84個(gè)蘋(píng)果栽培品種對(duì)斑點(diǎn)落葉病的抗性評(píng)價(jià)和全基因組關(guān)聯(lián)分析
2022-10-28 10:25:14儲(chǔ)寶華曹富國(guó)卞寧寧錢(qián)謙李中興李雪薇劉澤遠(yuǎn)馬鋒旺管清美
中國(guó)農(nóng)業(yè)科學(xué) 2022年18期
關(guān)鍵詞:關(guān)聯(lián)
儲(chǔ)寶華,曹富國(guó),卞寧寧,錢(qián)謙,李中興,李雪薇,劉澤遠(yuǎn),馬鋒旺,管清美
西北農(nóng)林科技大學(xué)園藝學(xué)院,陜西楊凌 712100
0 引言
【研究意義】中國(guó)是世界上蘋(píng)果的生產(chǎn)和出口大國(guó)。自2007年以來(lái),中國(guó)每年的蘋(píng)果總產(chǎn)量均占世界總產(chǎn)量的45%以上,2019年中國(guó)的蘋(píng)果產(chǎn)量甚至達(dá)到4 100萬(wàn)t,占世界蘋(píng)果總產(chǎn)量的54.07%(數(shù)據(jù)來(lái)源于蘋(píng)果產(chǎn)業(yè)大數(shù)據(jù)分析平臺(tái)(agdata.cn))。早期落葉病是威脅中國(guó)蘋(píng)果生產(chǎn)的主要病害之一,包括褐斑病、斑點(diǎn)落葉病、灰斑病、圓斑病、輪斑病等多種病害,其中斑點(diǎn)落葉病和褐斑病的危害最為嚴(yán)重。斑點(diǎn)落葉病主要通過(guò)危害葉片影響樹(shù)體生長(zhǎng),進(jìn)而影響蘋(píng)果的產(chǎn)量和品質(zhì)。感染斑點(diǎn)落葉病后,蘋(píng)果葉片表面產(chǎn)生褐色斑點(diǎn),隨后黃化脫落,造成樹(shù)勢(shì)早衰;此外,斑點(diǎn)落葉病也會(huì)感染幼果從而引發(fā)果實(shí)腐爛。斑點(diǎn)落葉病嚴(yán)重的果園中葉片感染率能夠達(dá)到90%左右,落葉率達(dá)到80%,給蘋(píng)果產(chǎn)業(yè)造成極大的經(jīng)濟(jì)損害。目前關(guān)于蘋(píng)果栽培品種對(duì)早期落葉病的抗性研究多集中于褐斑病、腐爛病、枝干輪紋病方面,關(guān)于斑點(diǎn)落葉病在蘋(píng)果生產(chǎn)中的研究則集中于農(nóng)業(yè)防治方面,且關(guān)于不同栽培品種對(duì)斑點(diǎn)落葉病抗性系統(tǒng)評(píng)價(jià)的相關(guān)研究還較少。目前鑒定到的高抗性蘋(píng)果栽培品種數(shù)量稀少,因此急需挖掘具有高抗病性的蘋(píng)果栽培品種以滿足市場(chǎng)的栽培需求。通過(guò)比較國(guó)內(nèi)外蘋(píng)果栽培品種對(duì)斑點(diǎn)落葉病的抗性,從而全面系統(tǒng)地認(rèn)識(shí)不同栽培品種的抗病能力,篩選出穩(wěn)定的抗性品種,是解決蘋(píng)果園受斑點(diǎn)病侵害的有效途徑。【前人研究進(jìn)展】全基因組關(guān)聯(lián)性狀分析(genome-wide association study,GWAS)是一種用于開(kāi)展連鎖標(biāo)記開(kāi)發(fā)和關(guān)鍵調(diào)控基因挖掘的有效方法,目前在各類(lèi)糧食作物如水稻、小麥、玉米上廣泛應(yīng)用。PAULINO 等通過(guò)GWAS分析發(fā)現(xiàn)抗枯萎病相關(guān)的關(guān)鍵 SNP位點(diǎn)和關(guān)鍵基因;YANG等整合多種分析方法,如Meta-QTL、GWAS和同源性分析,揭示了一個(gè)新的調(diào)控小麥粒徑大小的關(guān)鍵基因—。目前GWAS在蘋(píng)果上的應(yīng)用還很少,且現(xiàn)有的研究主要集中在果實(shí)品質(zhì)方面。如FARNETI等將基因組與蘋(píng)果揮發(fā)性物質(zhì)相關(guān)聯(lián),鑒定到9 142個(gè)SNP和兩個(gè)與芳香性物質(zhì)緊密相關(guān)的候選基因和;LIAO等也利用 GWAS發(fā)現(xiàn)與蘋(píng)果果實(shí)酸度緊密關(guān)聯(lián)的基因和。【本研究切入點(diǎn)】利用全基因組關(guān)聯(lián)分析挖掘蘋(píng)果抗斑點(diǎn)落葉病的單核苷酸多態(tài)性位點(diǎn)(SNP)及關(guān)鍵候選基因。【擬解決的關(guān)鍵問(wèn)題】本研究材料的選擇基于課題組現(xiàn)有的蘋(píng)果栽培品種 84份,通過(guò)離體葉片接種病原菌的試驗(yàn)評(píng)價(jià)斑點(diǎn)落葉病抗性,綜合多次試驗(yàn)結(jié)果分析病斑面積、病斑面積增長(zhǎng)率,結(jié)合聚類(lèi)分析的手段,篩選出蘋(píng)果栽培品種中穩(wěn)定的抗性品種。
