基于轉錄組學研究毛冬青三萜皂苷對脂多糖誘導小鼠腸損傷的保護作用及其機制
2022-11-18 02:24:34陳冰瑩伍鈺蓉張穎茵
中草藥 2022年22期
王 瑩,陳冰瑩,伍鈺蓉,張穎茵,王 倩,李 瑜,張 蕾*
基于轉錄組學研究毛冬青三萜皂苷對脂多糖誘導小鼠腸損傷的保護作用及其機制
王 瑩1,陳冰瑩1,伍鈺蓉1,張穎茵1,王 倩1,李 瑜2,張 蕾1*
1. 廣州中醫藥大學中藥學院,廣東 廣州 510006 2. 廣州中醫藥大學護理學院,廣東 廣州 510006
探討毛冬青三萜皂苷(triterpenoid saponin,IPTS)對脂多糖(lipopolysaccharide,LPS)誘導的小鼠腸損傷的作用及潛在機制。C57BL/6小鼠隨機分為對照組、模型組和IPTS低、中、高劑量(88、176、264 mg/kg)組,給予藥物干預7 d,ip LPS誘導小鼠小腸損傷,造模7 h后,采用蘇木素-伊紅(HE)染色法觀察小鼠回腸組織病理變化;檢測小鼠血清中丙二醛(malondialdehyde,MDA)水平和超氧化物歧化酶(superoxide dismutase,SOD)活性;采用qRT-PCR和Western blotting法檢測回腸組織炎癥因子白細胞介素-1β(interleukin-1β,)、腫瘤壞死因子-α(tumor necrosis factor-α,)mRNA和腸屏障緊密連接蛋白、mRNA及蛋白表達。通過轉錄組學測序挖掘IPTS作用于回腸的潛在機制。與模型組比較,IPTS組小鼠回腸組織完整,氧化應激損傷改善(<0.001),回腸炎癥因子水平降低(<0.01),腸屏障恢復(<0.05、0.01、0.001)。RNA-seq測序結果顯示,與對照組比較,模型組共篩選出2584個差異表達基因;與模型組比較,IPTS高劑量組共篩選出259個差異表達基因;這些差異表達基因主要富集在免疫球蛋白A(immunoglobulin A,IgA)相關的腸道免疫、花生四烯酸代謝、B細胞受體、瞬時受體電位(transient receptor potential,TRP)通道炎癥調控、亞油酸代謝等信號通路。IPTS可以改善LPS誘導的小鼠小腸損傷,可能與花生四烯酸代謝、IgA相關的腸道免疫代謝以及TRP通道炎癥調控通路密切相關。
毛冬青三萜皂苷;腸損傷;轉錄組學;氧化應激;炎癥;腸屏障
毛冬青Hook. et Am.是冬青科冬青屬植物,是我國南方常用中藥,其主要功效為清熱解毒、活血通絡。中醫通過辨證論治,廣泛應用毛冬青治療心腦血管疾病,其在降低不良反應等方面明顯優于化學藥,且療效穩定,無耐藥性和成癮性,患者容易接受[1]。植物化學和藥效學研究結果表明,毛冬青的主要活性成分為三萜皂苷[2]。然而,當將毛冬青三萜皂苷(triterpenoid saponin,IPTS)的體內過程納入藥理機制的闡釋中時,卻有系列問題亟待解決:毛冬青主要藥效物質三萜皂苷類成分口服給藥血藥濃度低,即便相對吸收特性較好的苷元如毛冬青甲素,單體口服給藥的絕對生物利用度<3%[3-4]。本課題組前期組織分布研究結果發現,大鼠ig毛冬青提取液后,IPTS主要分布于胃腸道,僅有少量分布于心臟,腦中更是痕量的分布,因此現有的基于非口服給藥途徑的藥理學研究結果尚無法詮釋其口服用藥抗心腦血管性疾病的作用機制。
