組拼接,選用NCBI數據庫中近緣參考植物變豆菜葉綠體基因組(MK208987.1)與拼接結果進blastn(version:BLAST 2.2.30+;參數:?evalue 1×10?5)比對,后續使用Gapcloser(Version:1.12)對gap(含N序列)進行補洞。
1.2.4 葉綠體基因組特征分析 利用專門針對葉綠體的注釋軟件CPGAVAS2(http://47.96.249.172: 16019/analyzer/annotate)[13]進行基因注釋并繪圖紅花變豆菜葉綠體基因組物理圖;使用VMATCH(http://www.vmatch.de/)軟件(參數:minimal repeat size 30 bp)查找葉綠體基因組中的散在長重復序列片段;采用MISA軟件[版本:1.0;默認參數;對應的各個重復單元(unit size)的最少重復次數分別為1-8、2-4、3-4、4-3、5-3、6-3]對葉綠體進行SSR檢測;使用CodonW(Version:1.4.4)對密碼子偏好性進行分析,統計估算相對同義密碼子的使用頻率;通過NCBI數據庫下載6種傘形科植物,包括變豆菜L.(MK208987)、明日葉L.(MW125613)、隔山香L.(MT501096)、遼藁本L.(MN652885)、防風L.(MN857472)、白芷L.(KT963037)以及2種非傘形科植物玉米L.(NC001666)和擬南芥L.(NC000932)的葉綠體基因組序列,使用MAFFT[14]軟件進行多序列比對,最后使用MEGA X[14]軟件基于ML法構建系統發育樹。
2 結果與分析
2.1 紅花變豆菜葉綠體基因組測序及結構解析
基于課題組已報道的研究發現,紅花變豆菜的葉綠體基因組大小為155 700 bp,具有典型的環狀四分體結構:包括1個LCS區(85 979 bp),1個SCC區(17 053 bp)和2個反向互補的IRs區(26 333 bp),GC含量為61.83%[14]。
