大熊貓糞便中mtDNA拷貝數與月齡的關系
2022-08-11 02:24:10任曉彤周永恒李天峰李仁貴李德生徐艷春
野生動物學報 2022年3期
任曉彤 周永恒 李天峰 李仁貴 馬 躍,4 李德生 黃 炎 徐艷春,4,5*
(1.東北林業大學野生動物與自然保護地學院,哈爾濱,150040;2.中國大熊貓保護研究中心,都江堰,611830;3.大熊貓國家公園珍稀動物保護生物學國家林業和草原局重點實驗室,都江堰,611830;4.國家林業和草原局野生動植物檢測中心,哈爾濱,150040;5.國家林業和草原局野生動物保護與利用工程技術研究中心,哈爾濱,150040)
線粒體DNA(mitochondrial DNA,mtDNA)拷貝數在其細胞能量代謝以及其他一系列細胞活動如鈣信號、鐵穩態、激素合成和細胞程序性死亡[1-3]中起著核心作用,單個哺乳動物細胞擁有成百上千個mtDNA拷貝,這種遠高于核DNA(nuclear DNA,nuDNA)的拷貝數在哺乳動物基因組復雜的進化中發揮了關鍵作用,平均每個細胞內mtDNA拷貝數(Nmpc)反映著線粒體耗竭、能量儲備和氧化應激等情況,也在一定程度上反映了機體當下的代謝狀態[4]。據此推測,機體在生長發育和衰老的過程中可能伴隨著一定規律的mtDNA拷貝數的變化。
糞便是動物的遺留物,也是動物研究中常用的非損傷性材料,無論是對野外還是飼養種群的采集都比較方便。糞便中包含的宿主DNA主要來自腸上皮細胞[5-6],機體的生長發育和衰老伴隨著腸道功能的發育和衰退,影響腸道的生理狀態和代謝能力[7],在這個過程中,作為能量供應中樞的線粒體中DNA拷貝數可能也會發生變動[8],但是,變動的方式尚不了解。
