β-折疊抗菌肽的研發及應用策略
2022-08-26 08:49:14邵長軒黨安凱戰昭含來振衡朱永杰單安山
畜牧獸醫學報 2022年8期
邵長軒,黨安凱,戰昭含,來振衡,朱永杰,單安山
(東北農業大學動物營養研究所,哈爾濱150030)
近幾十年來,抗生素在畜牧業中的不合理使用已導致細菌耐藥性產生和畜產品抗生素殘留問題,這些耐藥菌不僅造成了難以治療的畜禽疾病,還能通過動物相關食品傳播到人類,嚴重威脅人類健康,當下新型抗生素的研發速度相比于耐藥性細菌產生和變異的速度已趨于緩慢。早在2006年,歐盟就實行了“飼料全面禁抗”,而在2019年7月10日,中華人民共和國農業農村部發布了第194號公告,公布了“全面禁止促生長類藥物飼料添加劑”的決議,因此,尋找綠色、安全、高效的抗生素替代品已經成為當前迫切需要解決的問題。抗菌肽(antimicrobial peptides,AMPs)是宿主先天性免疫防御系統產生的一類抵抗外界病原體感染的小分子多肽,具有廣譜抗菌、抗病毒、抗癌活性以及免疫調節功能,還可促進上皮傷口愈合,用于人類疾病的治療。根據當前抗菌肽數據庫(Antimicrobial Peptide Database,APD)的最新結果顯示,目前,已經發現了超過3 000種具有多樣序列和結構的天然AMPs,在微生物、植物、無脊椎動物、以及更復雜的兩棲動物和哺乳動物等多物種中廣泛存在,其主要的物理破膜殺菌機制使病原體不易對其產生耐藥性突變,因此有望成為新一代抗生素替代物。
抗菌肽的分類依據多樣,例如:根據物種來源、合成機制或分子靶標等,但較常見的是基于結構的差異將它們進行劃分。如圖1所示,根據二級結構的類型,抗菌肽通常被分為4大類,分別是α-螺旋肽;不含α-螺旋和β-折疊成分但通常富含特定氨基酸(如Gly、Arg、Trp或Pro)的線性延伸結構;由一個或多個二硫鍵穩定的β-折疊肽;同時包含α-螺旋和β-折疊結構的肽。α-螺旋目前是AMPs中研究的最為深入和廣泛的一類二級結構,但已有研究表明某些α-螺旋肽提取工藝復雜、細胞毒性高以及抗菌效力參差不齊,限制了它們的應用。而β-折疊肽依靠保守的半胱氨酸殘基之間的分子內二硫鍵維持三維結構,在相同條件下這類肽具有更加穩定的二級構象甚至更強的抗酶解能力。有研究表明β-折疊肽與具有相同電荷和疏水性的α-螺旋肽相比,具有相近的抗菌活性和更優異的細胞選擇性,而且,當穩定β-發卡或β-折疊結構抗菌肽中的二硫鍵被破壞時,所形成的線狀多肽仍能保持其抗菌特性。由于β-折疊AMPs所含的復雜二硫鍵合成困難且成本昂貴,其研究并不廣泛,因此β-折疊抗菌肽的研發對于未來AMPs的應用轉化十分關鍵。
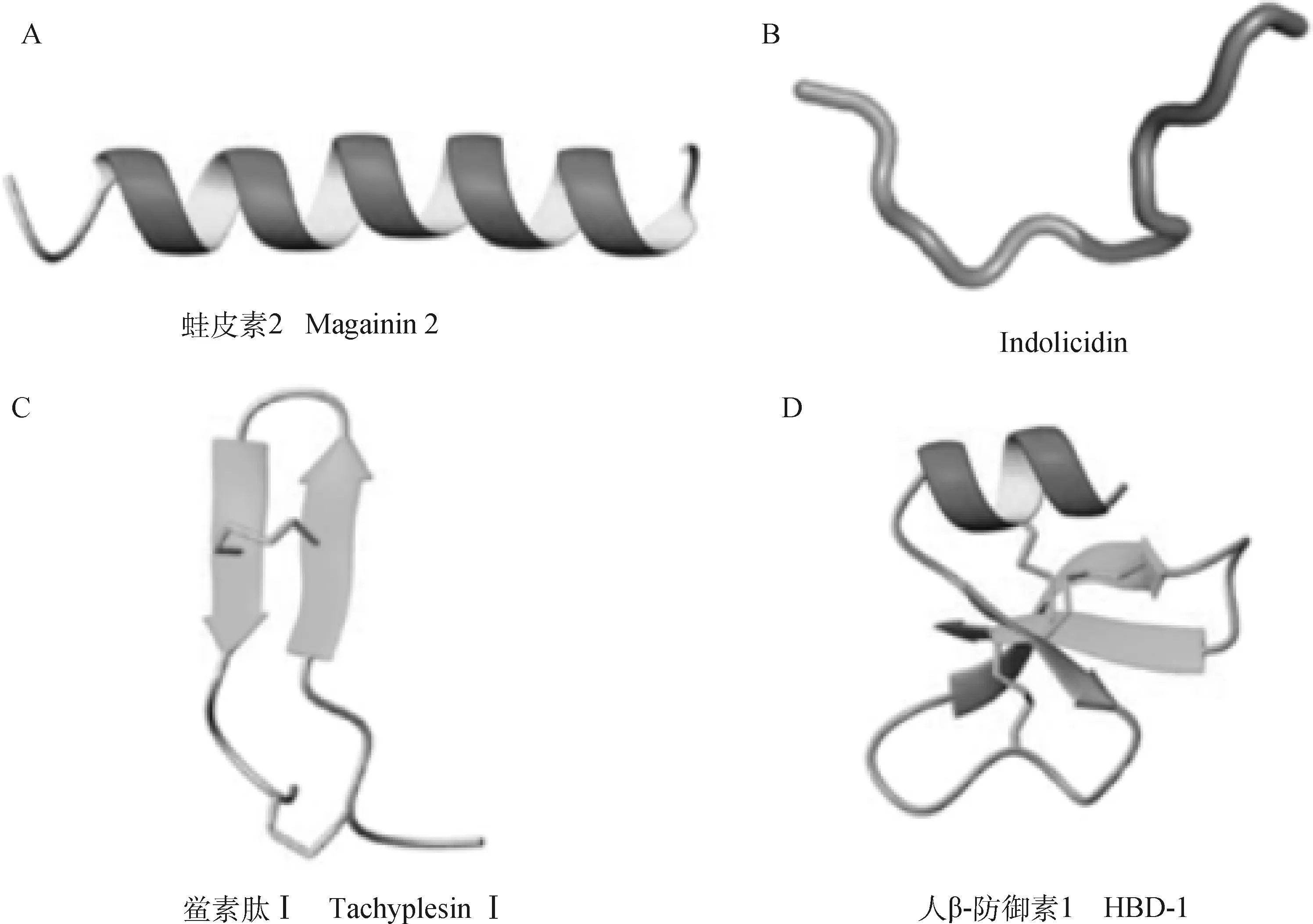