后續通過對紅花變豆菜葉綠體基因組物理圖譜的繪制(圖1)及進一步統計分析發現,測序有效數據Q20值為97.41%、Q30值為92.84%,根據比對覆蓋度發現變豆菜(MK208987.1)為最優參考序列,其覆蓋度為96.44%,不含有gap。IRs、LSC和SSC 4個區域的GC值都存在一定的差異,IRs區域的CG值最高,為42.92%;SSC區域的CG值最低,為32.56%,LCS區域的CG值介于二者之間,為36.39%(表1)。
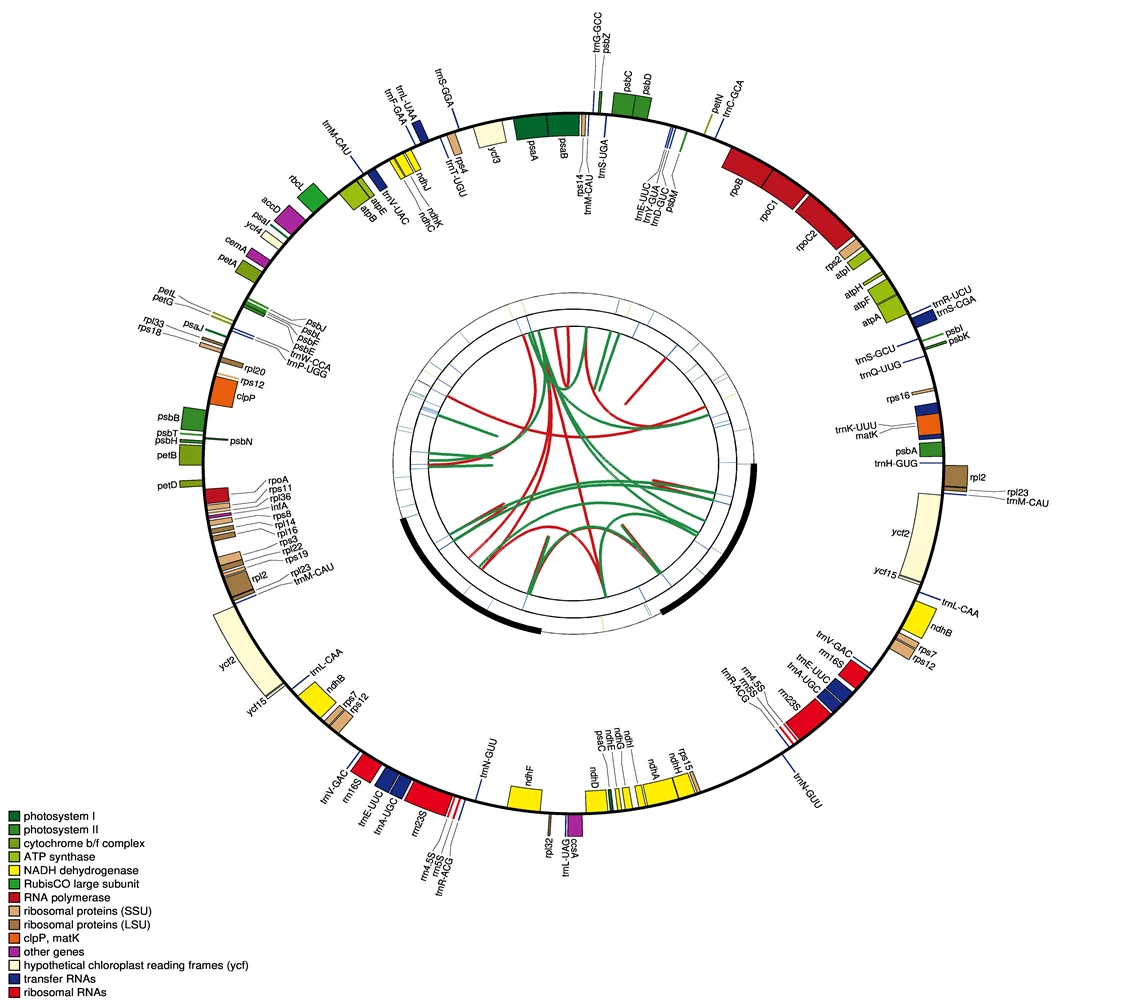
葉綠體基因組圖譜上4個環。從中心向外,第1圈顯示紅弧和綠弧分別連接的正向重復和反向重復;第2圈顯示用短杠標記的串聯重復;第3圈為簡單重復序列SSR;第4圈顯示質體上的基因結構;基因的顏色是根據他們的功能分類來區分
2.2 葉綠體基因組結構基本特征分析
基于課題組已報道的研究發現,紅花變豆菜葉綠體基因組共成功注釋了130個基因,其中蛋白編碼的基因86個,tRNA基因36個,rRNA基因8個[11]。
通過對以上結果進一步的統計分析發現,根據其基因編碼功能可以分為4類(表2),第1類是與自我復制有關的基因,分為5個組,分別是tRNA基因、核糖體小亞基、核糖體大亞基、RNA聚合酶亞基和核糖體RNA基因,成員數量分別為26、12、9、4和4個;第2類為與光合作用相關的基因,細分為7組,分別是光系統I、光系統II、ATP合成亞基、細胞色素亞基、二磷酸核酮糖羧化酶大亞基、NADPH乳酸脫氫酶亞基和依賴ATP蛋白酶P基因亞基,成員數量分別為5、15、6、6、1、11和1個;第3類為其他功能基因,包括成熟酶、外膜蛋白基因、C型細胞色素合成基因和乙酰輔酶A羧化酶亞基各1個;第4類為4個未知功能的基因,還需要進一步研究,以確定其功能。
2.3 長重復序列分析