大熊貓(Ailuropodamelanoleuca)是世界矚目的珍稀物種[9-10],主要以竹子為食。竹纖維經過簡單咀嚼后直接進入胃腸道,絕大多數的竹纖維都無法被消化分解[11],經過腸道時,會黏附大量的腸道上皮細胞,并帶入糞便中,為相關研究提供足夠多、質量足夠好的DNA。人工飼養種群具有準確的出生記錄和詳細的系譜信息,本研究選用出生月份記錄清晰的圈養大熊貓的糞便為材料,研究腸道上皮細胞中mtDNA拷貝數與月齡的關系,以期揭示大熊貓不同生長發育和衰老階段腸道代謝功能的變動規律,為飼養種群和野外種群的管理提供新的視角。
1 材料與方法
1.1 樣本收集與糞便DNA提取
2021年6—7月從中國大熊貓保護研究中心的都江堰、核桃坪和耿達3個基地共采集64只11~357月齡的大熊貓的新鮮糞便(圖1)。采集時用無菌手術刀和剪刀對每個糞便樣本的頂部、尾部和中間3個部位切取表面的竹纖維,得到200~300 g的樣品,立即置于冰箱中-20 ℃暫存,在干冰中運到實驗室備用。使用QIAamp Fast DNA Stool Mini Kit(Qiagen,德國),按照說明書的流程提取糞便總DNA。
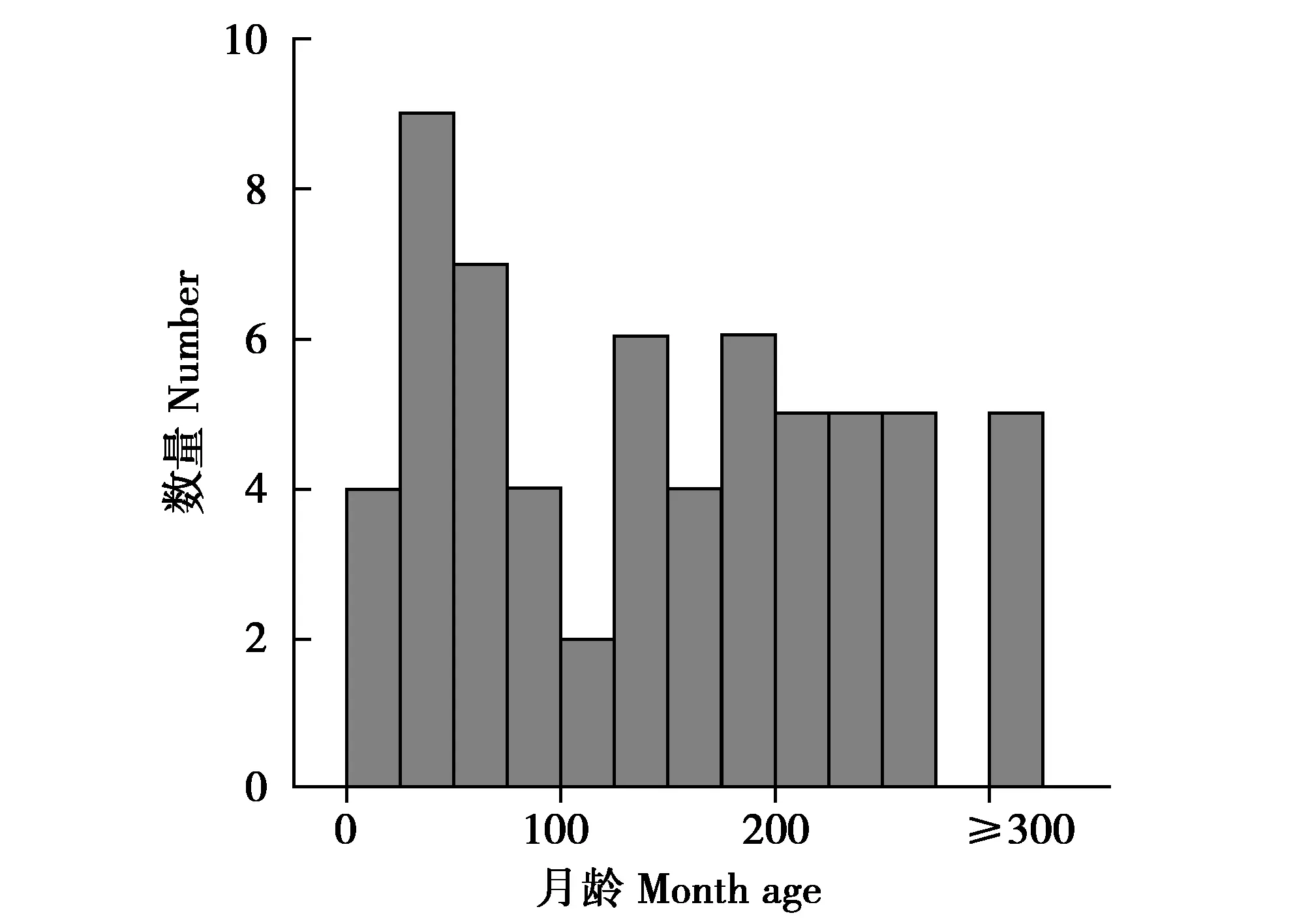
圖1 本研究采用的64只大熊貓的月齡分布
1.2 引物設計及有效性驗證
通過實時定量PCR方法測定nuDNA和mtDNA的拷貝數。分別以大熊貓nuDNA(NCBI參考序列號:NC048220.1)上β-actin基因和mtDNA(NCBI參考序列號:NC009492.1)上Cytb基因為目的基因設計引物對,同時以大熊貓基因組為比對數據庫通過NCBI-BLAST排除核內mtDNA片段(Numts)的干擾[12],最終選擇nuDNA的PCR引物對為Nβ(NβF:5′-GGGGATGGCGTTACTCATACT-3′;NβR:5′-ACATCACGAACAATCTCCCG-3′),mtDNA的PCR引物對為MC(MCF:5′-AAGTGCCCCGCCACATATTT-3′;MCR:5′-GGTCGGAATATCATGCTTCGTTG-3′)。