本課題組在研究IPTS抗動脈粥樣硬化作用機制時發現,IPTS不但可以調控腸道菌群結構改善動脈粥樣硬化,還能顯著降低動脈粥樣硬化小鼠血清中脂多糖(lipopolysaccharide,LPS)水平[5]。文獻報道,動脈粥樣硬化疾病常伴隨腸黏膜屏障功能障礙,使其通透性增大,從而導致腸道菌群產生的LPS透過腸屏障進入血液循環中。當LPS從腸道轉移到循環中時,與LPS結合蛋白形成復合體,可以與單核細胞上的CD14結合進而導致促炎細胞因子的產生[6]。基于“腸-心”軸理論、毛冬青在民間的抗炎用途和其組織分布特點,推測毛冬青具有抗小腸炎性損傷,維持腸黏膜屏障緊密連接和完整性,進而發揮其抗心腦血管疾病的作用。由于通過高脂飲食制備的動脈粥樣硬化模型伴腸道損傷周期長、成模率低,本研究通過ip LPS建立小鼠小腸損傷的炎癥模型,探討IPTS保護小腸黏膜屏障的作用,并結合轉錄組學測序分析,探索其潛在機制和作用靶點。
1 材料
1.1 動物
SPF級雄性C57BL/6小鼠,體質量18~22 g,購自珠海百試通生物科技有限公司,動物合格證號SCXK(粵)2020-0051。動物于廣州中醫藥大學中藥學院動物房SPF級環境中適應性飼養1周,溫度(24±2)℃、12 h光晝交替,自由進食飲水。實驗單位使用許可證編號為SYXK(粵)2019-0202,動物實驗經廣州中醫藥大學中藥學院實驗動物倫理審查委員會批準(批準號ZYD-2021-223)。
1.2 藥材
毛冬青購自廣州白云山中藥飲片有限公司,經廣州中醫藥大學張丹雁教授鑒定為冬青科冬青屬植物毛冬青Hook. et Am.的干燥根。
1.3 藥品與試劑
毛冬青皂苷B1對照品(質量分數>98%)由廣州中醫藥大學趙鐘祥教授課題組提供;4%多聚甲醛固定液(批號21211717)購自北京Biosharp公司; PBS緩沖液(批號GA21120122694)購自武漢賽維爾生物科技有限公司;LPS(批號12190801)購自美國Sigma公司;生理鹽水(批號0506A22)購自雷根生物公司;超氧化物歧化酶(superoxide dismutase,SOD)檢測試劑盒(批號20211119)、丙二醛(malondialdehyde,MDA)檢測試劑盒(批號20211120)購自南京建成生物工程研究所;TRIzol試劑(批號03877118721)、HiFiScript gDNA Removal RT Master Mix試劑盒(批號02322)、Magic SYBR Mixture試劑盒(批號09321)購自康為世紀公司;小鼠甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)引物(批號I620KA5091)購自生工生物工程公司;DEPC水(批號051821210702)購自北京索萊寶公司;GAPDH抗體(批號AC21110131A)購自武漢塞維爾生物科技有限公司;ZO-1抗體(批號GR3398704-6)、Occludin抗體(批號GR3383643-14)購自英國Abcam公司;山羊抗兔IgG抗體(批號S0001)購自Affinity公司;Marker(批號91226047)購自美國Thermo Fisher Scientific公司。
1.4 儀器
LC-LX-HR165A型冷凍離心機(上海力辰公司);Eclipse E100型正置光學顯微鏡(日本尼康公司);THM51 119300型全波長酶標儀、NC2000型Nanodrop紫外定量儀、ABI 7500型熒光定量PCR儀(美國Thermo Fisher Scientific公司);SDS-PAGE垂直電泳槽、垂直轉印槽(美國Bio-Rad公司);Illumina Misseq測序儀(美國Illumina公司)。
2 方法
2.1 IPTS的制備
毛冬青根莖飲片打成粗粉,用10倍量的蒸餾水浸泡24 h,浸泡2次,并定時攪拌,濾過,棄去濾液,濾過后的粗粉按料液比1∶10加入85%乙醇加熱回流提取2次,每次1 h,收集全部濾液,濃縮,干燥后得到毛冬青提取粉末。以毛冬青皂苷B1為對照品,采用比色法測定總三萜皂苷的質量分數,以對照品濃度為橫坐標(),吸光度為縱坐標(),得標準曲線=369.95-0.052 9(2=1.000),計算得三萜皂苷的質量分數為81.6%。