所有的長重復序列包括3種類型,包括正向重復序列(forward repeat sequence)、回文重復序列(palindromic repeat sequence)和串聯重復序列(tandem repeat sequence),這些長重復序列可能具有促進葉綠體基因組重排的功能,并且可以增加其居群遺傳多樣性[16]。在紅花變豆菜葉綠體基因組中共發現了1個正向重復序列,3個回文重復序列,并未發現串聯重復序列。其中1個正向重復序列和2個回文重復序列非常短,分別為34、34、30 bp,1個回文重復序列非常長,為26 333 bp,是葉綠體基因組的IR區域。利用這些重復序列,可以為以后開發種群進化標記研究提供基礎。
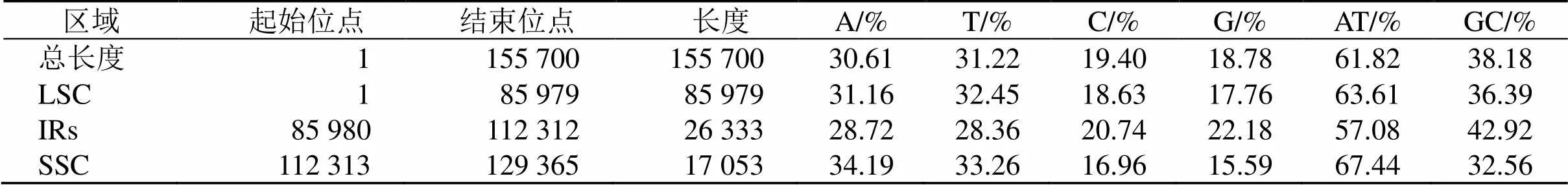
表1 紅變豆菜葉綠體基因組堿基組成
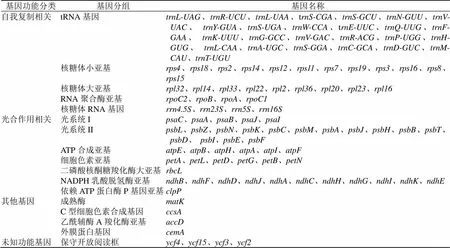
表2 紅花變豆菜葉綠體基因組注釋基因
2.4 SSR位點分析
SSR又稱短串聯重復標記、微衛星序列標記[17-18]。SSR表示由基因組中1~6個核苷酸組成的基本單位重復多次構成的一段DNA序列,廣泛存在于基因組的各個區域,且側翼序列通常都是保守性較強的單一序列,可用于個體或物種之間的多態性研究[19]。通過對紅花變豆菜葉綠體基因組的分析,共發現了168個SSR位點,分為4個類型,包括復雜重復類型112個,其數量最多;3個堿基重復類型2個,其數量最少;2個堿基重復類型47個;4個堿基重復類型7個,沒有單堿基重復類型。97.60%的SSR位點都含有A/T的堿基,僅有4個SSR位點由G/C堿基組成,說明A/T堿基具有堿基偏好性,這可能跟紅花變豆菜葉綠體基因組中A/T堿基含量占比高(61.82%)有關聯,造成這種偏好性的原因可能與解鏈難易程度有關。
2.5 密碼子偏好性分析
紅花變豆菜葉綠體基因組中86個蛋白由21 963個密碼子共同編碼(表3)。由AUU編碼的異亮氨酸數量最多,為876個;由UGC編碼的半光氨酸數量最少,僅有60個。3種終止密碼子UAA、UAG和UGA在紅花變豆菜的葉綠體基因組中使用,其在同義密碼子中的占比為50%、25%和25%。通過對86個蛋白編碼基因序列分析,得出了同義密碼子相對使用度(relative synonymous codon usage,RSCU)[20],RSCU是對同義密碼子使用偏好的評估,通過統計發現,由UUA編碼的亮氨酸使用最頻繁,偏好性最大;由AGC編碼的絲氨酸使用頻率最低,偏好性最小;由UGG編碼的色氨酸和由AUG編碼的甲硫氨酸無偏好性。