2對引物的熔解溫度(melting temperature,Tm)均為60 ℃,片段長度分別為170、171 bp,引物由庫美生物科技有限公司合成。
2個DNA片段采用相同擴增體系和擴增程序。擴增反應在10.0 μL體系中進行,包括2 ×RapidTaqMaster Mix(南京維諾贊生物科技有限公司)5.0 μL,10 μmol/L上下游引物各0.3 μL,DNA模板2.0 μL(~40 ng總DNA),ddH2O 2.4 μL。混勻后瞬時離心,用9700型PCR擴增儀(GeneAmp,USA) 擴增。擴增程序:95 ℃預變性3 min;95 ℃變性20 s,56 ℃退火20 s,72 ℃延伸20 s,30個循環;循環結束后72 ℃延伸 5 min。從2個擴增反應中分別取4 μL的擴增產物,1%瓊脂糖凝膠電泳檢測,電泳緩沖液為1×TAE,95 V下電泳25 min。
1.3 qRT-PCR熒光定量標準品制備
1.3.1 nuDNA和mtDNA目的基因片段處理
Nβ、MC2對引物的擴增體系等比放大至40 μL,將PCR產物在2%的瓊脂糖凝膠中,按照相同的條件電泳分離。紫外燈下切取2條目的條帶,用DNA純化回收試劑盒(Axygen,美國)分別進行回收,按照說明書操作。
純化回收的DNA經超微量分光光度計(Implen,德國)檢測濃度后,與pMD18-T載體(TaKaRa,日本)連接,將質粒轉化至EscherichiacoliDH5α感受態細胞(TaKaRa,日本)中,通過藍白斑篩選隨機挑取10個白色菌落,轉移至LB液體培養基中37 ℃下搖動培養16 h。