A. α-螺旋結構;B.線性延伸結構;C. β-折疊結構;D. 同時包含α-螺旋和β-折疊元素A. α-helical structure;B. Linear extension structure;C. β-sheet structure;D. Contain both α-helical and β-sheet elements圖1 天然抗菌肽的常見結構類型[13-14]Fig.1 Common structural classes of naturally occurring AMPs[13-14]
本文介紹了β-折疊抗菌肽的天然來源、分子設計及主要抑菌機理,同時對其應用的局限性以及相應的策略進行了綜述,全方位描述了β-折疊抗菌肽的研究進展,為該類肽的研發與利用提供參考。
1 天然源β-折疊抗菌肽
抗菌肽來源廣泛,作為非特異性天然防御系統的重要組成部分,是機體抵御病原微生物入侵的第一道防線。這些廣泛來源的抗菌肽通常有著不同的結構,β-折疊就被包括其中。根據來源,可以將β-折疊抗菌肽分為哺乳動物源、昆蟲源、植物源以及微生物源等大類,在表1中列舉了較為常見的幾種天然β-折疊抗菌肽及其主要特征。
基于半胱氨酸的含量和結構特征,可將β-折疊抗菌肽分為兩類,分別是β-發卡抗菌肽和α-defensins肽。β-發卡是反平行β-折疊結構的最簡單形式,由兩個β-鏈和單個二硫鍵組成的環狀區域定義。β-發卡抗菌肽尺寸小且生物活性廣泛,在實際應用方面受到研究人員更多關注,但天然β-發卡抗菌肽并不豐富,已知的β-發卡型抗菌肽家族不僅在結構和生物學功能上有顯著差異,在生物合成途徑上也有顯著差異。Defensins肽跨越多個結構類別,α-defensins是一個亞組,包括三條反向平行的β-側鏈,側鏈以“trans”排列與二硫鍵相連(即β鏈末端的兩個二硫鍵指向相反的方向,并連接兩個不同的側鏈)。α-defensins被認為是通過基因復制而進化的,存在于大多數哺乳動物中,具有廣譜抗細菌、真菌和病毒的活性。這些肽含有保守的鹽橋結構,有研究表明,鹽橋對于多肽的正常折疊和蛋白水解抗性是必需的,但對抗菌活性而言不是必需的。

表1 不同來源的幾種常見β-折疊抗菌肽
1.1 哺乳動物源
哺乳動物源抗菌肽已經廣泛在動物皮膚、腸道、免疫系統、神經系統和泌尿系統等多種生理系統和上皮組織表面分離鑒定,主要包括Cathelicidins、Defensins和Histatins 3大家族,研究最廣泛的為Defensins類抗菌肽。APD是一個集抗菌肽命名、分類、信息搜索、預測、設計和統計等多功能于一體的抗菌肽數據分析系統,截至目前APD數據庫共收錄有26種被明確鑒定為β-折疊結構的哺乳動物源AMPs,其中牛乳鐵蛋白Lfcin B和豬源Protegrin 1的研究較為深入。
1.2 昆蟲源