2.2 造模、分組及給藥
C57BL/6小鼠隨機分為對照組、模型組和IPTS低、中、高劑量(88、176、264 mg/kg)組,每組12只。IPTS溶于0.5%羧甲基纖維素鈉(CMC-Na)配制成質量濃度為8.8、17.6、26.4 mg/mL的混懸液,各給藥組ig相應藥物(10 mL/kg),對照組和模型組ig等體積的0.5% CMC-Na,1次/d,連續7 d。末次給藥后,模型組和各給藥組ip LPS(10 mg/kg),造模7 h后,記錄小鼠造模前后體質量及狀態變化,計算小鼠ip LPS前后體質量差值。
2.3 IPTS對腸損傷小鼠血清中MDA水平及SOD活性的影響
造模7 h后,對小鼠進行麻醉,摘眼球取血,室溫靜置1 h后,4 ℃、3800 r/min離心10 min,收集上清液,于?80 ℃保存。按照試劑盒說明書測定血清中MDA水平及SOD活性。
2.4 IPTS對腸損傷小鼠回腸組織病理變化的影響
小鼠脫頸椎處死后,在冰上取回腸,用PBS溶于小心沖洗,取0.5 cm置于4%多聚甲醛溶液中固定,經石蠟包埋后切片,進行蘇木素-伊紅(HE)染色,于光學顯微鏡下觀察小腸組織病理變化。
2.5 IPTS對腸損傷小鼠回腸組織炎癥因子白細胞介素-1β(interleukin-1β,IL-1β)、腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)和腸屏障蛋白ZO-1、Occludin mRNA表達的影響
取各組小鼠回腸組織20 mg,按照試劑盒說明書提取總RNA并合成cDNA,進行qRT-PCR分析。引物序列:上游引物5’-CACTACAGGCTCCG- AGATGAACAAC-3’、下游引物5’-TGTCGTTGCTT- GGTTCTCCTTGTAC-3’;上游引物5’-AGAGGTTGGCTATGGAGGCTGTG-3’、下游引物5’-CCTGCATCGTTGACGGTCTTCC-3’;上游引物5’-AACCCGAAACTGATGCTGTGGATAG-3’、下游引物5’-CGCCCTTGGAATGTATGTGGAGAG-3’;上游引物5’-TTGGCTACGGAGGTG- GCTATGG、下游引物5’-CCTTTGGCTGCTCTTG-GGTCTG-3’;上游引物5’-GGTTGTCTCCT- GCGACTTCA-3’、下游引物5’-TGGTCCAGGGTT- TCTTACTCC-3’。
2.6 IPTS對腸損傷小鼠回腸組織ZO-1和Occludin蛋白表達的影響
取各組小鼠回腸組織30 mg,加入RIPA裂解液和蛋白酶抑制劑,使用冷凍組織研磨儀進行勻漿(60 Hz、60 s),勻漿后放于碎冰上靜置20 min,4 ℃、1200 r/min離心5 min,取上清液。采用BCA蛋白定量試劑盒測定蛋白濃度,加入Loading buffer后混勻,沸水煮15 min使蛋白變性。蛋白樣品經十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳,轉至PVDF膜,加入5%脫脂牛奶,室溫封閉2.5 h;分別加入相應抗體,4 ℃孵育過夜;加入二抗,室溫孵育1 h,加入ECL工作液發光顯色,采用Image J軟件分析條帶灰度值。
2.7 RNA-Seq測序分析