1 材料與方法
1.1 供試材料
試驗(yàn)材料栽植于蘋(píng)果資源圃。蘋(píng)果栽培品種共84種,2018年4月將84個(gè)栽培品種采用嵌芽接的方式嫁接于砧木‘平邑甜茶’,每個(gè)品種均有5個(gè)生物學(xué)重復(fù)。試驗(yàn)分別于2019和2020年進(jìn)行,于8月中旬采集植物離體葉片,每個(gè)品種5個(gè)生物學(xué)重復(fù),每個(gè)生物學(xué)重復(fù)中采取植株中部健康完整的成熟葉片3—5片。
1.2 接種方法
試驗(yàn)所用的蘋(píng)果斑點(diǎn)落葉病病菌(f. sp.)來(lái)源于西北農(nóng)林科技大學(xué)孫麗英教授實(shí)驗(yàn)室,菌種保存,擴(kuò)繁的方式參考ZHANG等。具體接種方法如下:病菌活化培養(yǎng)4 d后,切取菌塊放置在PDA平板中央,黑布遮光,25℃培養(yǎng)4 d,待菌落直徑長(zhǎng)到2 cm時(shí),用5 mm打孔器取菌落周?chē)麻L(zhǎng)出的白色菌絲用于葉片接種。接種時(shí)將葉片清洗干凈,葉柄用濕棉花包裹以防葉片失水,用手術(shù)刀刀尖在葉片正面的葉脈左右兩側(cè)中部刺傷,于傷口處放置菌塊。接完種的葉片放于托盤(pán)中,覆蓋保鮮膜,黑布遮光培養(yǎng),于25℃培養(yǎng)4 d和8 d后分別拍照,統(tǒng)計(jì)葉背面病斑面積。
1.3 抗性評(píng)價(jià)的方法
統(tǒng)計(jì)接種后4 d和8 d葉背面的病斑面積大小,病斑面積越大,說(shuō)明植物的抗病能力越弱。此外,通過(guò)4—8 d病斑面積增長(zhǎng)率,來(lái)判斷品種抗擴(kuò)展能力。病斑增長(zhǎng)率計(jì)算公式:

1.4 全基因組關(guān)聯(lián)分析
使用高效混合模型關(guān)聯(lián)程序EMMAX進(jìn)行關(guān)聯(lián)分析,截止點(diǎn)=0.0000001(-LgP=7),進(jìn)行關(guān)聯(lián)位點(diǎn)的檢測(cè)。本研究利用全基因組深度測(cè)序獲得的1 243 071個(gè)高質(zhì)量SNP位點(diǎn),用EMMAX-Kinship程序計(jì)算,從所有的SNP中得到親緣關(guān)系矩陣。基于以上SNP信息,用GCTA(1.01版)程序?qū)μO(píng)果84個(gè)栽培品種進(jìn)行主成分分析并獲得前 10個(gè)主成分矩陣。最后,將84份蘋(píng)果栽培種接種后葉片的病斑面積數(shù)據(jù)作為表型,全基因組深度重測(cè)序獲得的1 243 071個(gè)高質(zhì)量SNP位點(diǎn)為遺傳標(biāo)記,結(jié)合親緣關(guān)系矩陣和主成分分析結(jié)果,采用EMMAX方法進(jìn)行全基因組關(guān)聯(lián)性狀分析。
1.5 候選基因注釋
用ANNOVAR軟件,結(jié)合蘋(píng)果參考基因組GDDH13(https://iris.angers.inra.fr/gddh13/)對(duì)全基因組關(guān)聯(lián)分析得到的SNP進(jìn)行注釋?zhuān)@得每個(gè)SNP上下游約50 kb的突變位點(diǎn)和SNP位點(diǎn)相鄰基因的MD號(hào)。使用NCBI-Blast-2.9.0+中 Blastp和擬南芥蛋白組 Tair10(https://www.arabidopsis.org/)進(jìn)行親緣關(guān)系比對(duì)。用Python3.7版對(duì)數(shù)據(jù)進(jìn)行處理,獲得SNP相鄰候選基因的注釋列表。
1.6 數(shù)據(jù)統(tǒng)計(jì)及分析
病斑面積用 Image J軟件統(tǒng)計(jì)分析,使用 Excel計(jì)算平均值和標(biāo)準(zhǔn)差,用SPSS分別對(duì)2020年和2021年的病斑面積和病斑面積增長(zhǎng)率進(jìn)行聚類(lèi)分析,聚類(lèi)分析方法采用瓦爾德法,聚類(lèi)距離采用平方歐式距離。
1.7 基因表達(dá)分析