吸取菌液1 μL,用1.2中的10 μL體系進行PCR擴增以及電泳檢測。確定陽性后,用EasyPure?Plasmid MiniPrep Kit(Trans,北京)提取質粒,操作流程按照說明書進行。提取到的2種質粒用Sange法對插入片段測序,將測序結果在NCBI上比對,確認得到目的片段。
1.3.2 建立標準品濃度梯度
用所得的陽性質粒制備標準品。用微量分光光度計測定其濃度,按照公式計算質粒拷貝數(C):
C=n×60.2×1014×(1/660N)。
式中:n為質粒濃度;N為插入片段的長度。
用滅菌的ddH2O按照10×等比例梯度稀釋質粒,以稀釋10-7~10-2的質粒為模板,進行熒光定量PCR擴增以建立標準曲線,每個濃度做3個重復。熒光定量PCR反應在10.0 μL體系中進行,包括1.0 μL質粒溶液、5.0 μL TB Green Premix ExTaqⅡ(TaKaRa,日本)、10 μmol/L上下游引物各0.4 μL和3.2 μL ddH2O。反應條件:95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火34 s,72 ℃延伸30 s,40個循環;熔解段95 ℃處理15 s,60 ℃處理1 min,95 ℃處理15 s。反應用BTK-96實時熒光定量PCR分析儀(無錫百泰克生物技術有限公司)完成,繪制標準曲線。最終得到濃度為102~107拷貝/μL的標準品(Ct值10~30)。
1.4 糞便DNA中mtDNA拷貝數估算
將來自每個糞便樣品不同部位的3份DNA稀釋成近似濃度并等量混合,以此作為該個體的DNA樣品。按照1.3.2中反應體系和程序,用Nβ和MC2對引物進行擴增,檢測Ct值,并按照1.3.2中得到的標準曲線估算nuDNA和mtDNA的拷貝數。再根據公式計算平均每個腸上皮細胞中mtDNA的拷貝數(Nmpc):Nmpc=2Nmt/Nnu,式中Nmt為樣品中總mtDNA拷貝數;Nnu為樣品中總nuDNA拷貝數。每個樣品重復3次,取均值作為最終數據。