2.7.1 RNA提取和RNA-Seq測序 分別取對照組、模型組、IPTS高劑量組回腸組織100 mg,置于RNase-Free凍存管內,于液氮中冷凍,委托(上海)美吉生物公司進行RNA提取和RNA-Seq測序,并使用該公司在線分析平臺進行分析。轉錄測序具體流程如下:采用TruSeqTMRNA sample preparation Kit試劑盒建立RNA文庫。首先利用帶有Oligo(dT)的磁珠從1 μg總RNA中富集有poly-A尾的mRNA,再加入fragmentation buffer,將mRNA隨機斷裂成300 bp左右的小片段,接著采用SuperScript double-stranded cDNA synthesis kit試劑盒,加入六堿基隨機引物,以mRNA為模板反轉合成一鏈cDNA,隨后進行二鏈合成,形成穩定的雙鏈結構。雙鏈的cDNA結構為黏性末端,加入End Repair Mix將其補成平末端,隨后在3’末端加上一個A堿基,用于連接Y字形的接頭。cDNA經過PCR富集后,beads(DNA clean beads)篩選200~300 bp的條帶。經TBS380定量后,使用Ilumina HiSeq xten/NovaSeq 6000測序平臺進行高通量測序,測序讀長為PE150。
2.7.2 測序濾過原始數據質控對比 對原始測序數據進行濾過,去除reads中的接頭序列,去除序列末端質量值小于20的堿基,去除含N比率超過10%的reads,舍棄去adapter及質量修剪后長度小于20 bp的序列,對濾過的數據與參考基因組比對分析。
2.7.3 基因差異表達分析 在RNA-Seq分析中,通過定位到基因組區域的序列數來計算基因的表達水平。使用RSEM軟件對基因的表達水平進行定量分析,獲得基因的序列數后,使用基于負二項分布的DESeq2軟件對Raw數進行分析,基于一定的標準化處理和篩選條件獲得比較組間表達差異的基因,最后按照表達量倍數差異和表達差異顯著性進行篩選,校正的值(adjust)<0.05且|log2FC|≥2。
2.7.4 基因本體(gene ontology,GO)功能和京都基因與基因組百科全書(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析 利用GO數據庫,使用Goatools軟件,進行Fisher精確檢驗,對差異基因進行GO功能顯著性富集分析。采用BH方法對值進行校正,當adjust<0.05時,認為此功能存在顯著富集情況。使用KOBAS進行KEGG通路富集分析,計算原理同GO功能富集分析,滿足adjust<0.05的KEGG通路定義為在差異表達基因中顯著富集的KEGG通路。
2.7.5 IPTS對LPS誘導的腸損傷小鼠回腸組織關鍵基因表達的影響 通過qRT-PCR驗證關鍵差異基因肌醇-1,4,5-三磷酸受體2(inositol-1,4,5-trisphosphate receptor type 2,)、嘌呤能受體P2Y2(purinergic receptor P2Y2,)、整合素亞基α4(integrin subunit α4,)、、趨化因子受體4(C-X-C motif chemokine receptor 4,)、細胞色素P450家族2亞家族C成員55(cytochrome P450 family 2 subfamily C member 55,)、、、、Ⅴ組磷脂酶A2(phospholipase A2 group V,)的表達。取小鼠回腸組織20 mg,按“2.5”項下方法進行qRT-PCR分析,引物序列見表1。

表1 引物序列
2.8 統計學分析
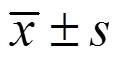
3 結果
3.1 IPTS對腸損傷小鼠體質量的影響
如圖1所示,與對照組比較,模型組小鼠體質量丟失明顯升高(<0.01);與模型組比較,各給藥組小鼠體質量丟失均明顯降低(<0.01)。表明小鼠ip LPS會造成體質量下降,IPTS可以緩解LPS造成的小鼠體質量下降。
