SLC39A13基因的泛癌分析
2022-11-08 14:09:20陳暢楊淑雅劉寶怡李冕魯艷芹
中國醫學工程 2022年10期
陳暢,楊淑雅,劉寶怡,李冕,魯艷芹,2
[1.山東第一醫科大學(山東省醫學科學院)生物醫學科學學院(省醫藥生物技術研究中心),山東 濟南 250117;2.山東第一醫科大學第一附屬醫院 內分泌科,山東 濟南250014]
SLC39A13基因位于11p11.2 染色體處,編碼ZIP13 蛋白,在維持細胞內外鋅離子平衡中起重要作用[1]。研究發現ZIP13 在結締組織發育中有關鍵作用,SLC39A13的異常突變會引起一種結締組織病脊柱發育不良埃勒斯-丹洛斯綜合征3 型(spondylo-dysplastic ehlers-danlos syndrome type 3)[1-2]。ZIP 家族作為重要的鋅轉運體在癌癥的發生發展中起重要作用。目前已有文獻證實ZIP 家族相關基因與癌癥有關,例如ZIP4 的高表達與胃癌和胰腺癌的惡化有關[3];胰腺癌早期發生RREB1/ZIP3 的下調導致鋅的減少[4]。但目前僅有1 篇文章指出SLC39A13/ZIP13 的高表達與卵巢癌的轉移、預后差有關[1]。尚未有SLC39A13的泛癌研究,因此本文首次對TCGA 和GEO 數據庫進行深入挖掘,研究SLC39A13在不同癌癥發病機制中的分子機制或臨床預后中的作用。
1 材料與方法
1.1 基因表達分析
在TIMER2.0 網站(http://timer.cistrome.org/)中輸入“SLC39A13”,比較SLC39A13在TCGA 數據庫中不同腫瘤(或特定腫瘤亞型)和相鄰正常組織中的表達差異。對于某些正常組織缺乏的腫瘤,利用GEPIA2(http://gepia2.cancer-pku.cn/#index)的“表達分析-框圖”模塊分析,來獲得這些腫瘤組織與GTEx 數據庫中對應的正常組織表達差異的數據,設置“P值<0.01,log2FC<1”和“輸入缺少正常組織對照的腫瘤”。此外,通過GEPIA2 網站的Stage Plot 模塊可以得到SLC39A13在TCGA 數據庫中腫瘤不同病理階段的表達情況,用log2(TPM+1)轉換后的數據作出“小提琴圖譜”。
1.2 生存預后分析
使用GEPIA2 的“生存分析”模塊獲得所有TCGA 數據庫中涉及SLC39A13基因的腫瘤總生存期(OS)和無病生存期(DFS)的顯著差異性圖,設置參數“Cutoff-high(50%),Cutoff-low(50%)”來區分高表達組和低表達組。假設檢驗采用對數秩和檢驗,通過GEPIA2 的“生存分析”模塊繪制生存圖。
1.3 基因變異分析
利用cBioPortal 網站(http://www.cbioportal.org/),在“快速選擇”區選擇“TGCA 泛癌圖譜”研究,查詢SLC39A13基因的遺傳變異情況。在“癌癥類型匯總”模塊中觀察所有TCGA 腫瘤的突變頻率、突變類型和拷貝數的改變值(copy number alteration,CNA)。SLC39A13的突變位點信息可以通過“突變”模塊顯示在蛋白質結構示意圖或3D 結構中。利用“比較”模塊來獲得TCGA癌癥病例在有或沒有SLC39A13基因突變時的總生存、無疾病、無進展和無疾病生存差異的數據,同時生成P 值為log-rank 的Kaplan-Meier圖。