昆蟲免疫防御僅依賴于先天免疫,當微生物突破身體的第一道防線時,昆蟲的先天免疫反應就會被激活,快速產生大量的AMPs,使昆蟲迅速消滅入侵病原體。昆蟲源β-折疊抗菌肽具有分子量小、熱穩定性強、水溶性好、無免疫原性等特點,不僅對細菌和真菌有廣譜抗菌活性,對病毒、原蟲及癌細胞也有殺傷作用。目前,研究較為深入的昆蟲源β-折疊抗菌肽是擁有廣譜抗菌活性的Thanatin,此外還有從長臂天牛體內分離出的擁有抗真菌功能的Alo-3。昆蟲作為世界上最大的生物種群,種類繁多,分布廣泛,因此昆蟲源β-折疊抗菌肽還具有巨大的發掘潛力。
1.3 植物源
植物中存在多種具有藥理作用的分子蛋白,β-折疊抗菌肽就是其中之一。當各種病原體入侵時,多種β-折疊抗菌肽(如從紫花苜蓿中分離出的NCR169)能夠通過干擾細胞壁或膜脂的生物合成發揮抗菌活性。此外,植物源β-折疊抗菌肽富含保守的半胱氨酸殘基,擁有極高的穩定性且能通過改變非半胱氨酸殘基的序列從而識別不同的靶點,預示著它們作為抗菌劑的巨大發展潛力。
1.4 微生物源
微生物源β-折疊抗菌肽是由微生物自身分泌的用來保護自己并殺滅或抑制其他細菌、真菌的小分子多肽,其主要來源于特定細菌,通常比抗生素穩定,可以承受高溫和極端pH,但在自然和有機環境中的生物半衰期較短,主要通過作用于靶細胞膜來發揮作用。迄今APD共收錄了15種來自細菌的微生物源β-折疊抗菌肽,在食品保鮮和微生物感染治療等領域有著廣泛的應用前景。
2 β-折疊抗菌肽抗細菌機理
大部分β-折疊抗菌肽可以直接作用于細菌細胞膜,但有一些抗菌肽除了引起細胞膜損傷之外,還被證實可以通過細胞膜轉運并結合到細胞內的目標來殺死細菌,產生一系列復雜的胞內活動。如圖2所示,根據作用位點的不同,一般β-折疊抗菌肽的抑菌機理可以分為兩類,分別是肽與細菌細胞膜相互作用以及肽與胞內靶點相互作用。

圖2 抗菌肽的抗細菌作用機理[38]Fig.2 Action mechanism of AMPs against bacteria[38]
2.1 肽與細菌細胞膜相互作用
β-折疊抗菌肽與細胞膜的作用過程,首先陽離子殘基與靶細胞膜陰離子表面通過靜電相互作用彼此靠近,然后肽的兩親結構域與磷脂雙分子層之間產生疏水相互作用,抗菌肽插入細胞膜導致細胞內容物滲漏和死亡。目前,最為經典的抗菌肽膜作用模型有桶壁模型、地毯模型和環形孔模型,這幾種作用機制模型并不是相互獨立的,而在細菌細胞膜被破壞過程中可能同時存在。由于β-折疊抗菌肽主要通過與細菌細胞膜相互作用,過程中不涉及特定的靶標,這也解釋了細菌難以產生耐藥性的原因。Bellamy等研究發現,Lactoferricin B能夠與細胞膜上帶負電荷的磷脂和脂多糖結合并增加膜的通透性,且伴隨著溶液中鹽離子所帶正電荷的增加抑菌效果顯著降低;Sinha團隊通過將Thanatin與DPC-d膠束相互結合闡述了Thanatin導致質膜結構擾動的機制;Mani等使用F和H自旋擴散核磁共振技術確定了Protegrin 1(PG-1)在陰離子膜和含膽固醇中性膜中的低聚物結構,驗證了PG-1能誘導細菌細胞膜快速通透從而產生殺菌活性。