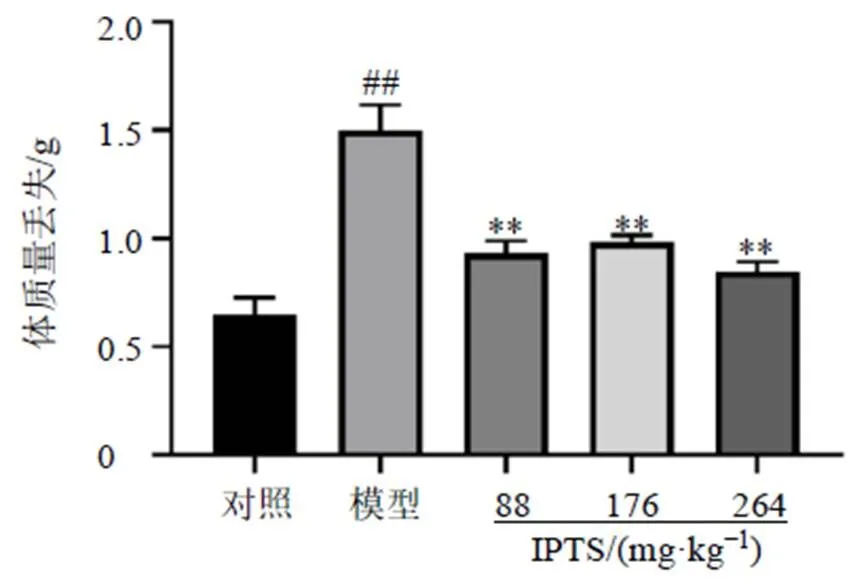
3.2 IPTS對腸損傷小鼠血清中MDA水平及SOD活性的影響
如圖2所示,與對照組比較,模型組小鼠血清中MDA水平顯著升高(<0.001),SOD活性顯著降低(<0.001);與模型組比較,IPTS高劑量組小鼠血清中MDA水平顯著降低(<0.001),各給藥組SOD活性均顯著升高(<0.001)。表明小鼠ip LPS后,體內發生氧化應激損傷,IPTS能夠緩解氧化應激損傷。
與對照組比較:#P<0.05 ##P<0.01 ###P<0.001;與模型組比較:*P<0.05 **P<0.01 ***P<0.01,下圖同
3.3 IPTS對腸損傷小鼠回腸組織病理變化的影響
如圖3所示,對照組小鼠回腸組織結構正常,絨毛排列緊密,細胞形態正常,無組織學病變。模型組小鼠回腸絨毛斷裂,腸黏膜上皮細胞脫落。與模型組比較,IPTS低、中劑量組小鼠回腸絨毛恢復,僅有小部分脫落;IPTS高劑量組小鼠回腸損傷較輕,腸黏膜結構基本完整,腸絨毛大部分排列整齊。
3.4 IPTS對腸損傷小鼠回腸組織炎癥因子IL-1β和TNF-αmRNA表達的影響
如圖4所示,與對照組比較,模型組小鼠回腸組織中和的mRNA表達水平均顯著升高(<0.01);與模型組比較,各給藥組小鼠回腸組織中mRNA表達水平顯著降低(<0.01),IPTS中、高劑量組回腸組織中mRNA表達水平顯著降低(<0.01)。表明IPTS能夠顯著降低LPS誘導的腸損傷小鼠回腸組織中炎癥因子表達。
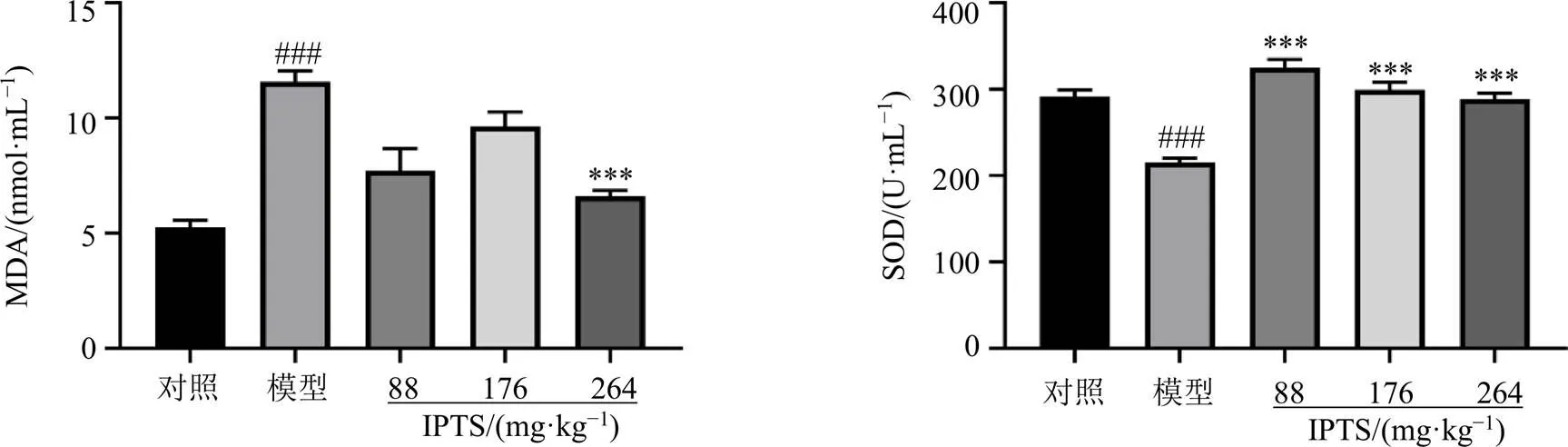
圖2 IPTS對腸損傷小鼠血清中MDA水平及SOD活性的影響(, n = 6)
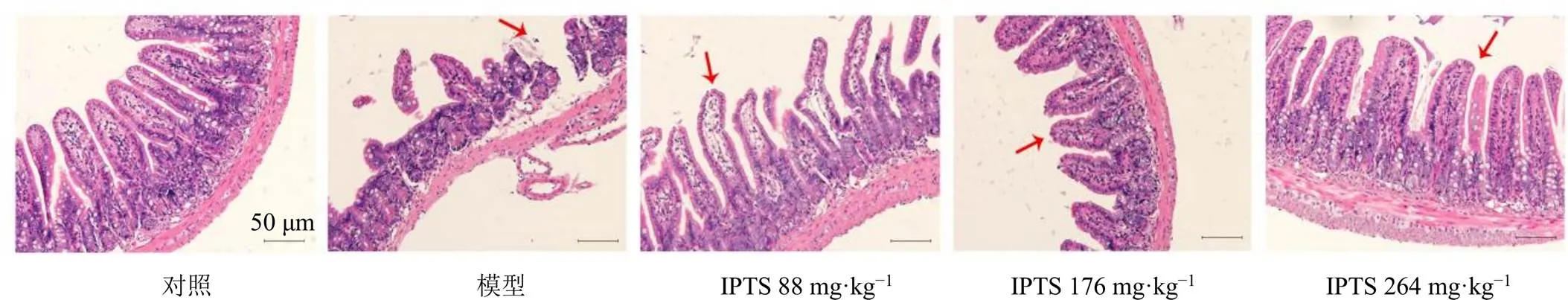
箭頭表示小腸絨毛受損
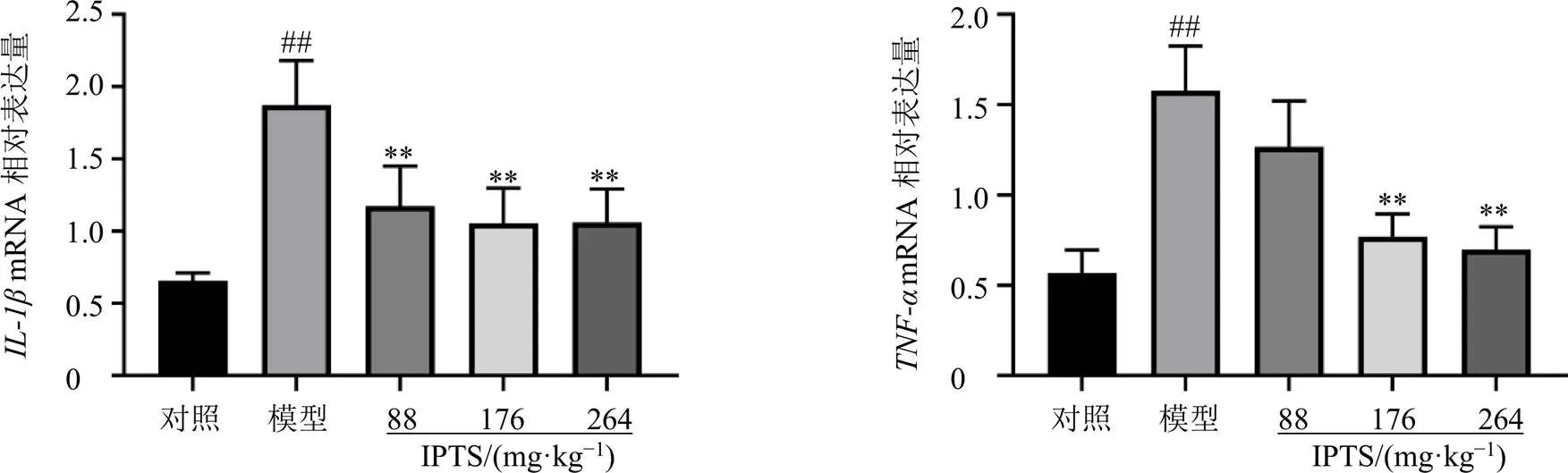
圖4 IPTS對腸損傷小鼠回腸組織IL-1β和TNF-αmRNA表達的影響(, n = 6)
3.5 IPTS對腸損傷小鼠回腸組織腸屏障蛋白ZO-1、OccludinmRNA和蛋白表達的影響
如圖5所示,與對照組比較,模型組小鼠回腸組織中屏障蛋白、的mRNA表達水平均顯著降低(<0.001);與模型組比較,IPTS高劑量組回腸組織中的mRNA表達水平顯著升高(<0.001),IPTS中、高劑量組回腸組織中的mRNA表達水平明顯升高(<0.05、0.001)。
如圖6所示,與對照組比較,模型組小鼠回腸組織中ZO-1和Occludin蛋白表達水平均顯著降低(<0.001);與模型組比較,IPTS中、高劑量組回腸組織中ZO-1和Occludin蛋白表達水平均顯著升高(<0.01、0.001)。
以上結果表明,小鼠ip LPS后,回腸組織中腸屏障蛋白和的mRNA及蛋白表達均顯著降低,造成回腸腸屏障損傷;IPTS能夠顯著恢復其表達,保護回腸腸屏障。
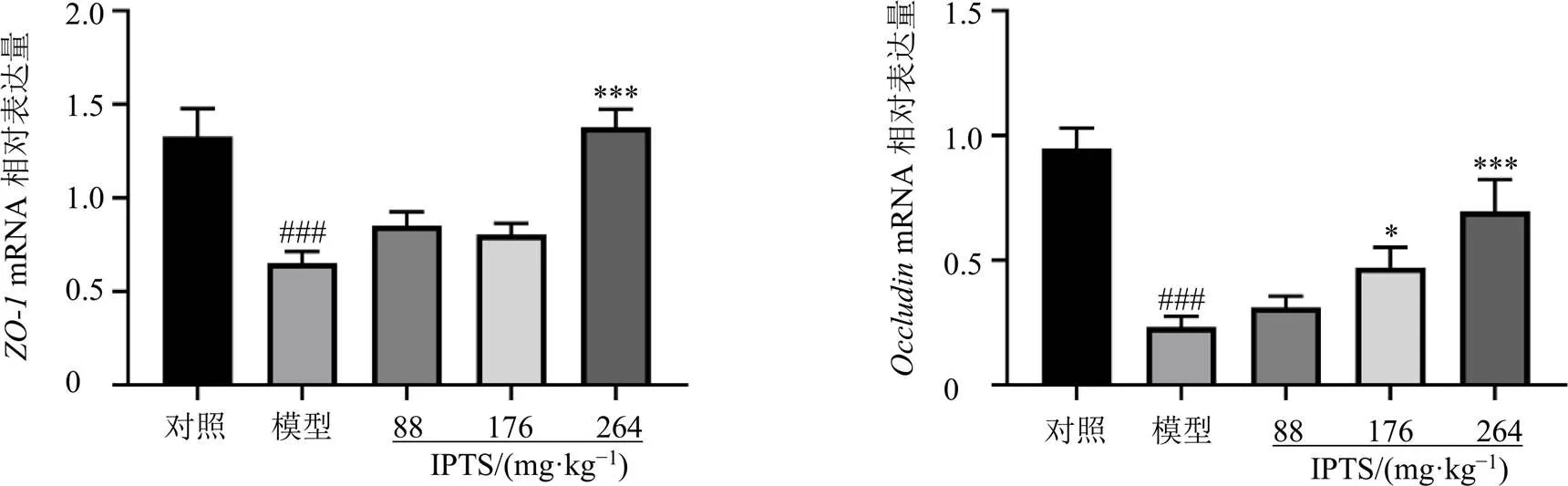
圖5 IPTS對腸損傷小鼠回腸組織ZO-1和OccludinmRNA表達的影響(, n = 6)
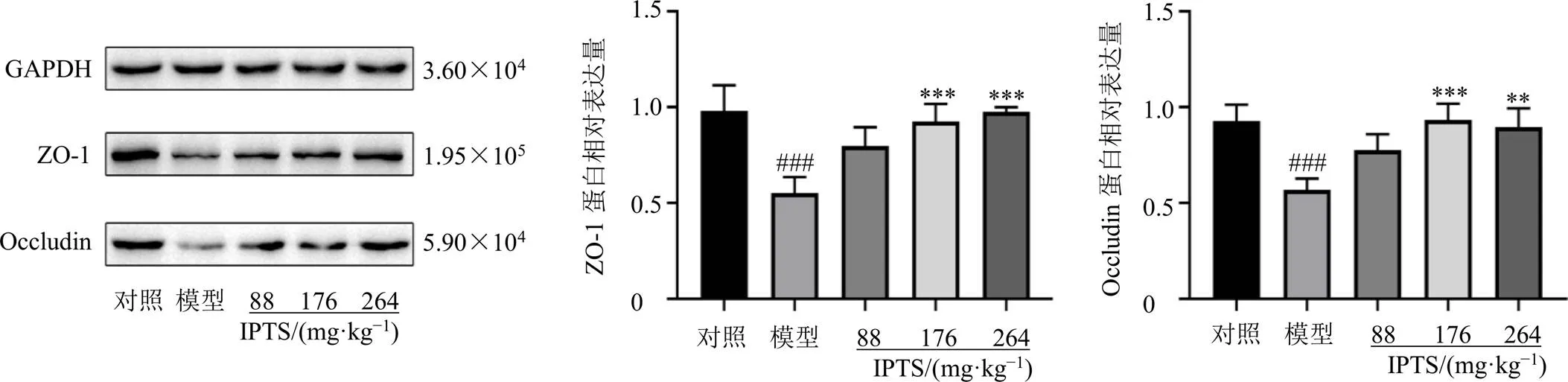
圖6 IPTS對腸損傷小鼠回腸組織ZO-1和Occludin蛋白表達的影響(, n = 6)
3.6 轉錄組學測序數據的質量評估
本研究共完成14個樣品(對照組4個、模型組5個、IPTS高劑量組5個)的轉錄組測序分析。經質量控制所有樣品的誤差率小于0.025 3%,所有樣品的20均大于97.85%,30均大于93.94%。根據飽和度曲線可知,所有樣品測序數據量已滿足定量要求。覆蓋度分布圖說明測序無偏向性,測序所得序列在基因上均勻分布。