2.6 紅花變豆菜與其他7種傘形科植物葉綠體基因組的比較分析
本研究基于近些年報道的直刺變豆菜[21]、變豆菜[22]、明日葉[23]、隔山香[24]、遼藁本[25]、防風[26]和白芷[27]共7種傘形科植物與紅花變豆菜的葉綠體序列長度、編碼基因數量及組成結構進行比較(表4)。經過數據統計分析,傘形科內8種植物葉綠體基因組的長度為146 918~155 919 bp,并且都是由典型的四分體結構組成;其大拷貝區和小拷貝區的序列長度差異不大;除了遼藁本的反向重復序列長度為明日葉和防風的2倍,其他7種反向重復序列長度差異也不大;從編碼的蛋白質數量來看,紅花變豆菜編碼的蛋白質數量最多,可能其葉綠體在生長過程中能行使更多的功能。紅花變豆菜相比與變豆菜與直刺變豆菜相比,編碼的蛋白最多和SSC區序列最長。除此之外,這8種傘形科植物的葉綠體基因組基因順序和結構與大多數已報道的被子植物葉綠體基因組相似,這說明在植物進化過程中,葉綠體基因序列具有高度的保守性[28]。
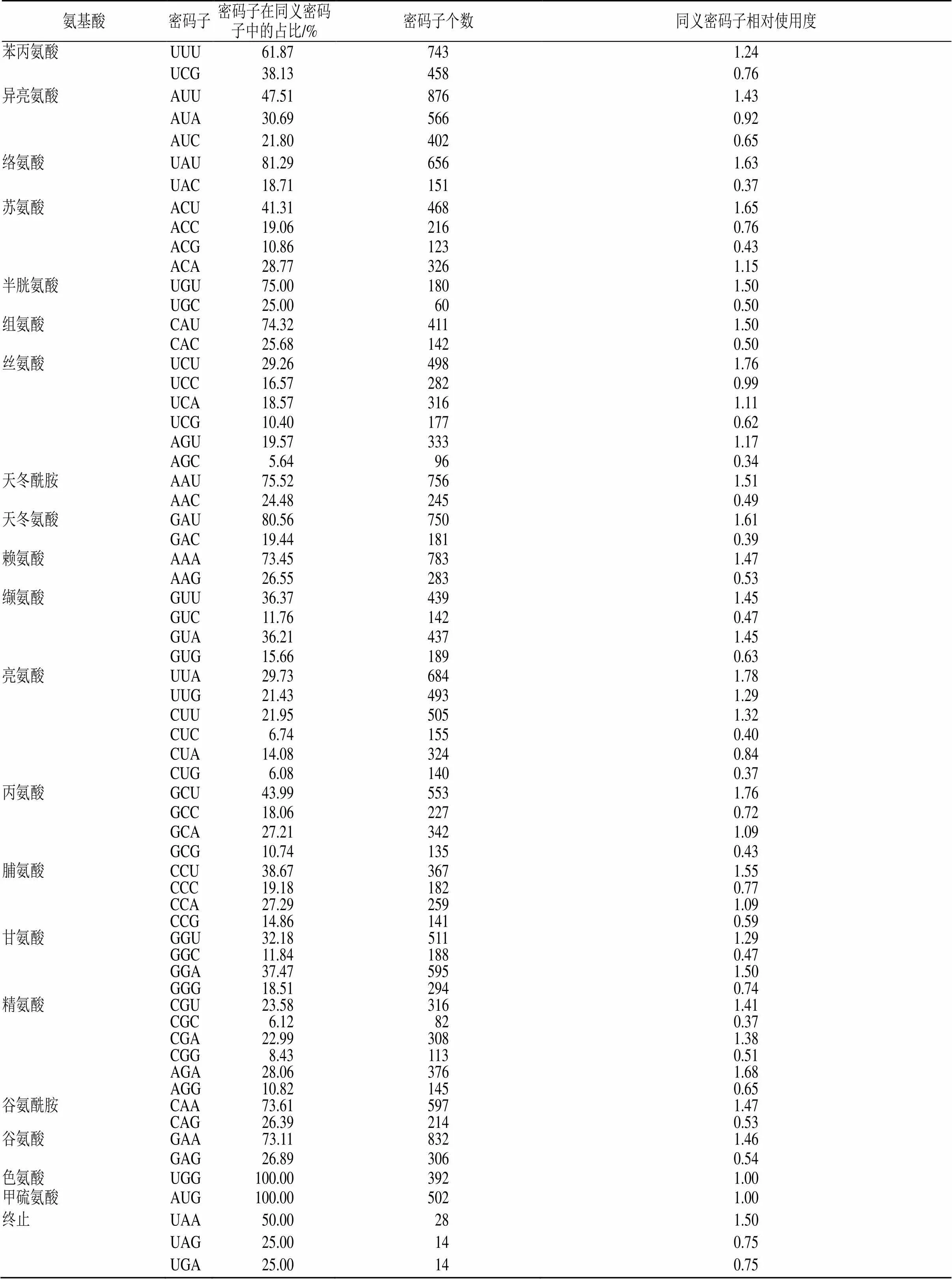
表3 紅花變豆菜葉綠體基因組密碼子使用情況
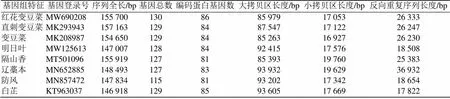
表4 傘形科內8種植物葉綠體基因組特征比較
2.7 葉綠體基因組全序列的聚類分析
葉綠體基因組的聚類分析對植物發育進化研究具有重要意義[28]。通過對紅花變豆菜與其他7種傘形科植物葉綠體基因組序列長度,基因結構以及編碼蛋白分析,發現其并未有太大差異,于是通過ML法對其構建系統發育樹,并加入玉米和擬南芥的葉綠體基因組序列作為外類群,對紅花變豆菜進行進化分析,以確定其在傘形科植物中的進化位置(圖2)。本課題組發現不同科的植物分為不同的進化分支,8種傘形科植物聚在了1個分支上;說明植物科間關系明確;在這個大的分支基礎上,紅花變豆菜與變豆菜和直刺變豆菜聚到了1個小分支上,并且自展值為100,說明這3個物種在漫長的進化過程中親緣關系最為接近。