1.5 數據分析
根據3σ原則,把測得的Nmt、Nnu和Nmpc取值超出(μ-3σ,μ+3σ)區間的認定為異常值(μ為均值,σ為標準差)。檢測到的異常值在后續分析中被剔除。對整理后的數據進行統計計算,得到Nmpc的平均值(mean)、標準差(SD)和變異系數(CV);對所有個體的Nmt、Nnu和Nmpc取自然對數并做頻率分布直方圖;將個體月齡與Nmpc及ln(Nmpc)做線性回歸分析,得到兩者之間的關系;計算每個單獨數據減去群體均值之后的絕對值作為變異度,與群體月齡做線性回歸曲線。
考慮到性別對糞便DNA拷貝數的潛在影響,把Nmpc按照性別分組,分別計算雌、雄2組的平均值、標準差和變異系數;分別對雌、雄2組所有個體的Nmt、Nnu和Nmpc取自然對數并做頻率分布直方圖;由于樣品中雌性和雄性月齡分布略有差異,按照性別及月齡層次分布進行分層隨機抽樣,減小月齡分布偏好度的影響,抽樣比例設置為60%,對抽樣數據取對數處理后進行秩和檢驗;分別將雌、雄2組的月齡與Nmpc做線性回歸曲線;計算雌、雄2組的變異度,分別與群體月齡做線性回歸曲線。以上分析均采用SPSS 24.0(IBM Corp.USA)完成。
2 結果與分析
2.1 引物對Nβ和MC的有效性
以3只大熊貓糞便總DNA為模板,引物對Nβ和MC均擴增了目的片段(圖2),表明這2對引物對nuDNA和mtDNA的擴增均是有效的。用大熊貓糞便DNA進行Nβ和MC引物的熒光定量PCR檢驗,引物Nβ和MC的熔解曲線如圖3所示,熔點峰值分別為85.1、80.0 ℃,熔解曲線峰值點僅有1個且高度重合,說明引物無非特異性擴增以及二聚體,可用于樣品拷貝數的分析。