2.2 肽與胞內靶點相互作用
起初人們認為β-折疊抗菌肽的跨膜機制是其作用的唯一模式,但是越來越多的研究提供證據表明僅出現膜通透性不足以導致細胞死亡,并由此發現了部分β-折疊抗菌肽存在細胞內靶點。例如通過抑制核酸的生物合成和代謝或抑制蛋白合成與折疊,進而抑制和殺死病原菌,此外還有些能抑制細胞分裂和細胞壁的生物合成。Leeuw等研究發現α-防御素類多肽HNP1可以通過與細胞壁前體脂質II相互作用抑制肽聚糖的合成,從而影響細胞壁合成以及膜功能導致細菌死亡;在海洋類動物的體腔細胞中分離出來的β-折疊類抗菌肽Arenicin-1可以誘導細胞內ROS的增加發揮抗真菌活性,介導細胞的凋亡過程;β-發卡肽Polyphemusin I通過作用于細胞內與DNA和RNA相關的蛋白質靶標從而造成細胞死亡。部分β-折疊多肽還可以與細胞穿透肽結合從而進入細菌細胞內,兩種作用模式之間會表現出協同關系使細菌對于此類殺傷機制更難產生突變抗性。
3 β-折疊抗菌肽的分子設計
3.1 天然源β-折疊抗菌肽的改造
天然β-折疊抗菌肽由于具有二硫鍵維持三維結構,穩定性相對會更高,但是一些二硫鍵較少的天然β-發卡抗菌肽仍然會受到高濃度酶的快速降解,從而失去抑菌活性。有部分天然β-折疊抗菌肽擁有足夠的蛋白酶抗性,但缺少強效的抗菌活性,所以當面臨這一系列的矛盾時,天然源抗菌肽的改造是一種有效手段。研發出具有良好抗菌活性和細胞選擇性的β-折疊抗菌肽,需要利用現有的生物工程技術對這些AMPs的抗菌活性、生物安全性和結構關系進行系統探究解析,近年來,各種優化策略和分子設計技術的進步為肽性能的合理改良和優化提供了參考。目前,該方向的研究已經取得較大進展,常見的改造方法包括氨基酸替換、肽鏈修飾、肽鏈雜合等。
β-折疊抗菌肽的功能及活性往往是由結構所決定的,研究發現影響這些因素的主要結構參數包括凈電荷數、疏水性、兩親性、序列長度、氨基酸組成等。總體帶正電荷使β-折疊抗菌肽能與微生物膜的負電荷成分產生靜電相互作用,疏水基團可以幫助其更深入地滲透細菌膜,從而決定β-折疊抗菌肽的抗菌效力,兩親性構象也是肽具有膜活性不可或缺的,然而研究表明這些參數體現在構效關系上時會有各自的閾值,它們之間縱橫交錯、相互影響。因此在改造中不能盲目追求單個參數的最佳完美,而是要綜合考慮各個參數的影響,以期達到一個最優的平衡點,使肽的抗菌活性和生物安全性趨向于更加合理。