3 討論
本研究采用高通量測序技術對紅花變豆菜葉綠體基因組進行重測序,并以已報道植物變豆菜葉綠體基因組為參考,成功組裝出其完整的葉綠體基因組。相對于傳統意義上葉綠體基因組序列的獲取,如紫荊澤蘭L.[29]和菝葜L.[30]都是采用先從植物樣本中分離出葉綠體,然后對其葉綠體進行DNA提取,最終再通過測序技術,實現葉綠體基因組序列的獲取,該方法復雜繁瑣、不易操作,不利于大范圍使用。利用高通量測序技術對紅花變豆菜全基因組DNA進行重測序,省略了先分離植物葉綠體再提取葉綠體DNA的復雜操作過程[31],只需提取紅花變豆菜全基因組DNA,進行高通量測序,選取已報道植物變豆菜葉綠體基因組序列作為參考基因組,將其所測得的全基因組序列與參考葉綠體基因組序列進行BLASTN比對,提取出關聯的葉綠體raw reads,使用過濾軟件SOAPnuke對reads進行低質量序列過濾得到clean reads,再利用SPAdes軟件對這些序列進行組裝及優化,最后使用GapCloser軟件對組裝結果進行補洞,最終得到完整的紅花變豆菜葉綠體基因組序列。本研究使用的方法相較于傳統方法,大大簡化了試驗步驟、降低了試驗所需的時間、減少了實驗成本,并且該方法實用性廣,限制條件少,為大量植物測序葉綠體基因組提供了可能。
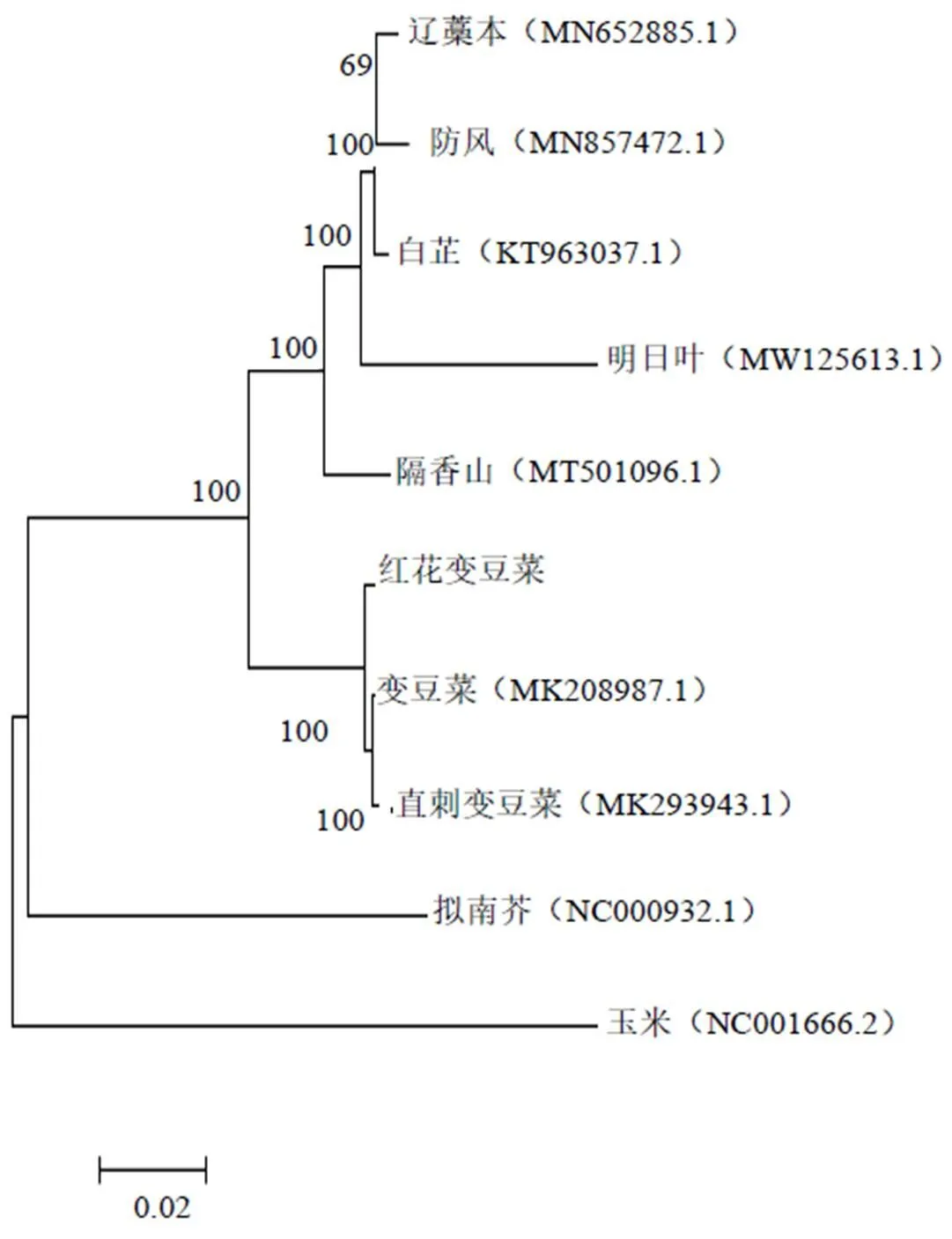
圖2 基于葉綠體全基因組10個物種的ML系統發育樹結果
變豆菜屬L.在克朗奎斯特系統(Cronquist system)、哈欽松系統、恩格勒系統(Engler system)、被子植物發育系統(angiosperm phylogeny group,APG)以及《中國植物志》等各個國家的植物分類體系中都被分為一個自然的類群,歸屬于傘形科(Apiaceae)、變豆菜亞科(Saniculoideae Drude)[32]。