根據(jù)GDR網(wǎng)站上、、、、、、的mRNA 序列(https://www.rosaceae.org/),在 NCBI網(wǎng)頁(yè)上設(shè)計(jì)熒光定量PCR引物(表1),將接種后的GL-3組培苗葉片樣品在液氮中充分研磨,用Wolact?Plant RNA Isolation Kit(Hong Kong,China)試劑盒進(jìn)行 RNA 提取, 使用HiScript Ⅱ Q RT SuperMix for qPCR (+gDNA wiper)進(jìn)行反轉(zhuǎn)錄,定量試劑用ChamQ SYBR Qpcr Master Mix(Q311-02/ 03),反應(yīng)體系參考試劑使用說(shuō)明。使用Bio-RadCFX96TM(Bio-Rad)熒光定量 PCR儀進(jìn)行反應(yīng),以接種 0、12、24和 48 h的葉片樣品為模板進(jìn)行基因的相對(duì)表達(dá)量分析,用作內(nèi)參基因,使用2計(jì)算基因的相對(duì)表達(dá)水平。

表1 引物序列Table 1 Primer sequence
1.8 蘋(píng)果葉片的瞬時(shí)轉(zhuǎn)化
將的基因編碼區(qū)(CDS區(qū))進(jìn)行克隆后構(gòu)建到入門(mén)載體pDONR222,通過(guò)LR反應(yīng)構(gòu)建至表達(dá)載體 PK7-203,引物設(shè)計(jì)見(jiàn)表1,構(gòu)建好的表達(dá)載體轉(zhuǎn)入農(nóng)桿菌C58C1中,通過(guò)農(nóng)桿菌介導(dǎo)的蘋(píng)果葉片瞬時(shí)表達(dá)系統(tǒng)。試驗(yàn)方法參考ZHANG等,具體步驟如下:將 100 μL 濃度為 100 μmol·L的乙酰丁香酮加入100 mL LB中,加入100 μL的載體轉(zhuǎn)化農(nóng)桿菌,在28℃培養(yǎng)箱,130 r/min振蕩培養(yǎng)12—16 h,直至OD=1,5 000 r/min離心10 min。將農(nóng)桿菌懸浮于100 mL液體培養(yǎng)基(10 mmol·LMES-KOH(pH 5.2)、10 mmol·LMgCl和 180 μmol·L乙酰丁香酮)中,使用前培養(yǎng)2 h。利用4周大的組培苗進(jìn)行抽真空,真空壓力為0.06—0.08 Mpa,抽真空時(shí)間約4—5 h,直至整個(gè)葉片出現(xiàn)水漬狀,之后將組培苗放回MS培養(yǎng)基進(jìn)行培養(yǎng)(25±1℃,白天/黑夜16 h/8 h)。在農(nóng)桿菌侵染4 d后進(jìn)行離體葉片接種試驗(yàn),接種3 d后進(jìn)行病斑面積統(tǒng)計(jì)。
2 結(jié)果
不同類(lèi)型的蘋(píng)果屬栽培品種在接種斑點(diǎn)落葉病原菌后,其葉面受侵染的嚴(yán)重度表現(xiàn)出顯著多樣性。為了系統(tǒng)評(píng)價(jià)不同栽培品種對(duì)斑點(diǎn)落葉病的抗性差異,本研究將葉片壞死的病斑面積作為評(píng)價(jià)不同蘋(píng)果屬栽培品種對(duì)斑點(diǎn)落葉病抗性的標(biāo)準(zhǔn),病斑面積越大,則植物的抗病能力越差;反之,病斑面積越小,則植物的抗病能力越強(qiáng)。由于單次試驗(yàn)結(jié)果具有片面性,本研究連續(xù)2年統(tǒng)計(jì)了4次試驗(yàn)結(jié)果(2次8 d結(jié)果,2次4 d結(jié)果),最終選擇4次數(shù)據(jù)評(píng)價(jià)穩(wěn)定的品種確定為抗性或感病品種。因此,本結(jié)果能夠更客觀地系統(tǒng)評(píng)價(jià) 84個(gè)蘋(píng)果栽培品種的抗病能力。此外,利用全基因組關(guān)聯(lián)性狀分析的手段鑒定抗病性相關(guān)的關(guān)鍵 SNP位點(diǎn),并篩選關(guān)鍵的潛在調(diào)控基因。
2.1 84個(gè)蘋(píng)果栽培品種的抗病性差異比較(2020年)
84個(gè)蘋(píng)果栽培品種的離體葉片在接種斑點(diǎn)落葉病菌株后均有不同程度的發(fā)病(表2),目前未發(fā)現(xiàn)完全免疫的品種。2020年接種8 d后的病斑面積數(shù)據(jù)顯示,病斑面積的分布范圍在 0.76—6.02 cm,‘凱密歐’的病斑面積最小,僅為(0.76±0.14)cm;而‘Chanterler’的病斑面積最大,達(dá)到(6.02±0.23)cm。另外,為了進(jìn)一步衡量品種抗擴(kuò)展能力,進(jìn)行了4—8 d內(nèi)的病斑面積增長(zhǎng)率的分析。數(shù)據(jù)顯示,‘凱密歐’病斑面積增長(zhǎng)率最低,僅為15.00%;而病斑面積增長(zhǎng)率最高的‘埃德?tīng)柌┧苟喾颉_(dá) 128.25%。綜合病斑面積和病斑增長(zhǎng)率兩類(lèi)指標(biāo),進(jìn)行聚類(lèi)分析(圖1-a)。84個(gè)栽培品種中,‘凱密歐’‘Kiku’‘JA’‘皇家嘎啦’‘秋光’‘蜜脆’‘藍(lán)皮爾曼’‘福拉瑞娜’‘Roho3615’‘艾達(dá)紅’‘威廉姆斯女士’‘弘前富士’‘Fujiko’‘紅蓋露’‘Gloster69’‘長(zhǎng)富2號(hào)’‘早紅1號(hào)’聚為一類(lèi),為抗病品種;‘Chanterler’‘埃德?tīng)柌┧苟喾颉鹊俑窈D贰艥饧t’‘赫拉森’‘阿麗亞娜’‘2001富士’聚為一類(lèi),為易感品種。