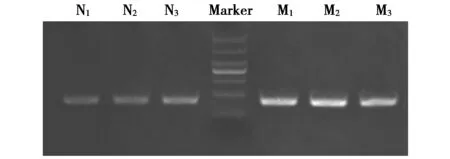
圖2 3只大熊貓Nβ和MC引物擴增產物的電泳結果
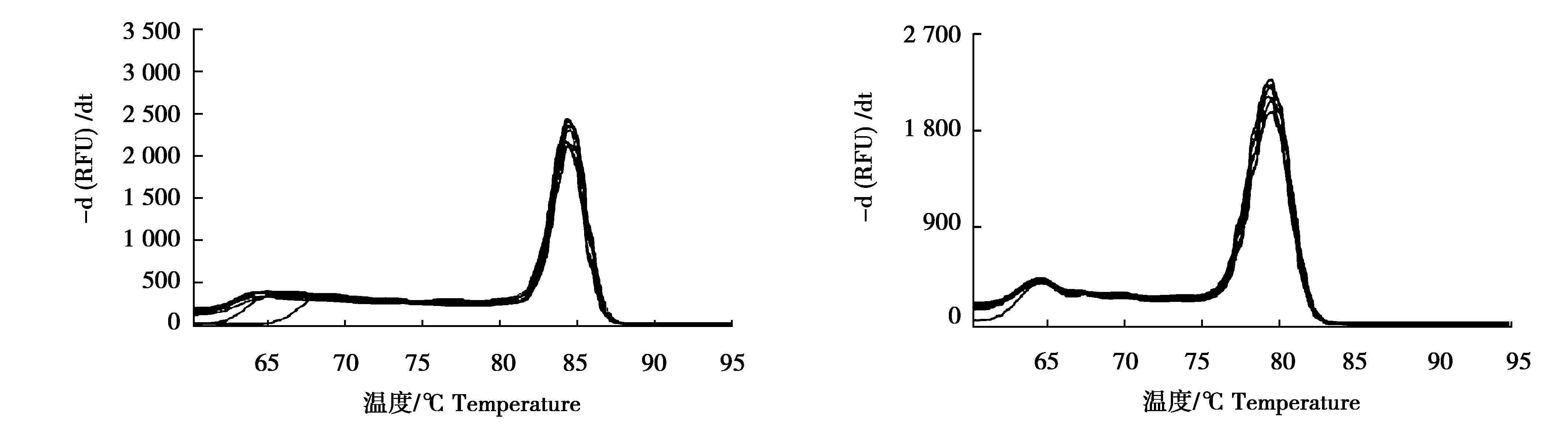
圖3 引物Nβ(左)和引物MC(右)的qRT-PCR熔解曲線
2.2 qRT-PCR標準曲線
以菌液為模板,通過引物對Nβ和MC進行PCR擴增,在瓊脂糖凝膠上檢測到在170 bp左右的條帶;測得序列與NCBI上大熊貓的參考序列NC048220.1和NC009492.1的同源性均為100%,證明插入質粒的片段是目的片段。
經微量分光光度計測定,nuDNA質粒的初始濃度為129.00 ng/μL,mtDNA質粒標準品的初始濃度為119.95 ng/μL。通過計算得到的nuDNA和mtDNA拷貝數分別為6.92×1010、6.40×1010拷貝/μL。經過10×梯度稀釋后,分別用引物對Nβ和MC的熒光定量PCR檢測Ct值,并與實際拷貝數進行回歸,繪制mtDNA和nuDNA 2種標準品的標準曲線,其中nuDNA的R2值為0.990,擴增效率(E)為97.95%,曲線方程Y=-3.811X+35.264;mtDNA的R2值為0.992,擴增效率為99.52%,曲線方程Y=-4.077X+39.063(圖4)。最終得到的拷貝數檢測下限為102。
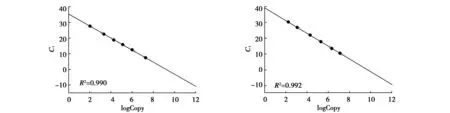
圖4 nuDNA(左)和mtDNA(右)標準品的qRT-PCR標準曲線
2.3 平均每個細胞mtDNA的拷貝數
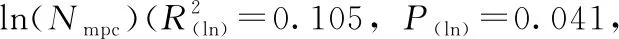