其中一些變豆菜屬植物具有一定的藥用價值,如變豆菜具有治療風濕、咳嗽和激活血液循環的功效,直刺變豆菜具有祛風止咳、活血通絡和清熱解毒的功效,也是一味著名的民族藥,收錄于《四川省中草藥標準(試行稿)》。中國變豆菜屬植物約有18種,屬于特有的為12種,但是鑒定困難,種間屬間易混淆,極其不利于此類植物的發展與利用。針對其分類已經有相關的研究,包括通過形態學研究和分子系統學研究[34]。本研究通過對紅花變豆菜葉綠體基本組進行測序及數據挖掘,分析了紅花變豆菜的長重復序列、SSR位點和密碼子偏好性,為變豆菜屬植物研究提供了更為詳細、完善的資料, 為該物種葉綠體基因工程和系統進化分析提供參考依據。
利益沖突 所有作者均聲明不存在利益沖突
[1] Wyk B E V, Tilney P M, Magee A R. African Apiaceae: A Synopsis of the Apiaceae/umbelliferae of sub-Saharan Africa and Madagascar [M]. Pretoria: Briza Academic Books, 2013: 288.
[2] Vandelook F, Assche J V. Deep complex morphophysiological dormancy in(Apiaceae) fits a recurring pattern of dormancy types in genera with an Arcto-Tertiary distribution [J]., 2008, 86: 1370-1377.
[3] 謝文遠, 馬丹丹, 陳鋒, 等. 黃花變豆菜: 浙江變豆菜屬(傘形科)一新種 [J]. 杭州師范大學學報: 自然科學版, 2019, 18(1): 9-12.
[4] 四川省中草藥標準(試行稿) [S]. 1979: 10.
[5] 周小江. 肺經草的品質研究[D]. 成都: 成都中醫藥大學, 2000.