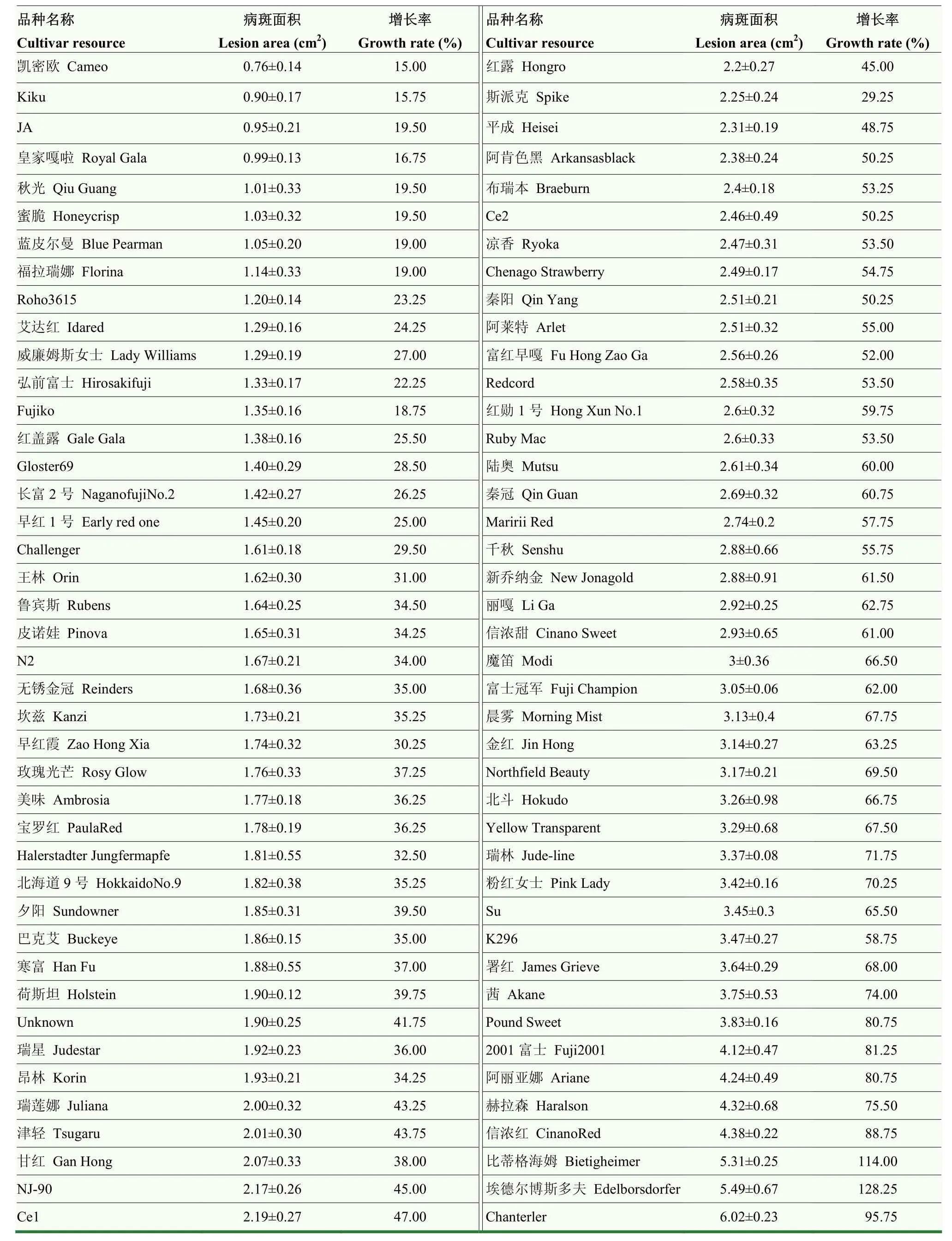
表2 84個(gè)蘋(píng)果屬栽培品種離體葉片接種病斑大小及病斑面積增長(zhǎng)率(2020年)Table 2 Lesion area and its growth rate of 84 apple cultivars inoculated with Alternaria alternata f. sp. Mali
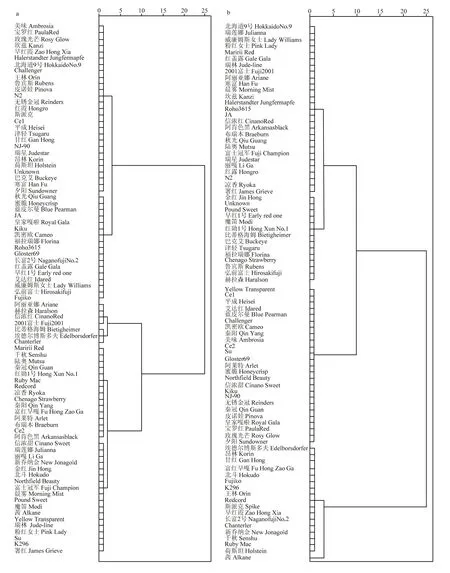
圖1 2020年(a)和2021年(b)84個(gè)栽培品種對(duì)斑點(diǎn)落葉病抗性的聚類(lèi)分析Fig. 1 Cluster analysis of resistance of 84 cultivars to Alternaria alternate f. sp. mali in 2020 (a) and 2021 (b)
2.2 不同類(lèi)型的栽培品種的抗病性差異比較(2021年)
為了進(jìn)一步證明試驗(yàn)結(jié)果的可靠性,于2021年進(jìn)行了重復(fù)試驗(yàn)(表3)。2021年接種8 d后的病斑面積數(shù)據(jù)顯示,病斑面積的分布范圍在0.3—2.83 cm。病斑面積最小的是‘藍(lán)皮爾曼’,僅為(0.3±0.11)cm。而接種8 d后病斑面積最大的是‘茜’,達(dá)到(2.83±0.47)cm。4—8 d內(nèi)的病斑面積增長(zhǎng)率分析結(jié)果顯示,‘藍(lán)皮爾曼’的病斑面積增長(zhǎng)率最低,僅為 4.75%。而病斑面積增長(zhǎng)率最高的品種是‘茜’,其病斑面積增長(zhǎng)率高達(dá) 56.25%。綜合病斑面積和病斑面積增長(zhǎng)率,可以看出病斑面積和病斑面積增長(zhǎng)率基本保持一致。2021年聚類(lèi)分析結(jié)果顯示(圖1),84個(gè)栽培品種中,‘藍(lán)皮爾曼’‘平成’‘艾達(dá)紅’‘Yellow Transparent’‘Ce1’‘NJ-90’‘無(wú)銹金冠’‘秦冠’‘皇家嘎啦’‘皮諾娃’‘玫瑰光芒’‘夕陽(yáng)’‘Northfield Beauty’‘信濃甜’‘寶羅紅’‘Kiku’‘Gloster69’‘阿萊特’‘蜜脆’‘Challenger’‘凱密歐’‘秦陽(yáng)’‘美味’‘Ce2’‘Su’聚為一類(lèi),說(shuō)明他們是抗性較強(qiáng)的品種;而‘茜’單獨(dú)聚為一類(lèi),是極其不抗的品種;此外,‘Fujiko’‘埃德?tīng)柌┧苟喾颉铩古煽恕甂296’‘昂林’‘早紅霞’‘甘紅’‘王林’‘Redcord’‘富紅早嘎’‘長(zhǎng)富2號(hào)’‘Chanterler’‘新喬納金’‘荷斯坦’‘Ruby Mac’聚為一類(lèi),是低抗性品種。
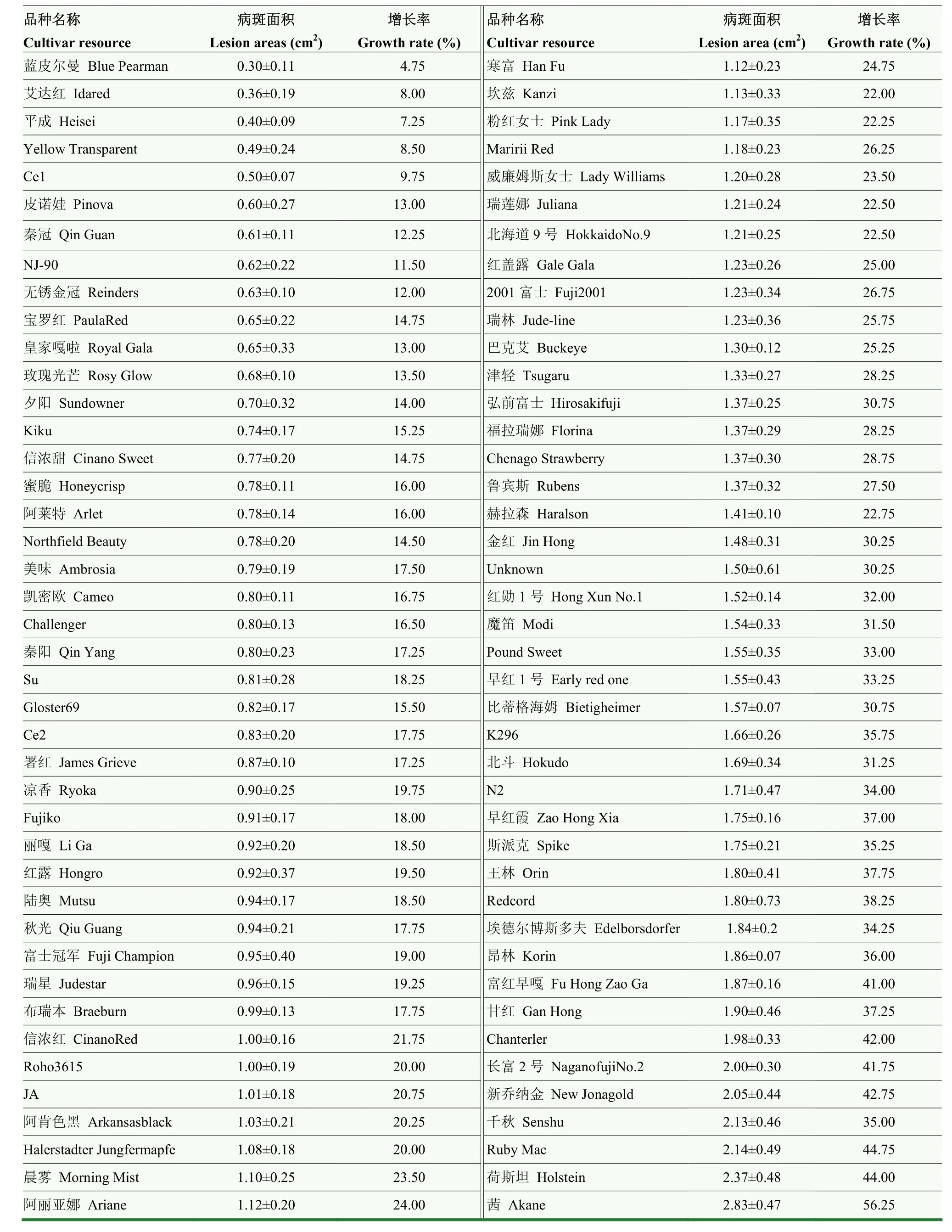
表3 84個(gè)蘋(píng)果屬栽培品種離體葉片接種病斑大小及病斑面積增長(zhǎng)率(2021年)Table 3 Lesion area and its growth rate of 84 apple cultivars inoculated with Alternaria alternata f. sp. mali
綜合兩年的聚類(lèi)分析數(shù)據(jù),‘凱密歐’‘Kiku’‘皇家嘎啦’‘蜜脆’‘藍(lán)皮爾曼’‘艾達(dá)紅’‘Gloster69’在兩年的抗性分析中均為抗性品種;而‘埃德?tīng)柌┧苟喾颉瓹hanterler’均為低抗性品種。
2.3 蘋(píng)果早期斑點(diǎn)落葉病抗病性與 SNP的全基因組關(guān)聯(lián)分析(GWAS)