3.7 基因差異表達分析結果
模型組和對照組之間有2584個差異表達基因(differentially expressed genes,DEGs),與對照組比較,模型組上調DEGs 1119個,下調DEGs 1465個;IPTS高劑量組和模型組之間有259個DEGs,與模型組比較,IPTS高劑量組上調DEGs 208個,下調DEGs 51個。由差異表達基因聚類分析圖(圖7)可知,IPTS在一定程度上回調了小鼠ip LPS后回腸組織DEGs。
3.8 DEGs富集分析
將篩選出的DEGs進行GO和KEGG富集分析。GO富集結果見圖8,模型組與對照組相比,與分子功能相關的差異基因主要富集在連接、催化活性;與細胞成分相關的主要富集于細胞部分、細胞器、細胞膜;與生物過程相關的主要富集在細胞過程、生物調控。KEGG通路主要富集在IgA生成相關腸道免疫網絡、甾體激素生物合成、磷脂酰肌醇3-激酶(phosphatidylinositol-3-kinase,PI3K)-蛋白激酶B(protein kinase B,Akt)信號通路、TNF信號通路、NOD樣受體信號通路、核因子-κB(nuclear factor-κB,NF-κB)信號通路等。
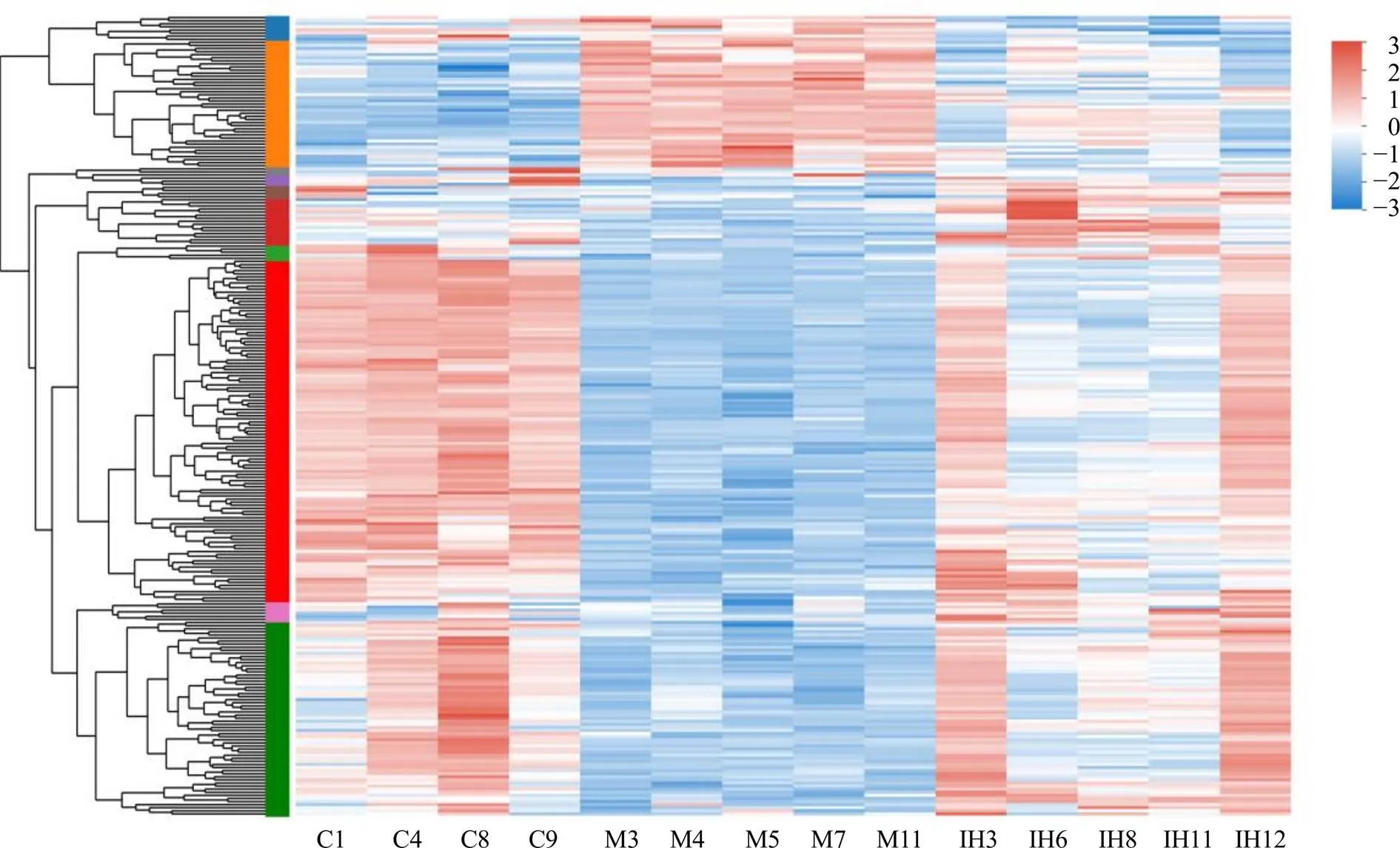
C-模型組 M-模型組 IH-IPTS高劑量組
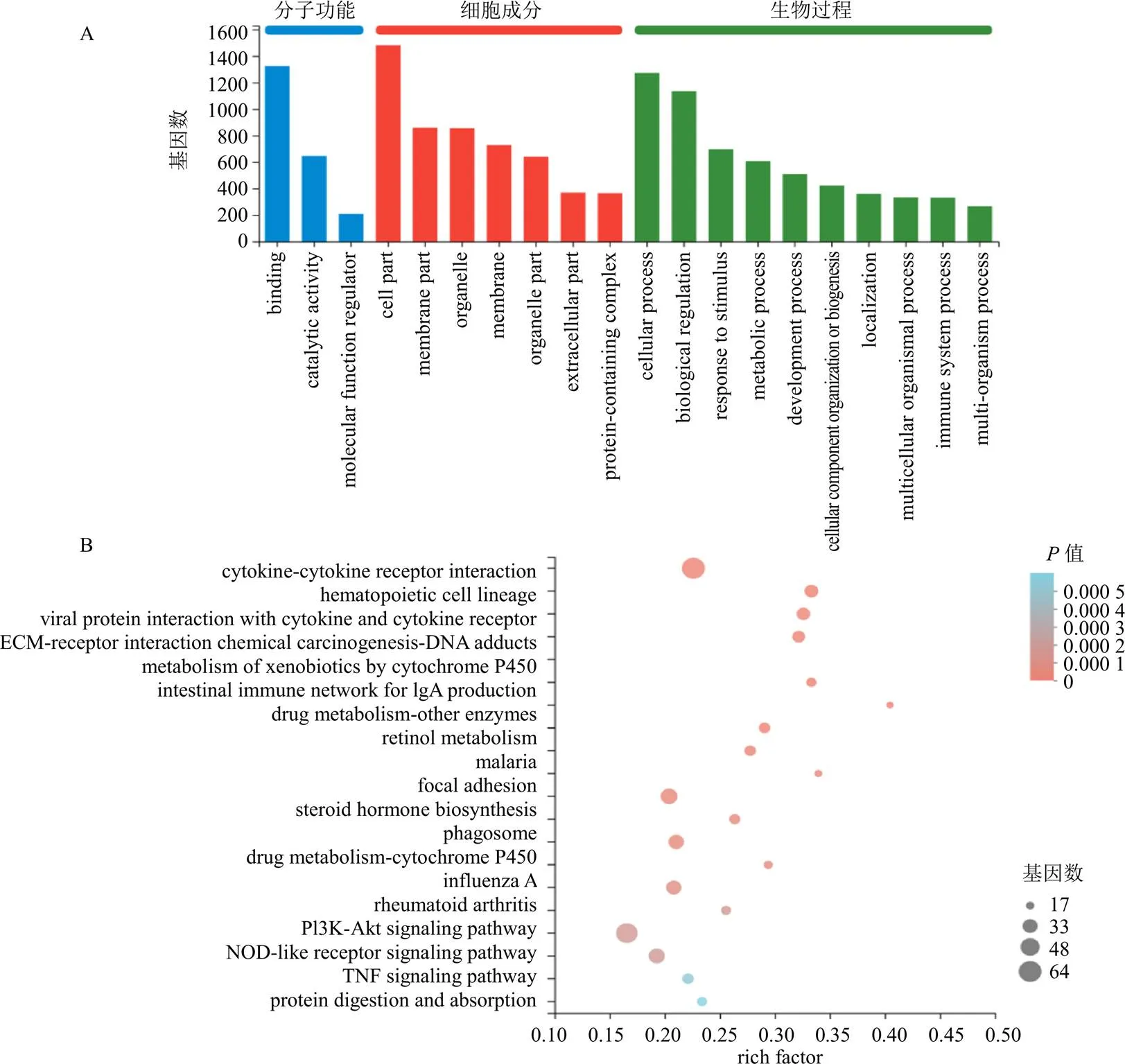
圖8 對照組與模型組DEGs的GO功能 (A)和KEGG通路 (B)富集分析
如圖9所示,IPTS高劑量組與模型組相比,與分子功能相關的差異基因主要富集在連接、催化活性;與細胞成分相關的主要富集于細胞部分、細胞器、細胞膜;與生物過程相關的主要富集在細胞過程、生物調控、代謝過程。KEGG通路主要富集在免疫球蛋白A(immunoglobulin A,IgA)生成相關腸道免疫網絡、亞油酸代謝、色氨酸代謝、花生四烯酸代謝、藥物代謝-細胞色素P450、B細胞受體信號通路、瞬時受體電位(transient receptor potential,TRP)通道炎癥調控通路等。
3.9 IPTS對腸損傷小鼠回腸組織關鍵基因表達的影響
以為內參基因,通過qRT-PCR對RNA-Seq分析結果中TRP通道炎癥調控、花生四烯酸代謝、IgA相關的腸道免疫相關通路的10個關鍵基因、、、、、、、、、進行mRNA表達水平的驗證。如圖10所示,與對照組比較,模型組小鼠回腸組織中、、、、、、、、mRNA表達水平均顯著降低(<0.05、0.01、0.001),mRNA表達水平顯著升高(<0.001);與模型組比較,IPTS高劑量組回腸組織中、、、、、、、、mRNA表達水平均顯著升高(<0.05、0.01),mRNA表達水平顯著降低(<0.01)。
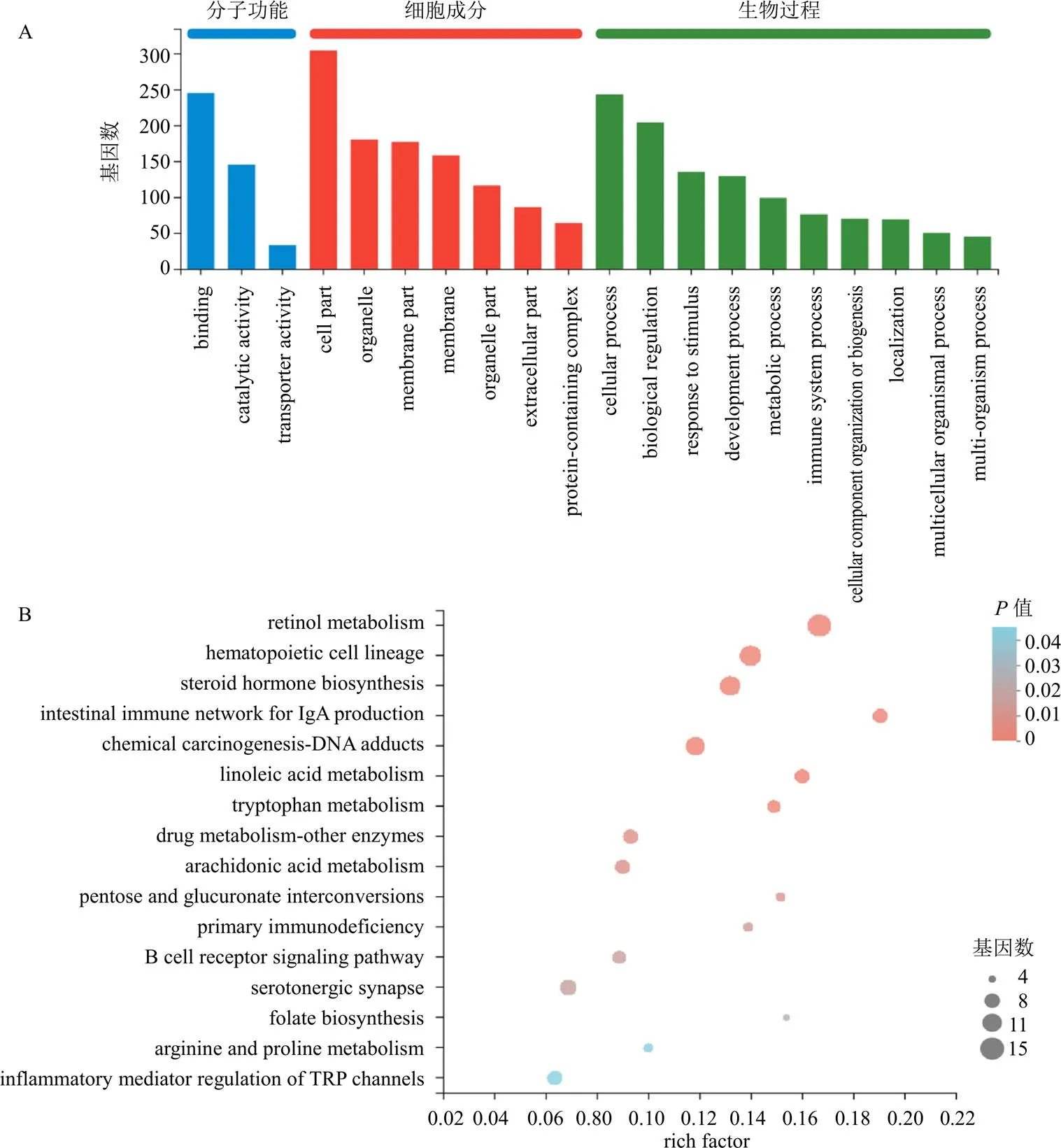
圖9 模型組與IPTS高劑量組DEGs的GO功能 (A)和KEGG通路 (B)富集分析
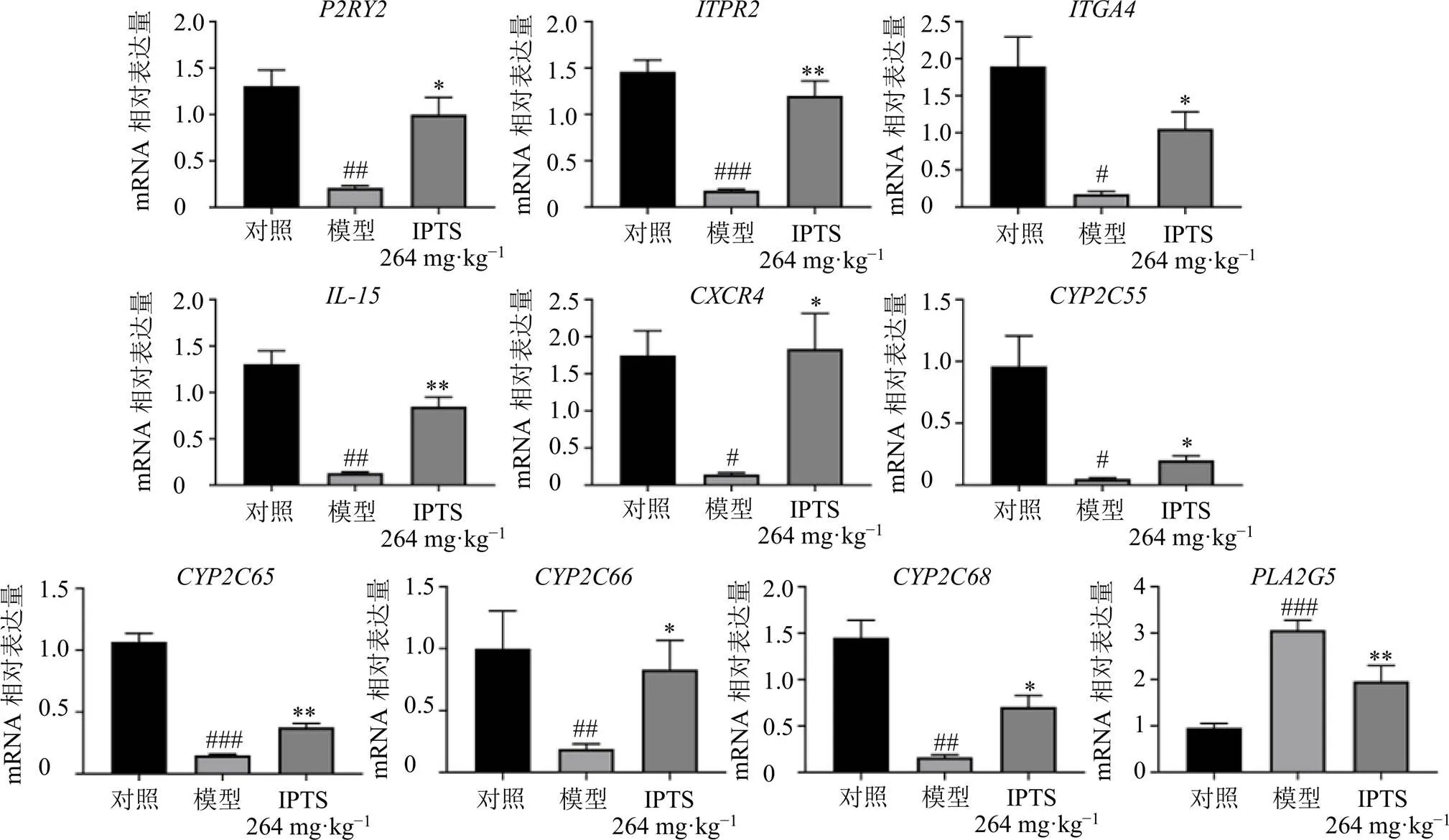
圖10 IPTS對腸損傷小鼠回腸組織中關鍵基因表達的影響(, n = 6)
4 討論
LPS是革蘭陰性菌細胞壁的一種成分,在腸屏障功能障礙中有著重要作用,LPS誘導的模型被廣泛認為是評價腸道損傷的代表性實驗模型[7]。在LPS刺激下,NF-κB信號通路被激活,釋放大量炎癥因子如TNF-α、IL-1β、IL-6等。腸道炎癥與腸屏障破壞有關,腸上皮炎癥可通過炎癥信號轉導下調緊密連接蛋白的表達[8]。腸道機械屏障是腸屏障中重要的組成部分,由單層的腸上皮細胞及細胞間的連接組成。其中,細胞間的連接方式包括緊密連接、黏附連接等,緊密連接是細胞間主要的連接方式。與緊密連接相關的蛋白有ZO蛋白家族、Occludin、Claudins蛋白家族等,這些蛋白的表達有助于腸屏障功能[9]。本研究結果顯示,IPTS可以下調腸損傷小鼠回腸組織中炎癥因子TNF-α、IL-1β的水平,上調回腸組織中緊密連接蛋白ZO-1、Occludin表達。