將月齡和對應的Nmpc數據按照性別劃分,雌性和雄性月齡分別為11~357、13~336月齡,平均值分別為172、123月齡;對應的Nmpc分別為24.00~7 803.00和46.00~8 467.00,平均值分別為1 177.00和1 990.00,標準差分別為1 840.34和2 449.51,變異系數分別為56.32%和123.08%;最終雌性和雄性的變異系數/平均月齡值分別為0.9%和1.0%。分別對雌、雄2組Nmt、Nnu和Nmpc進行自然對數轉換后,其頻率分布如圖5(D-F為雌性,G-I為雄性),對于雌性個體,Nmt跨度最大,Nmpc頻率分布最集中且跨度最小,Nnu頻率分布最分散;對于雄性個體,Nmt頻率分布最集中且跨度最大,Nmpc跨度最小,Nnu頻率分布最分散。對抽樣數據進行性別和Nmpc的秩和檢驗,顯著性為0.002。將每個性別的Nmpc進行自然對數轉換后與月齡進行線性回歸分析,結果顯示2個性別中Nmpc都隨著月齡的增加呈下降趨勢,但趨勢未達到顯著水平(圖7,雌性R2=0.060,P=0.176;雄性R2=0.110,P=0.074)。
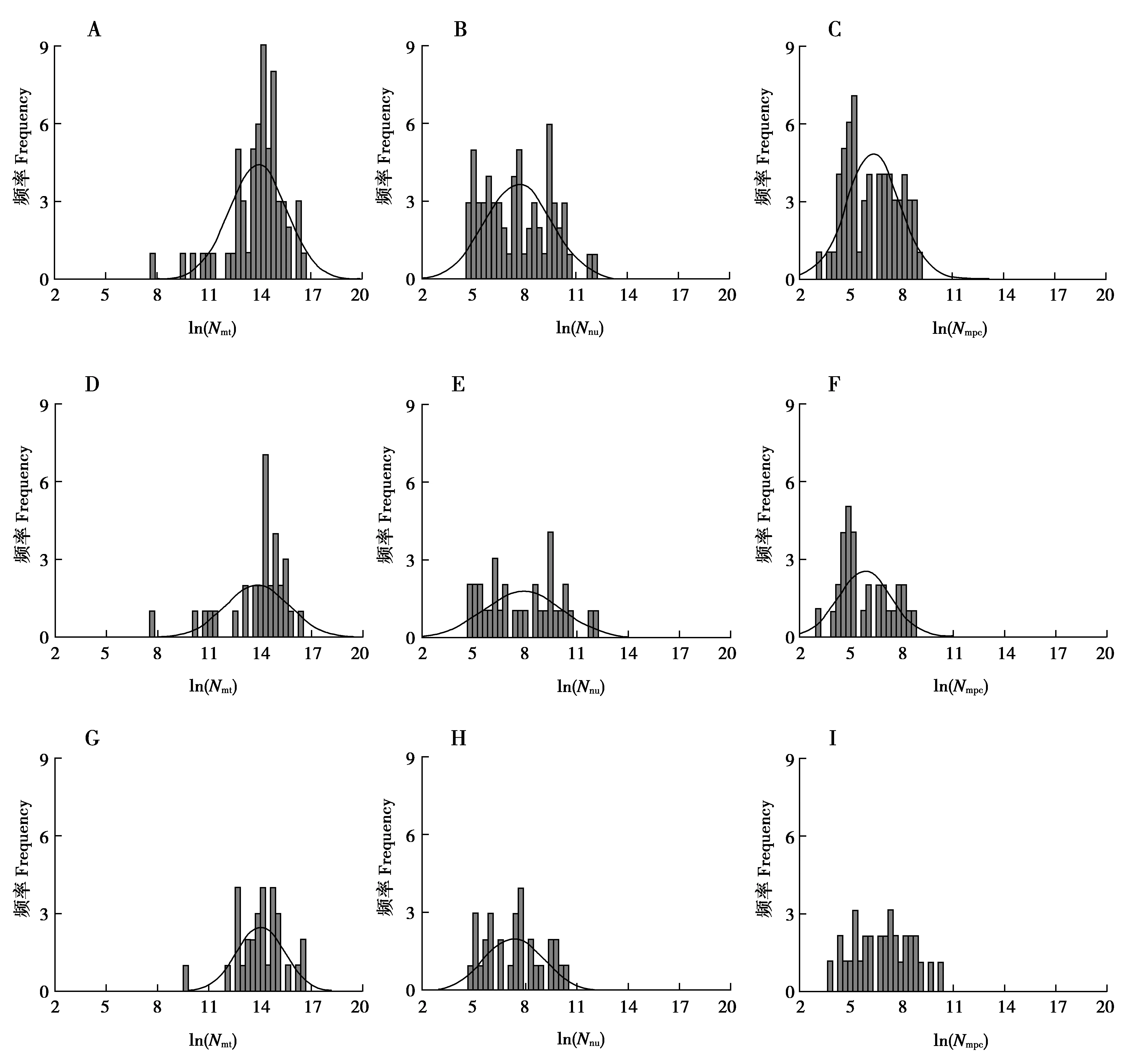
圖5 大熊貓糞便總DNA中Nmt、Nnu和Nmpc的頻率分布
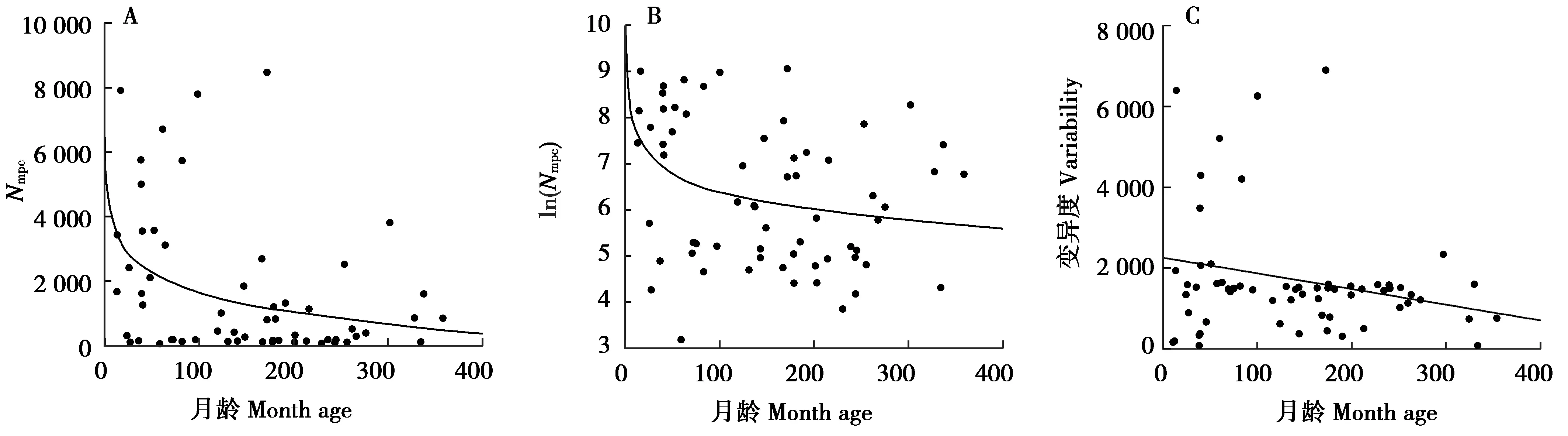
圖6 大熊貓糞便總DNA中Nmpc及其變異度與月齡的相關性
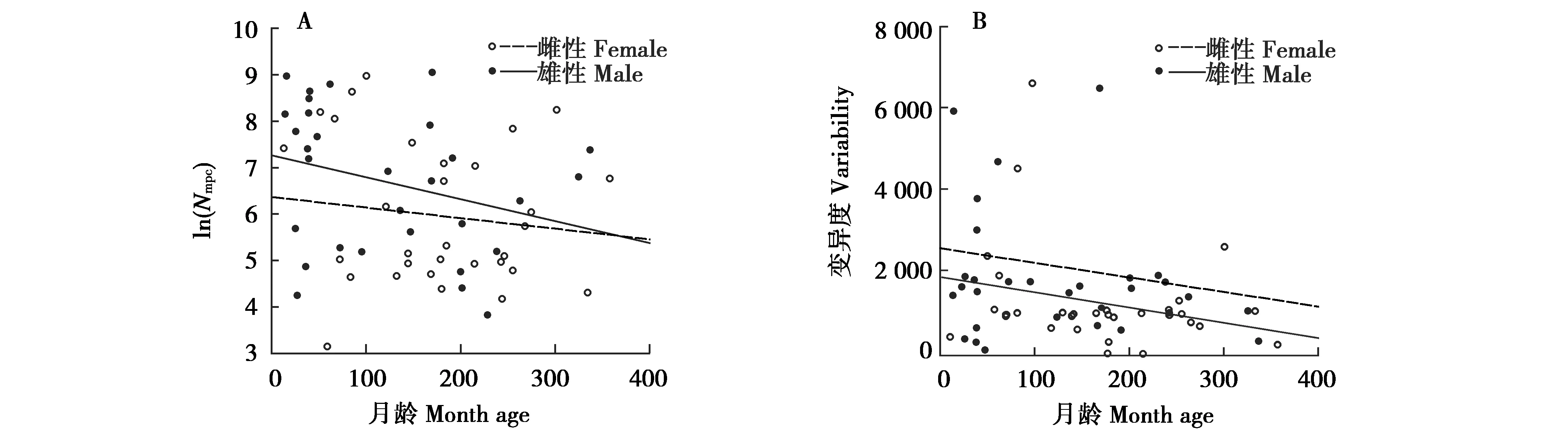
圖7 不同性別大熊貓糞便總DNA中Nmpc及其變異度與月齡的相關性
3 討論與展望