為了進(jìn)一步探索蘋(píng)果抗早期斑點(diǎn)落葉病的遺傳基礎(chǔ),對(duì) 84份蘋(píng)果栽培品種在接種蘋(píng)果斑點(diǎn)落葉病8 d后的病斑面積數(shù)據(jù)進(jìn)行GWAS分析。因?yàn)?020年8 d接種的病斑面積分布更符合正態(tài)分布的趨勢(shì),因而選其數(shù)據(jù)作為表型進(jìn)行全基因組關(guān)聯(lián)分析。供試群體的病斑面積最小0.3 cm,最大為2.14 cm。病斑面積的峰度和偏度分別為-0.74和0.34,二者的絕對(duì)值都小于1,說(shuō)明蘋(píng)果早期斑點(diǎn)落葉病抗病性具有正態(tài)分布特征,呈現(xiàn)數(shù)量性狀遺傳特征(圖2)。
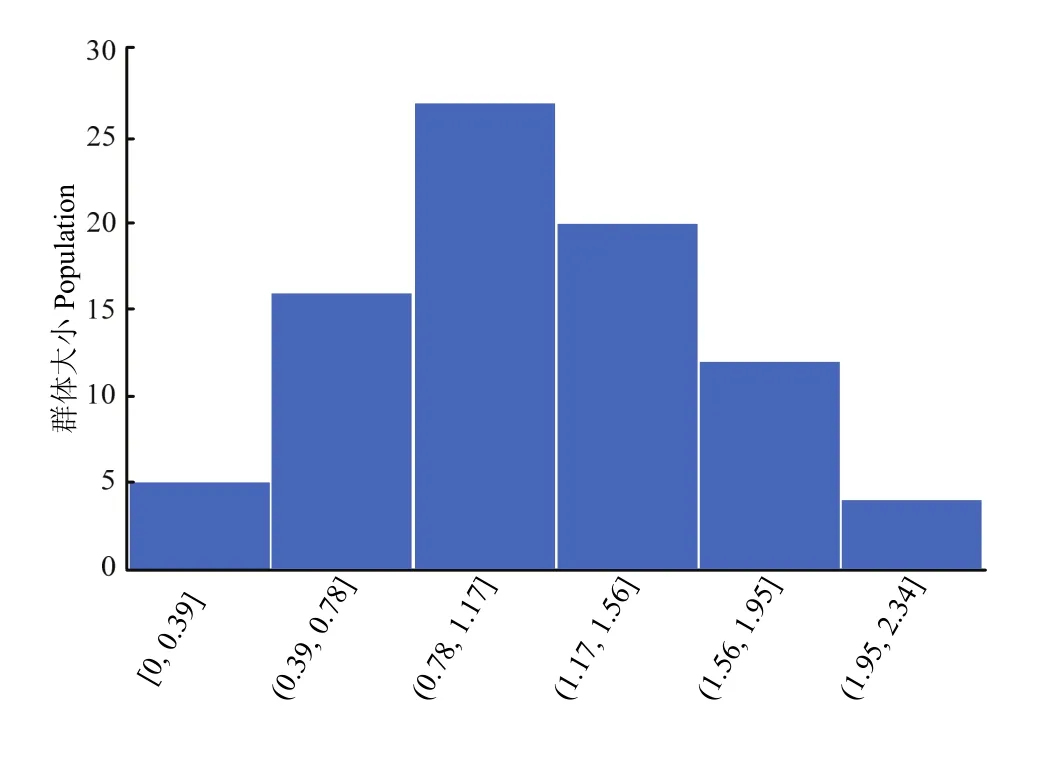
圖2 接種斑點(diǎn)落葉病8 d后的葉片病斑面積表型分布圖Fig. 2 Phenotypic distribution of lesion area after inoculated with Alternaria alternata f. sp. mali for eight days
從 GWAS結(jié)果的分位數(shù)-分位數(shù)圖(quantilequantile plot—Q-Q plot)中看出,Q-Q圖左下角觀察到的值與預(yù)期值一致(圖3)。因此,當(dāng)前統(tǒng)計(jì)模型得到的值符合期望值,證明該統(tǒng)計(jì)模型的合理性。同時(shí),Q-Q圖的右上角為顯著性較高的SNP位點(diǎn),這些候選位點(diǎn)可能與蘋(píng)果抗斑點(diǎn)落葉病相關(guān)聯(lián)。

圖3 斑點(diǎn)落葉病抗性相關(guān)性狀全基因組關(guān)聯(lián)分析曼哈頓圖和分位數(shù)-分位數(shù)圖(QQ圖)Fig. 3 Manhattan plots and Quantile-Quantile Plot (QQ plot) of genome wide association analysis of resistance to Alternaria alternata f. sp. mali in apple cultivars
如圖3所示,本研究共檢測(cè)到6個(gè)與蘋(píng)果早期斑點(diǎn)落葉病抗病性顯著關(guān)聯(lián)的位點(diǎn)(<0.0000001,-Lg≥7)(表4),集中分布在蘋(píng)果的5號(hào)染色體(3個(gè))和0號(hào)染色體(3個(gè))上。其中位于5號(hào)染色體9 336 893 bp位置上的SNP為最顯著的關(guān)聯(lián)位點(diǎn)。該位點(diǎn)位于基因和基因之間的基因間區(qū)。其中和擬南芥的為同源基因,屬于整合素連接蛋白激酶家族中的成員。前人報(bào)道該基因通過(guò)調(diào)控蛋白質(zhì)氨基酸的磷酸化在植物的先天性免疫應(yīng)答過(guò)程中發(fā)揮作用。和擬南芥的同源,編碼FMN連鎖氧化還原酶。此外,其他5個(gè)SNP也關(guān)聯(lián)到7個(gè)候選基因(表4、表5),其中基因被 3個(gè) SNP位點(diǎn)關(guān)聯(lián)到,被2個(gè)SNP位點(diǎn)關(guān)聯(lián)到,表明二者參與蘋(píng)果調(diào)控斑點(diǎn)落葉病抗性的潛在可能性。其他5個(gè)候選基因分別為核糖體結(jié)合因子A家族蛋白、B-box型鋅指家族蛋白、多蛋白/逆轉(zhuǎn)錄轉(zhuǎn)座子、植物特異性GATA型轉(zhuǎn)錄因子的一個(gè)新家族成員、堿性亮氨酸拉鏈(bZIP)轉(zhuǎn)錄因子家族蛋白。
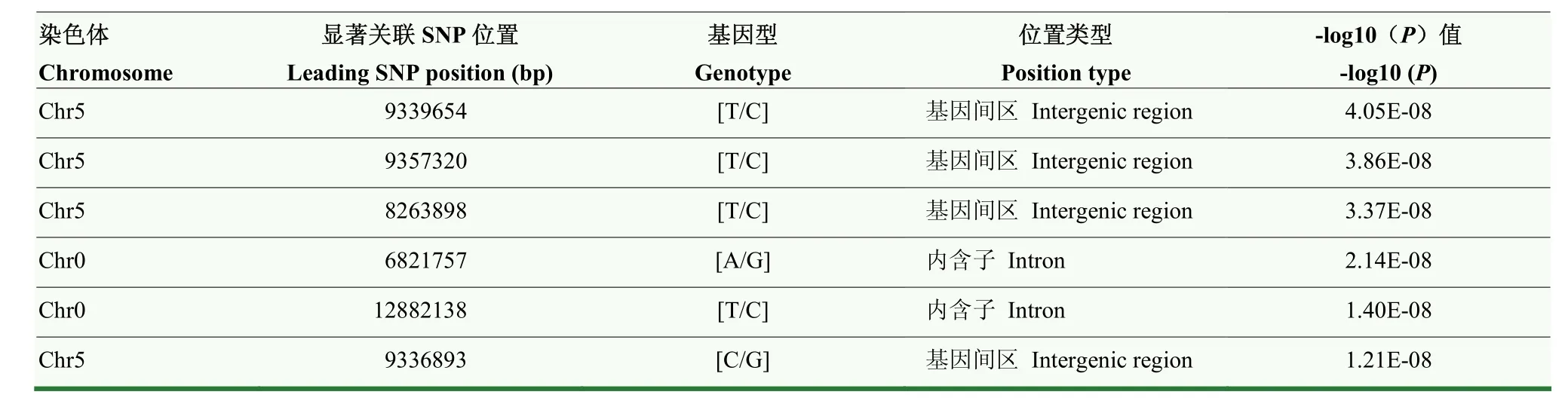
表4 與抗病性狀相關(guān)的顯著關(guān)聯(lián)SNP位點(diǎn)Table 4 SNPs identified to be associated with Alternaria alternata f. sp. mali resistance (P<0.0000001)
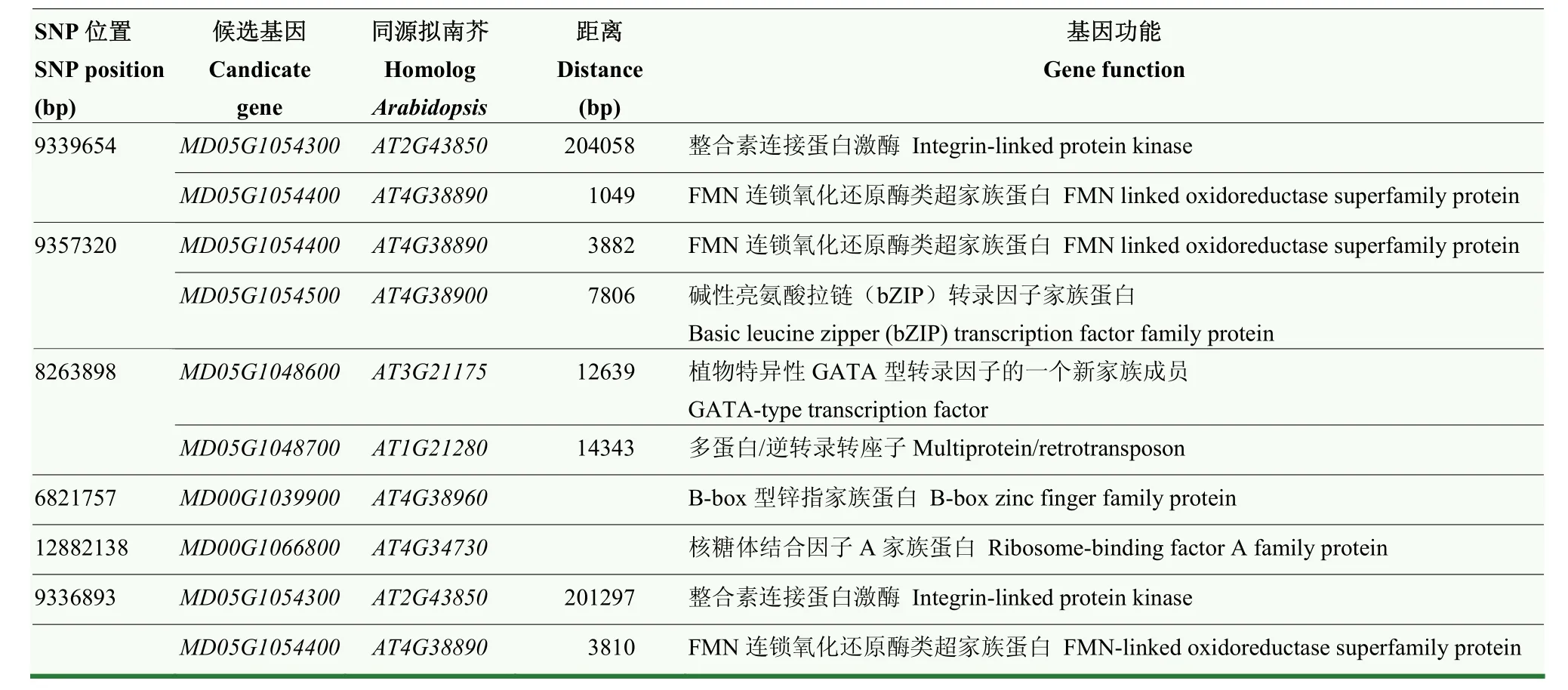
表5 蘋(píng)果抗斑點(diǎn)落葉病的候選基因功能注釋Table 5 Functional annotation of candidate genes
2.4 候選基因抗斑點(diǎn)落葉病的生物學(xué)功能驗(yàn)證