1.4 免疫浸潤分析
使用TIMER2.0 數據庫的“免疫基因”模塊來探索所有TCGA 腫瘤中SLC39A13表達與免疫浸潤之間的關系。選擇CD8+T 細胞作為癌相關成纖維細胞的免疫細胞。應用 CIBERSOR-ABS、QUANTISEO、EXCEL、MCPCOUNTER 和EPIC 算法進行免疫浸潤評估。根據純度調整后的Spearman's 等級相關實驗獲得P值和偏相關系數(COR)。數據被可視化為熱圖和散點圖。
1.5 SLC39A13 編碼蛋白ZIP13 的相互作用蛋白富集分析
首先使用STRING 網站(https://string-db.org/),查詢單一蛋白質名稱(SLC39A13/ZIP13)和物種(人類),隨后設置以下主要參數:所需的最低交互分數(低信度0.150);線條顏色表示交互證據的類型;最大交互的數量(不超過50)和來源(實驗證實)。最后篩選出10 個經實驗證實的與ZIP13 結合的蛋白。使用GEPIA2 的“相似基因檢測”模塊,基于所有TCGA 腫瘤和正常組織的數據集,獲得了前100 個SLC39A13相關的靶向基因。應用GEPIA2 的“相關分析”模塊進行相似基因與SLC39A13基因的相關性分析。應用log2TPM 方塊圖,得到P 值和相關系數R。此外,使用TIMER2 的“基因相關”模塊作出所用基因的數據熱圖。使用Venn 網站(http://bioinformatics.psb.ugent.be/webtools/Venn/)來進行交叉分析從而比較SLC39A13結合基因和相似基因。結合這兩組數據進行KEGG 路徑分析。將基因列表上傳到David 網站(https://david.ncifcrf.gov/)并選擇“OFFICIAL_GENE_SYMBOL” 和“Homo Spaiens”分別作為基因標識符和物種,獲得了功能注釋圖的數據。最后將P<0.05 的結果帶入R 包進行GO 富集分析。
1.6 統計學方法
以P<0.05 為差異有統計學意義。
2 結果
2.1 基因表達數據分析
應用TIMER2.0 方法分析了TCGA 數據庫的多種癌癥類型中SLC39A13的表達狀態。在乳腺癌(BRCA),結腸癌(COAD),頭頸鱗狀細胞癌(HNSC),肝癌(LIHC),胃癌(STAD)組織中SLC39A13的表達水平高于相應的正常組織,差異有統計學意義(P<0.001);宮頸癌(CESC)組織中SLC39A13的表達水平低于相應的正常組織,差異有統計學意義(P<0.01),見圖1A。
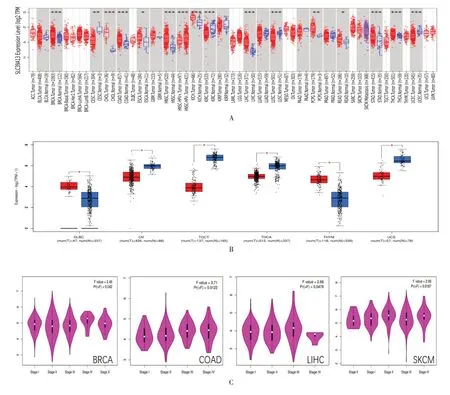
圖1 SLC39A13 在不同腫瘤及腫瘤分期中的基因表達分析