[6] 姚雪瑩, 陳志祥, 王奇志. 變豆菜屬11種植物的葉表皮微形態特征研究 [J]. 植物研究, 2019, 39(5): 683-691.
[7] 馬孟莉, 張薇, 孟衡玲, 等. 豆蔻屬藥用植物葉綠體基因組密碼子偏性分析 [J]. 中草藥, 2021, 52(12): 3661-3670.
[8] 邢少辰, Clarke J L. 葉綠體基因組研究進展 [J]. 生物化學與生物物理進展, 2008, 35(1): 21-28.
[9] Zhang T W, Fang Y J, Wang X M,. The complete chloroplast and mitochondrial genome sequences of: Insights into the evolution of plant organellar genomes [J]., 2012, 7(1): e30531.
[10] Daniell H, Lin C S, Yu M,. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering [J]., 2016, 17(1): 134.
[11] Wang Z, Ren W C, Yan S,. Characterization of the complete chloroplast genome ofF. Schmidt ex Maxim [J]., 2021, 6(7): 1999-2000.
[12] Bankevich A, Nurk S, Antipov D,. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing [J]., 2012, 19(5): 455-477.
[13] Shi L C, Chen H M, Jiang M,. CPGAVAS2, an integrated plastome sequence annotator and analyzer [J]., 2019, 47(W1): W65-W73.
[14] Rozewicki J, Li S L, Amada K M,. MAFFT-DASH: Integrated protein sequence and structural alignment [J]., 2019, 47(W1): W5-W10.
[15] Kumar S, Stecher G, Li M,. MEGA X: Molecular evolutionary genetics analysis across computing platforms [J]., 2018, 35(6): 1547-1549.
[16] Tourasse N J, Shtaida N, Khozin-Goldberg I,. The complete mitochondrial genome sequence of the green microalga()reveals a new type of palindromic repetitive repeat [J]., 2015, 16(1): 580.
[17] Farhangian-Kashani S, Azadi A, Khaghani S,. Association analysis and evaluation of genetic diversity in wheat genotypes using SSR markers [J]., 2021, 72(4): 441-452.
[18] Shioi S, Shimamoto A, Nakagami Y,. Precision length determination andsimulation in PCR of microsatellite repeat sequences [J]., 2021, 42(12/13): 1323-1332.
[19] 陳中蘇直, 田波, 蔡傳濤. 基于SSR分子標記的滇重樓遺傳多樣性研究 [J]. 中草藥, 2017, 48(9): 1834-1838.
[20] 周茜, 陳蕓, 王玉州, 等. 獨行菜LaBBX基因低溫表達響應與密碼子偏性分析 [J]. 核農學報, 2021, 35(6): 1253-1262.
[21] Chen Z X, Yao X Y, Downie S R,. Assembling and analysis ofchloroplast genome [J]., 2019, 27(4): 366-372.
[22] Chen Z X, Yao X Y, Wang Q Z. The complete chloroplast genome of[J].DNA Part B, 2019, 4(1): 734-735.
[23] Yan W J, Yang T G, Qin R,. Complete chloroplast genome of(Umbelliferae) [J]., 2021, 6(2): 536-537.
[24] Liao C Y, Gao Q, Wu F. The chloroplast genome of(Apiaceae), an endemic medicinal plant to China [J]., 2020, 5(3): 3782-3783.
[25] Wang S J, Zhang T T, Xu L,. The complete chloroplast genome of(Umbelliferae,L.) [J]., 2020, 5(3): 3025-3027.
[26] Li L L, Geng M L, Li Y S,. Characterization of the complete plastome of(Turcz.) schischk [J]., 2020, 5(1): 786-787.
[27] Zhang R, Xu B H, Cao T Y. Characterization of the complete chloroplast genome of(Apiaceae) as an herb in China [J]., 2020, 5(1): 678-679.
[28] 朱婷婷, 張磊, 陳萬生, 等. 1342個植物葉綠體基因組分析 [J]. 基因組學與應用生物學, 2017, 36(10): 4323-4333.
[29] Nie X J, Lv S Z, Zhang Y X,. Complete chloroplast genome sequence of a major invasive species, crofton weed () [J]., 2012, 7(5): e36869.
[30] Liu J, Qi Z C, Zhao Y P,. Complete cpDNA genome sequence ofand phylogenetic placement of Liliales: Influences of gene partitions and taxon sampling [J]., 2012, 64(3): 545-562.