為了進(jìn)一步確認(rèn)7個(gè)關(guān)鍵候選基因、、、、、、在抗斑點(diǎn)落葉病中發(fā)揮的生物學(xué)功能,首先對(duì) 7個(gè)關(guān)鍵候選基因在接種斑點(diǎn)落葉病菌 0、12、24和48 h后的基因表達(dá)水平進(jìn)行檢測(cè),試驗(yàn)結(jié)果顯示7個(gè)關(guān)鍵候選基因在接種12和24 h后,基因的表達(dá)量均顯著提高(圖4),預(yù)示7個(gè)關(guān)鍵候選基因均在抗病功能當(dāng)中發(fā)揮著重要作用。其中,在接種12和24 h后的基因上調(diào)水平達(dá)到 7倍左右,相比于其他 6個(gè)候選基因,表達(dá)水平響應(yīng)最為強(qiáng)烈。于是將構(gòu)建到超表達(dá)載體上,對(duì) GL-3(‘嘎啦’蘋(píng)果實(shí)生后代)組培苗蘋(píng)果葉片進(jìn)行瞬時(shí)轉(zhuǎn)化。隨后對(duì)瞬時(shí)超表達(dá)的蘋(píng)果葉片在第4天和第7天進(jìn)行表達(dá)水平的檢測(cè),發(fā)現(xiàn)在第7天時(shí)仍能過(guò)表達(dá)2倍。同時(shí)對(duì)瞬時(shí)超表達(dá)的蘋(píng)果葉片在第4天進(jìn)行斑點(diǎn)落葉病接種試驗(yàn),結(jié)果顯示,接種 3 d后在超表達(dá)的葉片中的病斑面積明顯小于非轉(zhuǎn)基因葉片對(duì)照中的(圖5)。
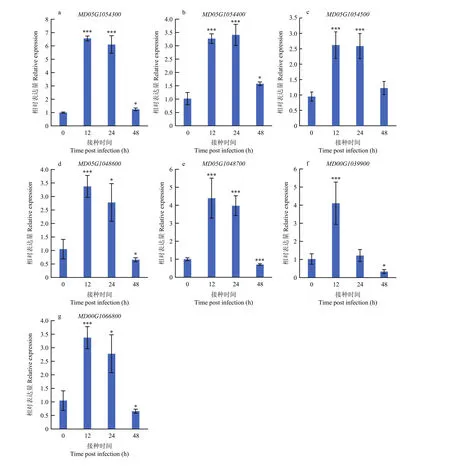
圖4 關(guān)鍵候選基因在接種斑點(diǎn)落葉病后的表達(dá)水平分析Fig. 4 Expression level of key candidate genes after inoculation with Alternaria alternata f. sp. mali
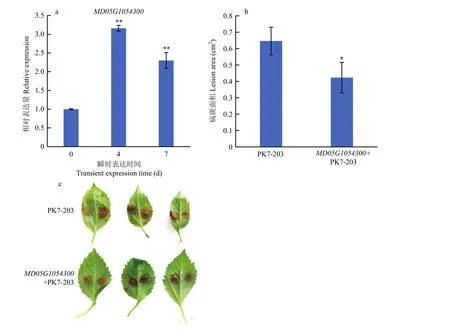
圖5 超表達(dá) MD05G1054300 的葉片抗病性Fig. 5 Disease symptoms of GL-3 leaves transiently expressing MD05G1054300
3 討論
3.1 蘋(píng)果栽培品種對(duì)斑點(diǎn)落葉病的抗性評(píng)價(jià)
蘋(píng)果早期落葉病嚴(yán)重威脅蘋(píng)果園的生產(chǎn)。早期落葉病爆發(fā)時(shí),一片葉子上就能產(chǎn)生數(shù)十至上百個(gè)病斑,嚴(yán)重時(shí)病斑甚至連成一片,造成果園早期大面積落葉;同時(shí)感染病菌的果實(shí)極易因受二次寄生而發(fā)生腐爛現(xiàn)象,最終造成蘋(píng)果園產(chǎn)量和果實(shí)品質(zhì)的急劇下滑,因此,有效防治蘋(píng)果早期落葉病一直是國(guó)內(nèi)外特別關(guān)注的領(lǐng)域。研究學(xué)者通過(guò)探究斑點(diǎn)落葉病的發(fā)生規(guī)律,進(jìn)行抗早期落葉病的基因定位分析,尋找有效的抗病基因,進(jìn)行分子育種,以期達(dá)到蘋(píng)果樹(shù)高抗病性的目的。近年來(lái),隨著離體葉片的室內(nèi)接種評(píng)價(jià)體系的完善,這種評(píng)價(jià)方式被廣泛應(yīng)用于蘋(píng)果斑點(diǎn)落葉病、褐斑病等葉片病害的抗性評(píng)價(jià)研究。這種離體方法可以避免由外界環(huán)境因素的多樣性和多變性對(duì)試驗(yàn)結(jié)果造成的干擾,也可以避免由于病原菌的數(shù)量、活力不同而造成的差異影響,評(píng)價(jià)結(jié)果具有穩(wěn)定可重復(fù)性的優(yōu)勢(shì)。本研究對(duì)84個(gè)蘋(píng)果屬栽培品種進(jìn)行斑點(diǎn)落葉病的抗性評(píng)價(jià),從中鑒定出7個(gè)穩(wěn)定抗性品種,如‘凱密歐’‘Kiku’‘皇家嘎啦’等。說(shuō)明蘋(píng)果屬栽培品種中包含豐富的抗性資源,可為生產(chǎn)中引種、培育抗性品種和果園中品種的合理布局提供指導(dǎo)。中國(guó)的很多自育品種具有優(yōu)良的性狀特征,如:‘秦冠’具有豐產(chǎn)和抗逆性強(qiáng)的特點(diǎn);‘秦陽(yáng)’具有早熟和易結(jié)果的特性;‘寒富’具有果實(shí)品質(zhì)優(yōu)良,抗寒性極強(qiáng)的特征。本研究中,‘秦冠’‘秦陽(yáng)’和‘寒富’均表現(xiàn)出對(duì)斑點(diǎn)落葉病的中等抗性。其中‘秦冠’的抗斑點(diǎn)落葉病能力最強(qiáng),但相較于本研究中篩選出的7份抗性品種,‘秦冠’的抗病性還略有欠缺。目前,中國(guó)大面積栽培‘富士’蘋(píng)果,但該品種的斑點(diǎn)落葉病抗性極差,因此,亟待進(jìn)行品種改良以促進(jìn)蘋(píng)果產(chǎn)業(yè)健康均衡發(fā)展。本研究鑒定到7份抗病性的蘋(píng)果品種,這些栽培種在果實(shí)品質(zhì)和果樹(shù)長(zhǎng)勢(shì)等方面均有相關(guān)報(bào)道,如路新創(chuàng)等研究發(fā)現(xiàn)美洲蘋(píng)果‘蜜脆’對(duì)斑點(diǎn)落葉病的抗病能力強(qiáng),這與本研究的結(jié)果一致。