將GTEx 數據集的正常組織作為對照,進一步評估SLC39A13在正常組織和腫瘤組織之間的表達差異,在彌漫性大B 細胞淋巴瘤(DLBC),胸腺癌(THYM)組織中,SLC39A13表達高于相應正常組織。在卵巢癌(OV),睪丸癌(TGCT),甲狀腺癌(THCA),子宮肉瘤(UCS)組織中,SLC39A13表達低于相應正常組織,見圖1B。使用GEPIA2 的“病理階段”模塊觀察SLC39A13的表達與癌癥病理階段的相關性,包括乳腺癌,結腸癌,肝癌和皮膚黑色素瘤(SKCM),在這些腫瘤組織中SLC39A13表達普遍呈現上升趨勢,差異有統計學意義(均P<0.05),見圖1C。
2.2 生存數據分析
根據SLC39A13的表達水平,將癌癥病例分為高表達組和低表達組,研究了SLC39A13的表達與不同腫瘤患者預后的相關性。高表達的SLC39A13與宮頸癌(P=0.027),結腸癌(P=0.023),多形性膠質母細胞瘤(GBM)(P=0.002),腦低級別膠質瘤(LGG)(P=0.003),間皮瘤(MESO)(P=0.009)總體預后不良有關。SLC39A13低表達與胰腺癌(PAAD)(P=0.015)總體預后不良有關,見圖2A。無病生存率分析數據顯示SLC39A13的高表達與頭頸鱗狀細胞癌(P=0.044)、腦低級別膠質瘤(P=0.009)、肝癌(P=0.021)、肺鱗狀細胞癌(LUSC)(P=0.011)、間皮瘤(P=0.040)、前列腺癌(PRAD)(P<0.001)患者預后差有關,見圖2B。以上數據表明SLC39A13的表達高低與不同類型腫瘤的預后存在差異。
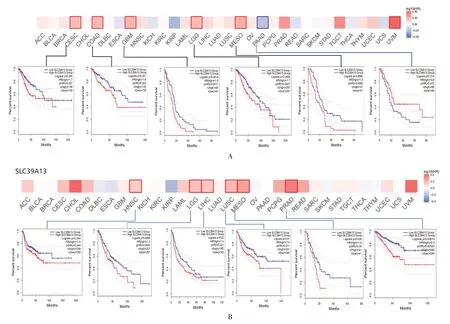
圖2 SLC39A13 基因與腫瘤生存預后的關系
2.3 基因變異分析
以SLC39A13突變為主要變異類型的子宮內膜癌(UCEC)患者SLC39A13基因變異頻率最高(>3%)。皮膚黑色素瘤患者僅發生SLC39A13突變,其變異頻率>2%。SLC39A13擴增是腎上腺皮質癌(ACC)和頭頸鱗狀細胞癌患者的唯一類型,變異頻率分別為1.1%和0.96%,見圖3A。進一步顯示SLC39A13遺傳變異的類型和位點,見圖3B,發現SLC39A13錯義突變是遺傳變異的主要類型。此外,還探討了SLC39A13的基因突變與不同類型癌癥患者臨床預后之間在潛在聯系,見圖3C。與無SLC39A13基因突變的病例相比,SLC39A13突變后與子宮內膜癌患者的OS(P=0.252)、DFS(P=0.267)、疾病特異性生存期(DSS)(P=0.164)無明顯相關,與無進展生存期(PFS)(P=0.044)有關,見圖3C。
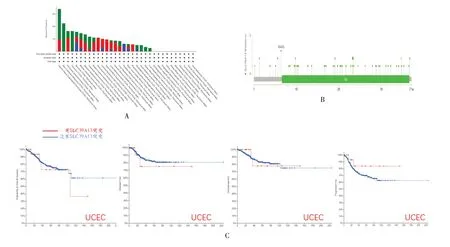
圖3 SLC39A13 基因突變的分析
2.4 免疫浸潤數據分析
腫瘤浸潤性免疫細胞,作為腫瘤微環境的重要成分,與癌癥的啟動、進展和轉移密切相關[5-6]。據報道,腫瘤微環境基質中的癌癥相關成纖維細胞參與調節各種腫瘤浸潤性免疫細胞的功能[5-6]。在此,使用一些算法來挖掘免疫細胞浸潤水平和SLC39A13基因在TCGA 數據庫中不同癌癥類型中的表達水平之間的潛在關系。發現SLC39A13的表達與免疫浸潤呈正相關的腫瘤有結腸癌、食管癌、胰腺癌、直腸癌、胃癌、睪丸癌,SLC39A13的表達水平與腎乳頭狀細胞癌(KIRP)相關成纖維細胞浸潤水平呈負相關,見圖4。