MDA是脂質過氧化的產物,被認為是過度氧化應激的指標。SOD可以催化O2?歧化為H2O2和O2,參與抗氧化系統。小腸過度氧化應激反應可以破壞脂質和蛋白質等重要細胞成分,進而導致腸屏障損傷[7]。本研究發現,IPTS降低腸損傷小鼠血清中MDA水平,升高SOD活性,表明IPTS可以抑制機體過度氧化應激反應,保護腸屏障。
本研究對對照組、模型組、IPTS高劑量組進行轉錄組學測序。KEGG通路富集分析結果顯示,DEGs主要富集在IgA相關的腸道免疫、花生四烯酸代謝、B細胞受體、TRP通道炎癥調控、亞油酸代謝等信號通路。IgA是黏膜免疫系統中含量最豐富的抗體亞型,分泌型免疫球蛋白A是由二聚體IgA、J鏈和分泌成分組成的聚合物免疫球蛋白,是機體黏膜局部抗感染免疫的主要抗體[10]。腸上皮中的分泌型免疫球蛋白A可以攔截入侵的病原體,防止病原體對腸道屏障的滲透,并將病原體從組織中排泄出來[11]。本研究選取IgA相關的腸道免疫通路上的、基因進行驗證。其中,IL-15是一種T細胞生長因子,可誘導生發中心B細胞增殖、分化和Ig合成[12]。趨化因子受體CXCR4是趨化因子基質細胞衍生因子12(C-X-C motif chemokine ligand 12,CXCL12)的特異受體。CXCL12對淋巴細胞有強烈的趨化作用[13]。因此,IPTS可能是通過調節其表達進而調節小腸免疫系統,維持腸屏障功能。
花生四烯酸是一種ω-6多不飽和脂肪酸。花生四烯酸代謝通路上的關鍵基因,具有將細胞膜內表面酯化的花生四烯酸水解成游離形式的作用,再由然后由環氧合酶、脂氧合酶和細胞色素P450酶進一步代謝成一系列生物活性介質,包括前列腺素、白三烯、二十碳四烯酸等,有助于細胞炎癥信號的傳遞,被認為是炎癥性疾病和心血管疾病的預防和治療靶點[14-15]。本研究結果顯示,IPTS可能是通過下調PLA2G5的表達,上調細胞色素P450家族(CYP2C55、CYP2C65、CYP2C66、CYP2C68)的表達,抑制小腸炎癥的發生。
此外,TRP通道是一種陽離子選擇性通道,廣泛存在于胃腸道,作為各種刺激的檢測器和G蛋白偶聯受體的二級轉導。TRP通道的激活可以引發相關神經肽的炎癥并啟動免疫反應,在控制單核細胞和巨噬細胞的吞噬、產生趨化因子和細胞因子等方面發揮重要作用[16],是炎癥通路中的重要信號成分[17]。該通路的關鍵基因屬于嘌呤能受體家族,能被三磷酸腺苷(adenosine triphosphate,ATP)或其他核苷酸激活。其中,P2Y受體主要為G蛋白偶聯受體,可以調節TRP通道。該通路另一關鍵基因是一種內質網鈣釋放通道,該基因編碼的蛋白家族成員是細胞內第2信使鈣釋放通道。因此,IPTS可能是通過調節P2RY2和ITPR2的表達,影響TRP通道,進一步抑制小腸炎癥反應。
綜上所述,本研究通過LPS誘導建立小鼠小腸損傷模型,通過檢測炎癥因子水平、腸屏障蛋白的表達等方法證明IPTS具有保護小腸腸屏障的作用。利用轉錄組學測序的手段,深入探究潛在機制,結果顯示IPTS通過介導IgA相關的腸道免疫信號通路、花生四烯酸代謝的信號通路、TRP通道炎癥調控通路和B細胞受體信號通路等多種途徑發揮保護作用。
利益沖突 所有作者均聲明不存在利益沖突
[1] 喬宛虹. 毛冬青的藥理作用及臨床應用研究概況 [J]. 中國現代藥物應用, 2008, 2(5): 104-106.