3.1.1 氨基酸替代 在自然界中,許多生物體能產生天然β-折疊抗菌肽,利用這些肽作為起始模板對序列進行修改,可以達到提高肽的抗菌活性和細胞選擇性的目的。氨基酸替代主要是對肽模板添加、刪除、替換一個或幾個氨基酸殘基。Benfield等設計了一種新型肽,首先對Gomesin(GM)骨架進行環化得到一種GM類似物cGM,當濃度高于2 μmol·L時cGM會產生毒性并破壞癌細胞的細胞膜,后來用D-Arg取代了原本在它第4、10和18位的Arg殘基從而合成了[R/r]cGM,在保留了殺菌功能的同時蛋白酶抗性明顯提高,并且解決了毒性過大的問題,可作為靶向和抗癌藥物的輸送系統而不損害健康細胞。另外,Dong等將Protegrin 1(PG-1)的β轉角序列添加上幾個簡單的氨基酸重復序列,設計了一系列末端對稱肽(XR)H(RX)(=1、2;X代表I、F、W和P;H代表CRRRFC),并利用治療指數(therapeutic index,TI),即最小溶血濃度MHC與最小抑菌濃度MIC幾何平均值的比值,評估多肽的細胞選擇性,其中,序列為(IR)H(RI)的β-折疊結構IR2具有最高的TI,顯示出比PG-1更高的抗微生物效力,溶血活性和細胞毒性顯著低于PG-1。在改造過程中需要注意的是,β-折疊抗菌肽是一種結構復雜的分子,合理地去除非必須性質的部分肽區域可以獲得較短的活性基序,降低大規模生產成本和衍生物的毒性或免疫原性,但此方法需要控制在合理范圍,因為氨基酸組成的輕微改變可能會改變整個幾何構型甚至物理化學性質,進而會影響這些肽殺滅微生物的能力和對宿主細胞的毒性。
3.1.2 肽鏈修飾 β-折疊抗菌肽常見的肽鏈修飾方法主要有環化、聚乙二醇修飾、膽固醇修飾、末端區域修飾(乙酰化或酰胺化)等,這些肽鏈修飾策略的可行性已被廣泛證實,不僅能夠用于提高肽的活性、穩定性和溶解性,提供更長的半衰期,也可以調節分子內相互作用和疏水性甚至改變多肽呈現的二級結構。Lei等將人α-defensin 5(HD5)的末端進行了十四烷酸(肉豆蔻酸)修飾,結果發現合成的肉豆蔻酰化HD5在體外具有顯著增強的廣譜殺菌活性,并且穩定性明顯提高;Imura等將聚乙二醇連接到β-折疊肽Tachyplesin I的N端,發現多肽聚乙二醇化后膜通透性并沒有改變,而且抗菌活性顯著減弱,但細胞毒性顯著降低;Chan等采用固相化學合成法合成了環狀β-折疊抗菌肽Gomesin,與天然Gomesin相比,環化提高了Gomesin的體外穩定性,增強了其對癌細胞的殺傷作用以及抗瘧疾活性。相比于其他改造方法,肽鏈修飾策略能夠賦予β-折疊抗菌肽更多本身所不具備的性能,甚至定向賦予某項特性,更加可控與便捷,是一種性價比很高的肽設計方法。
3.1.3 肽鏈雜合 肽鏈雜合是將不同優勢肽或優勢肽段通過某種方式組合成新的嵌合肽的一類方法,這種方法通常將有效但毒性相對較大的肽序列與活性較低但毒性較低的肽序列融合,同樣是設計新型β-折疊肽的一種簡單有效策略,研究表明,嵌合肽活性的增加歸因于每種肽組分同時作用于細菌的獨特機制。為了優化具有較強細胞特異性和抗炎活性的抗菌肽,Liu等設計了基于Progetrin 1、Bovine lactoferricin的雙發卡雜合肽,發現合成的雜合肽LB-PG不僅對多種微生物表現出較高的選擇性,細胞毒性和溶血活性變低,還能抑制LPS誘導的促炎細胞因子和趨化因子的表達。雖然研究表明了雜合肽比親本肽更有效,但雜合后導致肽鏈長度增加從而帶來的合成成本增加的問題也是不容忽視的,因此需要綜合考慮多種因素的影響。