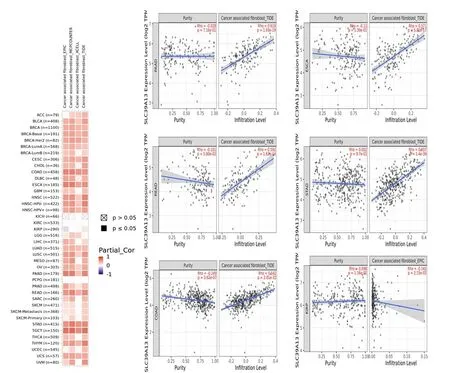
圖4 SLC39A13 基因的免疫浸潤富集分析
2.5 ZIP13 的相互作用蛋白分析
為了進一步研究SLC39A13基因在腫瘤發生中的分子機制,篩選出SLC39A13編碼蛋白ZIP13 相互作用蛋白,基于STRING 工具,總共獲得了10個ZIP13 結合蛋白的實驗數據。圖5A 顯示了這些蛋白質相互作用的網絡。使用GEPIA2 數據庫結合TCGA 所有腫瘤的表達數據,獲得了與SLC39A13表達相關的前100 個基因。圖5B 所示,SLC39A13的表達水平與FAM65A、VEGFB、PLA2G15、GLIS2基因(均P<0.001)呈正相關。相應的熱圖數據也顯示,SLC39A13與上述的4 個癌癥類型的基因呈正相關,見圖5C。兩組數據的交叉分析顯示沒有共同基因,見圖5D。
結合兩組數據進行KEGG 和GO 的富集分析。KEGG 數據顯示出SLC39A13可能通過參與“代謝途徑”和“溶酶體”通路進一步影響腫瘤的發生。GO 富集分析數據進一步表明,這些基因中的大多與酶活動和離子轉運密切相關,見圖5E。
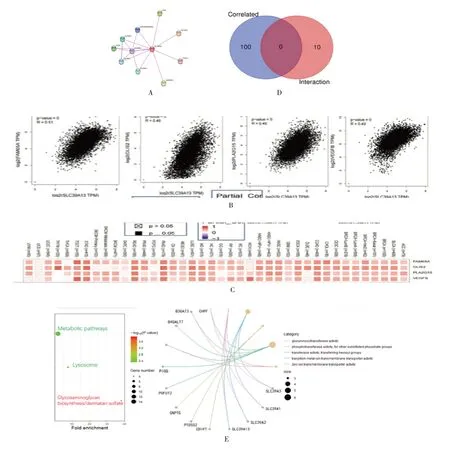
圖5 SLC39A13 編碼蛋白ZIP13 相互作用蛋白及其基因富集分析與腫瘤相關性分析
3 討論
目前尚無從整體角度對SLC39A13進行泛癌分析的文獻。本文首次系統地通過生信分析發現SLC39A13在乳腺癌、結腸癌、卵巢癌等腫瘤組織中高表達。已有文獻證實在卵巢癌中SLC39A13/ZIP13 是一種重要的腫瘤干細胞調節因子,在卵巢癌組織中過度表達,通過Src/FAK 信號通路調控轉移相關基因,促進卵巢癌的轉移[1]。這與本文分析結果一致。SLC39A7基因是乳腺癌細胞生存的關鍵基因,是乳腺癌診斷和預后的重要分子標志物[7]。生信分析發現SLC39A13在乳腺癌組織中高表達,可能作為潛在的表乳腺癌預后分子標志物。高表達的SLC39A13基因與結腸癌的預后差相關。結腸癌曾是全球第五大癌癥死亡原因[8],而根據最新數據預計2021 年結腸癌將成為全球癌癥死亡的主要原因[9]。結腸癌也是我國最常見的消化道惡性腫瘤之一[10],大約四分之一的患者在初次診斷時就發現有遠處轉移,導致預后較差[11]。基質金屬蛋白酶(MMP)參與結腸癌的局部和遠端侵襲,血漿中MMP 高表達與結腸癌預后差相關[12]。據報道,EMT 相關基因的表達與T 細胞的抗腫瘤免疫相關。EMT 基因的高表達與結腸癌的預后差有關,而浸潤性T 細胞豐度(ITA)越高與結腸癌的預后好相關[13]。本文發現SLC39A13基因在結腸癌患者中的異常表達也有可能作為結腸癌診斷和預后分析的分子標志物。宮頸癌組織中SLC39A13的表達低于正常組織,其高表達與患者預后差有關。宮頸癌是世界上第四大常見的惡性腫瘤,每年新增超過52 萬例,其中超過85%發生在發展中國家[14]。