3.2 蘋(píng)果斑點(diǎn)落葉病抗性研究機(jī)制
通過(guò)比較蘋(píng)果栽培品種對(duì)斑點(diǎn)落葉病的抗性,篩選高抗品種,進(jìn)而通過(guò)品種改良使蘋(píng)果園免受斑點(diǎn)病侵害。但是由于傳統(tǒng)的蘋(píng)果育種周期長(zhǎng),且雜交育種的不定性較強(qiáng),近年來(lái)通過(guò)分子育種手段選育高抗病性蘋(píng)果種質(zhì)的研究越來(lái)越多。本研究中,通過(guò)GWAS關(guān)聯(lián)到顯著性候選基因中(附表),多次關(guān)聯(lián)到參與細(xì)胞凋亡及防衛(wèi)反應(yīng)的含有NB-ARC結(jié)構(gòu)域的抗病相關(guān)蛋白基因(NB-ARC domain-containing disease resistance protein)和,其在病害脅迫中發(fā)揮著重要作用。目前關(guān)于蘋(píng)果抗斑點(diǎn)落葉病的抗菌機(jī)制主要集中在抗菌蛋白與病原菌幾丁質(zhì)的結(jié)合、誘導(dǎo)植物木質(zhì)素的沉積、激素響應(yīng)、增強(qiáng)植物氧化還原能力等方面。蘋(píng)果由真菌幾丁質(zhì)誘導(dǎo),其蛋白可以與幾丁質(zhì)結(jié)合,還能通過(guò)加強(qiáng)HO積累和胼胝質(zhì)沉積增加蘋(píng)果對(duì)病原真菌的防御反應(yīng)。有研究發(fā)現(xiàn)能夠增厚細(xì)胞壁,誘導(dǎo)茉莉酸響應(yīng)途徑,增強(qiáng)抗氧化相關(guān)酶活性從而提高植物的抗病性。ZHANG等研究發(fā)現(xiàn)兩種含有卷曲螺旋、核苷酸結(jié)合和富含亮氨酸重復(fù)(CCR-NB-LRR)結(jié)構(gòu)域的抗性(R)蛋白MdRNL2和 MdRNL6相互作用形成一種 MdRNL2-MdRNL6復(fù)合物,與新型發(fā)病機(jī)制相關(guān)(PR)蛋白MdPR10-1相互作用以抑制真菌生長(zhǎng);在其 2018年的研究中發(fā)現(xiàn)‘金冠’()和‘寒富’()的啟動(dòng)子區(qū)域中鑒定出單核苷酸多態(tài)性(SNP),即-1 186 bp的motif b(G-T)。當(dāng)突變感病品種中啟動(dòng)子 motif b的G為T(mén)時(shí),啟動(dòng)子活性喪失,表明中的 SNP與斑點(diǎn)落葉病抗性相關(guān)。因此,該 SNP可作為區(qū)分對(duì)斑點(diǎn)落葉病具有抗性或易感性的蘋(píng)果品種的標(biāo)記。ASR(ABA/water stress/ripening-induced)基因家族在植物應(yīng)對(duì)斑點(diǎn)落葉病過(guò)程中存在不同的基因表達(dá)模式,暗示 Md-ASR基因家族在病害脅迫中發(fā)揮著重要作用。有研究者通過(guò)對(duì)新疆野蘋(píng)果的抗斑點(diǎn)落葉病轉(zhuǎn)錄組研究發(fā)現(xiàn),其響應(yīng)斑點(diǎn)落葉病的主要方式為茉莉酸信號(hào)途徑、乙烯信號(hào)途徑和水楊酸信號(hào)途徑。此外,該研究還發(fā)現(xiàn)為蘋(píng)果抗斑點(diǎn)落葉病的主基因之一,該基因能夠增強(qiáng)蘋(píng)果氧化還原酶的能力,降低活性氧含量,從而使蘋(píng)果更抗斑點(diǎn)落葉病。上述研究結(jié)果表明,植物的抗病機(jī)制和氧化還原能力相關(guān),而本研究定位到的關(guān)鍵候選基因(FMN,連鎖氧化還原酶類(lèi)超家族蛋白),參與植物以氧化還原為特征的各類(lèi)生理反應(yīng)。