3.2 全新設計
在抗菌肽幾十年的發展過程中,為進一步降低抗菌肽的生產成本,研發活性高、序列短且組成簡單的β-折疊抗菌肽,抗菌肽的全新設計成為了研究的熱門,常見的方法有模板確立法和數據庫過濾篩選技術。
3.2.1 基于模板的設計 經過幾十年的發展,抗菌肽模板的研究已經越來越全面。模板序列可以通過比較已知的天然肽序列并篩選出對功能重要的參數獲得,與傳統的序列修飾方法相比,該方法的優點是其減少了為獲得理想結果而需要合成的肽的數量,從而在保持序列信息的同時降低研發成本。Shao等使用一種預測算法設計了最簡單的反平行β-發卡抗菌肽模板RWYXYZZRWYXY-NH,由兩條含有陽離子(X=Arg、Lys、His)和疏水殘基(Y=Val、Ile、Leu、Phe)的非共價鍵β-側鏈,通過剛性轉向區域(ZZ=Pro-Gly、D-Pro-Gly、Asn-Gly)連接而成,研發出的WKF-PG(RWFKFPGRWFKF-NH)和WRF-NG(RWFRFNGRWFRF-NH)可以通過直接的抗菌效果和抑制機體炎癥來治療細菌感染。Wu等設計了基于(XYXY)為模板的一系列肽,發現兩個β-折疊結構的水溶性多肽(IKIK)-NH和(IRIK)-NH對包括真菌在內的多種微生物表現出強殺滅作用,同時保持低毒并具有中和內毒素的能力,是真菌性角膜炎藥物的理想候選者。
3.2.2 數據庫過濾篩選 數據庫過濾篩選技術是一種利用數據庫對具有某種特定功能多肽的關鍵參數進行多步篩選,從而最終設計出具有目標活性、性能更優的全新肽的方法(圖3)。與傳統的設計方法相比,該方法可以在最短的時間內篩選出最優序列,不僅節省大量成本,還可以用來預測或進一步增強多肽的抗菌活性。隨著近年來大數據和人工智能技術的發展,基因組、蛋白質組和功能信息的數據庫已逐漸成為抗菌藥物研發不可或缺的工具。Cherkasov等迭代構建了兩個由9個氨基酸組成的隨機大型肽庫,通過Artificial Neural Networks(一種強大的機器學習技術)隨機預測超過10萬條虛擬肽的活性,篩選出的肽比目前最先進的臨床候選抗菌肽更加有效,并且能有效殺滅超級耐藥細菌;Fjell等使用遺傳算法的啟發式搜索方法生成候選肽序列,它在識別新型高抗菌能力多肽方面有很大提高。盡管這種方法被認為是有效的,但目前仍沒有明確的研究使用該方法設計全新β-折疊抗菌肽。

圖3 數據庫過濾篩選技術[69]Fig.3 Database filtering technology[69]