鈣活化氯離子通道2(CLCA2)表達的降低可能是宮頸癌患者的一個潛在預后標志物,但其分子機制還有待進一步研究[15]。前膠原賴氨酸,2-酮戊二酸5-雙加氧酶(PLOD)是一種有效的預后標志物,與宮頸癌的免疫浸潤有關[16]。SLC39A13基因在宮頸癌患者中表達情況也可以作為一個診斷和治療的輔助標志物。在胰腺癌組織中SLC39A13的表達量低于正常組織,其低表達與患者預后差有關。根據美國癌癥協會2019年公布的數據,胰腺癌引起的死亡率在惡性腫瘤中排名第四[17-18]。雖然在了解胰腺癌的分子發病機制方面取得了重大進展,但胰腺癌患者的預后仍然非常差,過去20 年的5 年生存率僅從5%增加到8%。胰腺癌的新分子預后標志物和潛在治療靶點仍是未來研究方向[19]。臨床和實驗證據顯示與正常/良性胰腺組織相比,人胰腺癌組織中鋅的含量持續降低[4]。胰腺癌患者血清鋅濃度降低表明,鋅和鋅轉運相關基因可能作為這種惡性疾病的生物標志物或治療靶點。鋅穩態受到細胞中SLC39A/ZIP、SLC30A/ZnT 和金屬硫蛋白(MT)的調控。鋅穩態失調可能影響癌細胞的異常特征[20]。鋅轉運體ZIP4、ZIP11、ZnT 和ZnT6 的表達增加預示著胰腺癌的預后不良[20]。生信分析表明ZIP13 的表達降低預示著胰腺癌的預后不良,這為理解鋅穩態與胰腺癌之間的復雜關系提供了新的線索。鋅穩態相關基因在胰腺癌中的預后和治療作用機制值得進一步探索[20]。SLC39A13表達量的不同對不同類型的腫瘤影響是不一樣的,在臨床中,還需要考慮到其他臨床特征,通過更深入的分子實驗證據來確定SLC39A13的表達是否與上述腫瘤的發生發展有關。
SLC39A13最主要的變異類型是錯義突變。子宮內膜瘤患者中發生該基因的錯義突變和擴增。子宮內膜瘤是一種遺傳性疾病,通常伴有基因突變或異常表達[21]。分析發現SLC39A13的擴增與患者的預后差相關,因此SLC39A13基因突變也有可能是子宮內膜瘤患者預后差的潛在分子標志物。SLC39A13表達與免疫浸潤呈正相關的腫瘤有結腸癌、食管癌、胰腺癌、直腸癌、胃癌和睪丸癌,SLC39A13表達水平與腎乳頭狀細胞癌相關成纖維細胞浸潤水平呈負相關。腫瘤免疫浸潤細胞從血液遷移到腫瘤組織,并在免疫調節中起重要作用[22]。越來越多研究表明,腫瘤免疫浸潤細胞與免疫檢查點抑制和預后密切相關[22]。SLC39A13基因與免疫浸潤的關系有望為免疫治療提供新思路。
綜上所述,SLC39A13的泛癌分析表明其表達與臨床預后、基因突變、腫瘤細胞的免疫浸潤之間存在相關性,這有助于從多因素綜合分析SLC39A13在腫瘤發生發展中的作用,為臨床診斷治療及基礎研究提供幫助。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
西南國防醫藥(2016年7期)2016-12-01 06:01:15
山東工業技術(2016年15期)2016-12-01 05:31:22
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
河北醫科大學學報(2011年6期)2011-03-25 10:16:48