目前,GWAS分析是一種有效的復(fù)雜性狀功能定位的正向遺傳學(xué)分析策略,可利用群體內(nèi)所有個(gè)體全基因組水平上的等位遺傳變異和表型變異進(jìn)行相關(guān)性分析,從而鑒定出與目標(biāo)性狀顯著連鎖的等位變異位點(diǎn),最后分析等位基因型對(duì)表型的遺傳效應(yīng),在植物數(shù)量性狀遺傳結(jié)構(gòu)解析中取得了豐碩成果。GWAS分析現(xiàn)已被廣泛用于農(nóng)作物定位,如小麥籽粒品質(zhì)性狀和小麥抗倒伏性狀的關(guān)聯(lián)位點(diǎn)和候選基因的預(yù)測(cè)。但是由于參考基因組雜合度高,性狀統(tǒng)計(jì)較難等原因,GWAS分析在蘋(píng)果中的研究還很少。本研究以前期獲得的高質(zhì)量 SNP數(shù)據(jù)為基礎(chǔ),結(jié)合84份蘋(píng)果栽培品種在接種斑點(diǎn)落葉病8 d后的表型數(shù)據(jù)進(jìn)行GWAS分析,最終得到6個(gè)顯著SNP位點(diǎn),并隨后關(guān)聯(lián)到7個(gè)關(guān)鍵候選基因。其中(FMN,連鎖氧化還原酶類(lèi)超家族蛋白)被關(guān)聯(lián)到3次,(ILK1,整合素連接蛋白激酶)被關(guān)聯(lián)到2次,且這兩個(gè)基因在蘋(píng)果‘金冠’基因組的5號(hào)染色體上緊密相連。前人研究證明核黃素可以直接參與植物以氧化還原為特征的各類(lèi)生理反應(yīng),從而誘導(dǎo)植物對(duì)多種病害的防治,而FMN是核黃素在體內(nèi)具有生物活性的主要形式,說(shuō)明蘋(píng)果FMN在參與抗斑點(diǎn)落葉病的潛在可能性。此外,在擬南芥中整合素連接蛋白激酶(ILK1)已被報(bào)道有助于提高植物對(duì)病原體的防御,本研究在蘋(píng)果中超表達(dá)能提高蘋(píng)果斑點(diǎn)落葉病抗性,表明其具有和類(lèi)似的功能。本研究預(yù)測(cè)的關(guān)鍵候選基因中有 3個(gè)轉(zhuǎn)錄因子,包括堿性亮氨酸拉鏈(bZIP)轉(zhuǎn)錄因子()、植物特異性GATA型轉(zhuǎn)錄因子()和B-box(BBX)家族轉(zhuǎn)錄因子()。這三者均被證明對(duì)植物的抗逆和生長(zhǎng)發(fā)育起關(guān)鍵調(diào)控作用。如大米的屬于bZIP家族蛋白,具有能夠直接結(jié)合下游抗病基因啟動(dòng)子的能力,進(jìn)而調(diào)控抗病相關(guān)基因的表達(dá),也有研究報(bào)道辣椒中過(guò)表達(dá)能夠增強(qiáng)植物的抗病性;小麥中過(guò)表達(dá)GATA型轉(zhuǎn)錄因子—TaGATA1則能夠顯著增強(qiáng)小麥對(duì)紋枯病的抗性;甘薯的B-box轉(zhuǎn)錄因子—IbBBX24可以通過(guò)調(diào)控茉莉酸(JA)通路來(lái)響應(yīng)枯萎病的侵害。盡管本研究中鑒定到的候選基因(、和)在蘋(píng)果的抗病研究中還未見(jiàn)報(bào)道,但是根據(jù)同源基因的功能預(yù)測(cè),推測(cè)這些基因可能與蘋(píng)果抗病性緊密相關(guān)。后續(xù)研究可將本文鑒定到的這些候選基因作為重點(diǎn),深入探究其在蘋(píng)果抗斑點(diǎn)落葉病中的生物學(xué)功能及分子機(jī)制。
4 結(jié)論
在 84份蘋(píng)果屬栽培品種中鑒定到斑點(diǎn)落葉病的抗性品種7份、易感品種2份。此外,鑒定到6個(gè)與蘋(píng)果斑點(diǎn)落葉病抗病性顯著關(guān)聯(lián)的SNP位點(diǎn),并關(guān)聯(lián)到7個(gè)關(guān)鍵候選基因,其中位于5號(hào)染色體9 336 893 bp位置上的SNP為最顯著關(guān)聯(lián)位點(diǎn),且這個(gè)位點(diǎn)上下游的兩個(gè)基因 FMN連鎖氧化還原酶類(lèi)超家族蛋白()和整合素連接蛋白激酶()為關(guān)鍵的候選基因,其中超表達(dá)可顯著提高蘋(píng)果斑點(diǎn)落葉病抗性。
猜你喜歡
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:30
當(dāng)代陜西(2021年17期)2021-11-06 03:21:36
原道(2020年2期)2020-12-21 05:47:06
當(dāng)代陜西(2019年15期)2019-09-02 01:52:00
中國(guó)非營(yíng)利評(píng)論(2018年2期)2018-06-18 10:48:50
學(xué)苑創(chuàng)造·A版(2018年11期)2018-02-01 06:29:20
自動(dòng)化學(xué)報(bào)(2017年1期)2017-03-11 17:31:17
讀者(2017年5期)2017-02-15 18:04:18
西藏科技(2016年5期)2016-09-26 12:16:39
振動(dòng)工程學(xué)報(bào)(2015年1期)2015-03-01 01:15:42