4 β-折疊抗菌肽的應用策略
抗生素一直是人類抗擊致病菌最重要的武器,但由于無節制的使用導致細菌耐藥問題愈發嚴重,每年都有數以萬計的人和動物因耐藥菌感染而死亡,因此世界上許多國家對抗生素的使用提出了嚴格限制。由于優良的生物學活性、低毒性、豐富的化學多樣性和天然來源的可得性,抗菌肽成為抗生素的理想替代品。經過幾十年的發展,抗菌肽已逐漸應用到畜牧、食品、化學、醫藥等許多行業。為使β-折疊抗菌肽得到更加合理的應用,切實有效的策略是必不可少的,以下論述針對目前的研究熱點,從現實角度出發,介紹了β-折疊抗菌肽能夠在未來應用轉化的兩個候選方向,分別是肽與納米載體的結合、肽與抗生素的協同應用。
4.1 肽與納米載體的結合
無論是天然還是人工合成的β-折疊抗菌肽,大多數都具有廣譜抗菌活性。傳統觀點通常認為抗菌肽所擁有的抗菌譜越廣,其應用范圍就會越大,其實不然。當一些活性較強的β-折疊抗菌肽應用到機體尤其是被動物采食進入胃腸道之后,除了起到殺滅病原微生物的功效以外,還可能會不分青紅皂白地殺死體內益生菌,造成機體菌群失調,甚至破壞健康微生物區系和免疫系統之間的動態平衡。此外,當β-折疊抗菌肽進入體內后,盡管分子內二硫鍵的存在會使它們的穩定性有所提高,但仍然需要面臨唾液、胃黏膜、血液和組織中存在的鹽離子和蛋白酶等影響,使其生物半衰期降低。而且,β-折疊抗菌肽在體內的擴散率也有限,運輸效率相對較差,致使大多數給藥方法都需要高劑量。所以如果有一種能夠更加穩定地將肽靶向運輸到體內特定病原體附近而又不會殺滅益生菌的方法,將會極大緩解這些問題,無疑會更好地加速β-折疊抗菌肽的應用進展。
近年來,傳遞和釋放生物分子的納米生物技術得到快速發展。目前,已有的肽遞送納米載體材料提供了一種有效方法,這些納米載體材料往往尺寸小、比表面積大、靶向能力強、毒性低,它們通過包裹、持續釋放和延長活性來穩定以及更好地保護β-折疊抗菌肽不受酶降解的影響,利用載體固有的靶向特點將β-折疊抗菌肽運送至特定目標以防止對有益菌造成殺傷。因此β-折疊抗菌肽結合納米材料后在體內可能會具有長效的抗菌活性、更低的毒性和更高的穩定性。一些納米顆粒(如納米纖維),它們有很大的接觸面,可以攜帶幾個單層或多層抗菌肽并將其引導到特定的目標,是β-折疊抗菌肽制劑理想的輸送系統。此外,一些無機納米材料(如金屬納米顆粒、磁性納米顆粒)以及一些有機納米材料(如脂質體納米材料、聚合物納米材料等),均可作為β-折疊抗菌肽良好的遞送載體。直徑<10 nm的顆粒會通過腎迅速代謝排出體外,>70 nm的顆粒在體內滯留時間較長,最終會到達脾被吞噬細胞清除,因此納米顆粒的尺寸范圍也極大地降低了這些顆粒從體內清除時所帶來的風險。
新的納米制劑策略的有效性仍受到生產技術、苛刻溶劑的使用以及肽的各種物理化學性質的限制。而且,用于生產這種納米載體的材料要求必須無毒、生物兼容、在加工過程中穩定且適合與肽一同使用,因此,尋找用作納米載體的新型材料也是一項需要克服的任務。