[2] 范星岳, 余繼英. 毛冬青活性成分及檢測方法的研究進展 [J]. 華西藥學雜志, 2019, 34(5): 512-518.
[3] 柳文媛, 李萍, 馮鋒, 等. 大鼠血漿中冬青素A的LC-MS測定法及其藥動學研究 [J]. 中國藥學, 2010,19(1): 38-42.
[4] 李美芬, 趙鐘祥, 林朝展, 等. 毛冬青皂苷元ilexgenin A在大鼠體內的藥動學研究 [J]. 華西藥學雜志, 2012, 27(3): 296-297.
[5] 白榮鈺, 易歡, 陳豐連, 等. 毛冬青三萜皂苷對動脈粥樣硬化大鼠腸道菌群的影響 [J]. 中草藥, 2021, 52(20): 6245-6253.
[6] Verhaar B J H, Prodan A, Nieuwdorp M,. Gut microbiota in hypertension and atherosclerosis: A review [J]., 2020, 12(10): E2982.
[7] Zhuang S, Zhong J, Bian Y F,. Rhein ameliorates lipopolysaccharide-induced intestinal barrier injury via modulation of Nrf2 and MAPKs [J]., 2019, 216: 168-175.
[8] Wu W C, Wang S S, Liu Q,. Metformin protects against LPS-induced intestinal barrier dysfunction by activating AMPK pathway [J]., 2018, 15(8): 3272-3284.
[9] Sánchez de Medina F, Romero-Calvo I, Mascaraque C,. Intestinal inflammation and mucosal barrier function [J]., 2014, 20(12): 2394-2404.
[10] Hand T W, Reboldi A. Production and function of immunoglobulin A [J]., 2021, 39: 695-718.
[11] Pietrzak B, Tomela K, Olejnik-Schmidt A,. Secretory IgA in intestinal mucosal secretions as an adaptive barrier against microbial cells [J]., 2020, 21(23): E9254.
[12] Patidar M, Yadav N, Dalai S K. Interleukin 15: A key cytokine for immunotherapy [J]., 2016, 31: 49-59.
[13] Mousavi A. CXCL12/CXCR4 signal transduction in diseases and its molecular approaches in targeted-therapy [J]., 2020, 217: 91-115.
[14] Wang B, Wu L J, Chen J,. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets [J]., 2021, 6(1): 94.
[15] Sztolsztener K, Chabowski A, Harasim-Symbor E,. Arachidonic acid as an early indicator of inflammation during non-alcoholic fatty liver disease development [J]., 2020, 10(8): 1133.
[16] Santoni G, Morelli M B, Amantini C,. “Immuno-transient receptor potential ion channels”: The role in monocyte-and macrophage-mediated inflammatory responses [J]., 2018, 9: 1273.
[17] Rajasekhar P, Poole D P, Veldhuis N A. Role of nonneuronal TRPV4 signaling in inflammatory processes [J]., 2017, 79: 117-139.
Effect and mechanism oftriterpenoid saponins on lipopolysaccharide-induced intestinal barrier injury in mice based on transcriptome
WANG Ying1, CHEN Bing-ying1, WU Yu-rong1, ZHANG Ying-yin1, WANG Qian1, LI Yu2, ZHANG Lei1
1. School of Pharmaceutical Science, Guangzhou University of Chinese Medicine, Guangzhou 510006, China 2. School of Nursing, Guangzhou University of Chinese Medicine, Guangzhou 510006, China
To investigate the effect and potential mechanism oftriterpenoid saponins (IPTS) on lipopolysaccharide (LPS)-induced small intestinal injury in mice.C57BL/6 mice were randomly divided into control group, model group and IPTS low-, medium-and high-dose (88, 176, 264 mg/kg) groups. After 7 d of drug intervention, mice were ip LPS to induce small intestine injury. After 7 h of modeling, hematoxylin eosin (HE) staining was used to observe pathology of ileum in mice; Level of malondialdehyde (MDA) and activity of superoxide dismutase (SOD) in serum of mice were detected; qRT-PCR and Western blotting were used to detect inflammatory factor interleukin-1β (), tumor necrosis factor-α () mRNA and intestinal barrier tight junction protein,mRNA and protein expressions in ileum. The potential mechanism of IPTS on ileum was explored by transcriptome sequencing.Compared with model group, ileum in IPTS group was intact, oxidative stress injury was improved (< 0.001), level of ileitis factor was decreased (< 0.01), and intestinal barrier was restored (< 0.05, 0.01, 0.001). RNA-seq sequencing results showed that 2584 differentially expressed genes were screened from model group compared with control group; Compared with model group, 259 differentially expressed genes were screened out in IPTS high-dose group; These differentially expressed genes were mainly enriched in signaling pathway such as immunoglobulin A (IgA) related intestinal immunity, arachidonic acid metabolism, B cell receptor, transient receptor potential (TRP) channel inflammation regulation, linoleic acid metabolism, etc.IPTS can improve LPS induced intestinal injury in mice, which may be closely related to arachidonic acid metabolism, IgA related intestinal immune metabolism and TRP channel inflammatory regulation pathway.
triterpene saponins; intestinal barrier injury; transcriptome; oxidative stress; inflammation; intestinal barrier
R285.5
A
0253 - 2670(2022)22 - 7102 - 10
10.7501/j.issn.0253-2670.2022.22.013
2022-08-16
廣東省科技計劃項目(2016A020226031);廣東省自然科學基金資助項目(2018A030313328);廣東省教育廳創新強校工程項目(2016KTSCX019)
王 瑩(1998—),女,碩士研究生,研究方向為中藥藥效物質基礎及其作用機制。Tel: 13128253065 E-mail: 839072446@qq.com
張 蕾,女,碩士生導師,教授,研究方向為中藥藥效物質基礎及其作用機制。Tel: 15622182155 E-mail: zhangleic431@163.com
[責任編輯 李亞楠]
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
數學物理學報(2020年2期)2020-06-02 11:29:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