[31] 王虹雨, 李錦萍, 張瑞峰, 等. 基于高通量技術的橢圓葉花錨葉綠體全基因組測序及系統發育分析[J/OL]. 分子植物育種: [2022-10-21]. http://kns.cnki.net/kcms/ detail/46.1068.S.20210611.1420.012.html.
[32] 姚雪瑩. 直刺變豆菜的譜系地理學和遺傳多樣性研究 [D]. 泉州: 華僑大學, 2020.
[33] 陳志祥, 姚雪瑩, Stephen R D, 等. 直刺變豆菜葉綠體全基因組及其特征 [J]. 生物多樣性, 2019, 27(4): 366-372.
[34] 楊晨, 陳志祥, 姚雪瑩, 等. 中國15種變豆菜屬植物的花粉形態及系統學分析 [J]. 植物研究, 2020, 40(6): 805-812.
Assembly and sequence analysis of chloroplast genome of
WANG Zhen1, LIU Chi2, REN Wei-chao1, ZHANG Mei-qi1, LIU Xiu-bo1, MA Wei1, 3, 4
1. College of Pharmaceutical Sciences, Heilongjiang University of Chinese Medicine, Harbin 150040, China 2. School of Information, Technical University of Kemnitz, Kemnitz 09111, Germany 3. Jiangsu Kanion Parmaceutical Co., Ltd., Lianyungang 222001, China 4. State Key Laboratory of New-tech for Chinese Medicine Pharmaceutical Process, Lianyungang 222001, China
The chloroplast genome was assembled and the sequence characteristics analyzed with fresh leaves ofas the experimental materials through high-throughput sequencing, so as to provide guidance for the further development of the evolution and genetic development of.The experimental procedures were carried out according to Illumina's standard operating procedures, including the sample quality test, the construction of library, the quality test of the sample library and the sequencing of the library, the reported chloroplast genome ofwas used as a reference sequence, and the complete chloroplast genome ofwas obtained and assembled, and the assembly characteristic sequence analysis and evolutionary analysis were performed.The chloroplast genome ofhad a typical circular quadripartite structure with a size of 155 700 bp in length. In which a large single-copy (LSC) of 85 979 bp and a pair of inverted repeats (IRs) of 26 333 bp were disconnected by a small single-copy (SSC) of 17 053bp. A total of 130 genes were annotated, including 86 protein-coding genes (PCGs), 36 transfer RNA genes (tRNAs) and eight ribosomal RNA genes (rRNAs). AT content accounted for 61.83% of the chloroplast genome. One forward repeat sequence and three palindromic repeat sequences were detected. In addition, 168 simple sequence repeats (SSR) sites were detected. The codon corresponding to the protein coding gene preferred to use A/T base. The codon (I) encoding isoleucine had the highest frequency of use, while the codon (C) encoding cysteine had the lowest frequency. A phylogenetic tree with maximum likelihood (ML) was constructed based on the chloroplast genomes of seven reported Apiaceae species and two other species. It was found thatwere most closely related withand.The chloroplast genome ofprovides more detailed and complete data for the study of Apiaceae plants, and provides a reference basis for chloroplast genetic engineering and systematic evolution analysis of this species.
Fr. Schmidt; complete chloroplast genome; high-throughput sequencing; phylogeny; maximum likelihood
R282.12
A
0253 - 2670(2022)22 - 7183 - 08
10.7501/j.issn.0253-2670.2022.22.022
2022-02-02
中央本級重大增減支項目:名貴中藥資源可持續利用能力建設項目(2060302);中藥制藥過程新技術國家重點實驗室開放基金項目(SKL2020M0302)
王 震(1995—),男,碩士,研究方向為藥用植物工程。Tel: (0451)87266827 E-mail: 1021707041@qq.com
馬 偉,博士,研究員,博士生導師,研究方向為藥用植物生物工程與中藥資源。Tel: (0451)87266827 E-mail: mawei@hljucm.net
劉秀波(1975—),教授,研究方向為藥用植物生物工程。Tel: 13796353268 E-mail: 358270831@qq.com
#共同第一作者:柳 馳(1991—),男,碩士,研究方向為信息分析。Tel: (0451)87266827
[責任編輯 時圣明]
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