4.2 肽與抗生素的協同應用
很長一段時間來,研究人員對抗菌肽的研究主要集中在提純和鑒定抗菌肽、闡明各種肽的作用機制、生產類似的合成肽并檢測其生物物理特性以及對合成的抗菌肽進行修飾來獲得具有更強殺菌性能的多肽等方面,但目前抗菌肽的商業化仍然存在許多障礙。在對抗菌肽研究中出現的一個新理念是抗菌肽與常規抗生素的協同作用。雖然許多肽本身具有顯著的抗菌活性,但要使肽在體內發揮有效的活性往往需要更高的濃度。多肽與抗生素的混合物被發現能夠極大地提高抗微生物效果,而且導致耐藥性細菌出現的概率更低,這樣就同時避免了肽的大量使用和抗生素的高劑量使用,從而降低了成本并減少了耐藥性產生,因此肽與抗生素之間的協同作用備受關注。目前,這一策略在β-折疊抗菌肽領域已經有了成功的例子,Kim等設計了一種序列為FKVKFKVKVK-NH的β-折疊肽,他們將肽固定在聚乙二醇-聚苯乙烯樹脂上合成了多肽-樹脂結合物,當該結合物與萬古霉素和四環素混合使用時,發現溶液對金黃色葡萄球菌和藤黃微球菌的殺滅效果顯著增強;劉又銘等將LfcinB分別與土霉素、慶大霉素和阿莫西林聯合使用,驗證了LfcinB和阿莫西林聯合作用的協同效應最強,而與土霉素協同效應最差,同時發現LfcinB與抗生素的協同作用對于革蘭陰性菌的抑制效果要優于革蘭陽性菌。
目前,要完全放棄使用抗生素是不現實的,但可以將抗菌肽或擬肽類藥物與抗生素的優勢結合起來,相輔相成。在未來,β-折疊抗菌肽與抗生素優勢的結合可能會成為對抗耐藥細菌的有效手段,在減少細菌耐藥性降低成本方面發揮更大的作用。
5 小 結
過去幾十年,抗生素的過度和不當使用導致了藥物殘留和細菌耐藥性問題出現,構成了嚴重的全球健康問題,隨著各國政府一系列禁抗法令的頒布,給畜牧生產等行業造成極大沖擊,迫切需要尋求新的抗生素替代物質。抗菌肽因對多種細菌、真菌、原生動物和包膜病毒具有廣泛的活性,受到了越來越多的關注。盡管結構上的一些優勢讓β-折疊抗菌肽被認為是很有前途的飼用抗生素替代品,但是動物體內環境高濃度的蛋白酶、生理濃度鹽離子等因素仍會造成一些短鏈β-發卡抗菌肽快速失活,此外β-折疊抗菌肽還面臨著活性差異大、生產成本高等問題,阻礙其在生產中的廣泛應用。目前的方法,一方面可以通過對天然β-折疊抗菌肽進行改造或者全新設計解決β-折疊抗菌肽活性低、毒性和成本高等問題,另一方面可通過與納米載體的結合或與抗生素的協同來提高其應用潛力。這些研究進展對于促進β-折疊抗菌肽的應用起到了一定的推動作用,但關于β-折疊抗菌肽在動物體內的應用方法和應用效果均有待進一步探究。相信隨著現代分子生物學技術的不斷進步和各學科的交叉融合,在不遠的將來,β-折疊抗菌肽會成為替代抗生素的新一代飼料添加劑,為畜牧業的可持續發展提供新的動力。