生物體內線粒體功能的發揮隨著個體生長發育而改變,長期以來,與衰老相關的線粒體功能下降一直被認為是多種生物學變化的基礎,線粒體功能改變的機制涵蓋多個領域,包括能量(ATP)產生和能量儲備的下降[13-14],自由基產生的增加[15],凋亡和有絲分裂速率的改變,以及融合、分裂的改變。這些關鍵的細胞內變化會導致細胞功能障礙、組織改變和疾病風險增加[16]。已有一些研究證明mtDNA異質性與衰老之間具有相關性[17-20],有關每個細胞的mtDNA分子的數量,即每個細胞mtDNA的拷貝數與月齡之間相關性的研究卻很少。
機體細胞mtDNA拷貝數的普遍減少會破壞線粒體的整體效能,過度增加也會帶來疾病風險[21],如人類的mtDNA拷貝數改變與癌癥、神經退行性疾病和糖尿病等衰老相關的疾病有關[22-23]。因此,從維持細胞和組織穩態的角度,mtDNA拷貝數應受到機體的嚴格控制。
本研究對大熊貓糞便中mtDNA拷貝數與月齡之間的相關性進行分析,發現mtDNA拷貝數隨著月齡的增加呈現總體下降的趨勢,并達到顯著水平(P=0.004,圖6A)。筆者推測這種趨勢可能源于mtDNA拷貝數是線粒體復制和細胞能量轉換的標志,當個體處于幼齡期時,基礎代謝率較高,線粒體的總體活動更為活躍,導致細胞中mtDNA拷貝數普遍較高;而老年個體基礎代謝水平降低[24],在這一水平上維持能量轉化的穩態所需的mtDNA拷貝數相對減少。
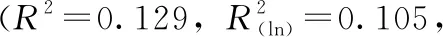

綜上所述,本研究證明大熊貓腸道脫落上皮細胞中mtDNA拷貝數隨著月齡的增加而減少,且減少速率逐漸放緩。無論老幼,拷貝數都存在著一個基本水平,反映了組織對線粒體發揮功能存在基本要求。此外,性別對于mtDNA拷貝數具有明顯的影響,同一生長階段的雄性比雌性更高。這些發現都為進一步探究腸道上皮細胞mtDNA拷貝數與生長發育和衰老之間的關系,以及利用其作為指標來評估機體的生理狀況提供了依據。同時,本研究以糞便為材料獲取腸道上皮細胞的機能狀態信息,為進一步擴大糞便的應用范圍提供了范例。
