兩種扇貝雜交和自交子一代生長性狀的比較
2022-07-20 02:24:14趙春暖蔡忠強王曉梅鄭言鑫
水產科學 2022年4期
陳 洋,趙春暖,蔡忠強,王曉梅,鄭言鑫,于 濤
(中國水產科學研究院 長島增殖實驗站,山東 煙臺 265800)
蝦夷扇貝 (Patinopectenyessoensis)原產于日本,于1982年引入中國,并迅速在山東、遼寧等地區進行養殖,是我國重要的養殖貝類之一[1]。經過多年的養殖,蝦夷扇貝大規模死亡現象頻發,長海縣作為我國最大的蝦夷扇貝增養殖基地[2],每年蝦夷扇貝死亡造成的經濟損失就達10億元[3]。研究表明,蝦夷扇貝大規模死亡除了受懸浮物、高溫刺激等極端環境因素影響外[4-5],種質退化、高密度養殖、環境變化、病原侵襲等也是我國蝦夷扇貝死亡的主要原因[6-7]。其中種質退化導致蝦夷扇貝自身抵抗環境、病原的能力下降,因此培育性狀優良的品種是解決蝦夷扇貝死亡的途徑之一。
雜交育種是培育新品種的一種傳統、有效的育種方法,是改良遺傳性狀、培育優良品種的可靠途徑。在貝類中,相關學者陸續開展了扇貝[8-10]、牡蠣[11-14]、鮑[15-19]等養殖品種種內、不同地理群體、種間等的雜交育種試驗,所得雜交種在成活率、生長速度和抗逆性等方面顯示出了良好的生產性狀和一定的雜種優勢。
楊愛國等[20-21]分別開展了櫛孔扇貝(Chlamysfarreri)(♀)×蝦夷扇貝(♂)的遠源雜交研究,雜交后代表型性狀偏向母本,但在生長速度和抗逆性方面表現出一定的雜種優勢,促進了櫛孔扇貝養殖業的健康發展。筆者以蝦夷扇貝為母本、櫛孔扇貝為父本進行雜交選育,分析子一代個體生長、存活情況,探究雜交子代推廣的價值,以期改善蝦夷扇貝種質,提高其生長、存活和抗逆能力,保障蝦夷扇貝養殖業的健康可持續發展。
1 材料與方法
1.1 試驗材料
蝦夷扇貝選自長島南隍城海域人工養殖群體,殼高(10.45±0.48) cm;櫛孔扇貝選自膠南人工養殖群體,殼高(6.73±0.32) cm。
1.2 試驗時間和地點
親本暫養、雜交及幼蟲培育試驗在中國水產科學研究院長島增殖實驗站內進行;稚貝暫養及養成在長島南隍城海域進行。
1.3 試驗設計
設計4個試驗組合:蝦夷扇貝♀×蝦夷扇貝♂(蝦夷扇貝自交組合)、蝦夷扇貝♀×櫛孔扇貝♂(正交組合)、櫛孔扇貝♀×櫛孔扇貝♂(櫛孔扇貝自交組合)、櫛孔扇貝♀×蝦夷扇貝♂(反交組合)。
1.4 試驗方法
1.4.1 親貝暫養
采用浮動網箱暫養,暫養密度30~50個/m3;前期每日倒池1次,后期每日早、晚各換水1次,每次換水1/3;餌料和升溫方式為:蝦夷扇貝采用自然水溫暫養,投喂三角褐指藻(Phaeodactylumtricornutum),經過至少2次嚴格的挑選后將雌體和雄體分開;櫛孔扇貝采用逐步升溫的方法進行暫養促熟,餌料以三角褐指藻為主,搭配球等鞭金藻(Isochrysisgalbana)和螺旋藻(Spirulina)進行營養強化,經過至少2次嚴格的挑選后將雌體和雄體分開。
1.4.2 產卵、受精、孵化
性腺充分發育成熟后,再次確認同一群體無雌雄混雜。采用陰干、升溫、間斷充氣的方法誘導蝦夷扇貝、櫛孔扇貝雌體在池中產卵,同時將蝦夷扇貝、櫛孔扇貝雄體分別放于2個玻璃缸水槽中排精。將精、卵混合受精后,每個卵子周圍有3~4個精子,連續取樣觀察胚胎發育情況,直到發育至D形幼蟲。
1.4.3 幼蟲培育
當80%受精卵發育至D形幼蟲,立即用虹吸法選優進行培養。培育適宜密度為8~12個/mL;光照低于500 lx;餌料以球等鞭金藻為主,日投喂量為1.5×104~5.0×104個/mL,日投喂3~6次;日換水2次,每次換水1/2,每5 d倒池1次;培育水溫:蝦夷扇貝自交組、正交組培育水溫16 ℃,櫛孔扇貝自交組、反交組培育水溫19 ℃。
培育期間,每日檢測試驗組幼蟲生長情況,包括殼高、密度等,以確定其生長速度和成活率。
1.4.4 附著基投放
當幼蟲眼點率達到30%~40%時,投放附著基。
1.4.5 稚貝暫養及養成
將附有稚貝的附著基放入60目網袋中,垂掛于海區的浮綆上,殼長達0.3 cm時分入20目網袋中,稚貝殼長約1 cm時裝入扇貝籠養殖,完成稚貝的海區中間培育。
1.5 數據分析
受精率、孵化率、幼蟲生長及存活中的相關數據計數使用生物顯微鏡(重慶奧特B204LED)。稚貝及成貝的殼高測量使用游標卡尺。
各指標計算公式如下:
R1=n2/n1×100%
(1)
R2=n4/n3×100%
(2)
R3=n6/n5×100%
(3)
R4=n8/n7×100%
(4)
式中,R1為受精率(%),n1為所有卵子數(個),n2為出現第一極體的卵子數(個),R2為孵化率(%),n3受精卵數(個),n4為孵化出的D形幼蟲數(個),R3為幼蟲期存活率(%),n5為第1天的幼蟲數(個),n6為測量日存活的幼蟲數(個),R4為稚貝期存活率(%),n7為掛入海區的稚貝總數(個),n8為測量日存活稚貝數(個)。
幼蟲期,于第1、3、5、7、9、12、15、18、20天取樣,每組每次取樣30個個體,于顯微鏡下測量殼長,并統計存活率。
稚貝海區中間培育及養成期,于1月齡至8月齡,每月每組取樣30個個體,并在11月齡分苗、15月齡收獲時分別取樣,測量殼高,并統計存活率。
用Excel軟件對各試驗數據進行整理,采用SPSS 19.0軟件統計分析軟件進行單因素方差分析及鄧肯多重比較,結果用平均值±標準差形式表示。
2 結 果
2.1 受精率、孵化率比較
自交、雜交組合均可完成受精過程,受精率為97.19%~98.15%;自交組合略高于雜交組合,以蝦夷扇貝為母本的組合略高于以櫛孔扇貝為母本的組合,但單因素方差分析及鄧肯多重比較結果顯示,相互之間差異不顯著(P>0.05)(表1)。
自交、雜交組合的孵化率也較高,為89.09%~92.76%;自交組合略高于雜交組合,以櫛孔扇貝為母本的組合略高于以蝦夷扇貝為母本的組合,但單因素方差分析及鄧肯多重比較結果顯示,組合間差異不顯著(P>0.05)(表1)。
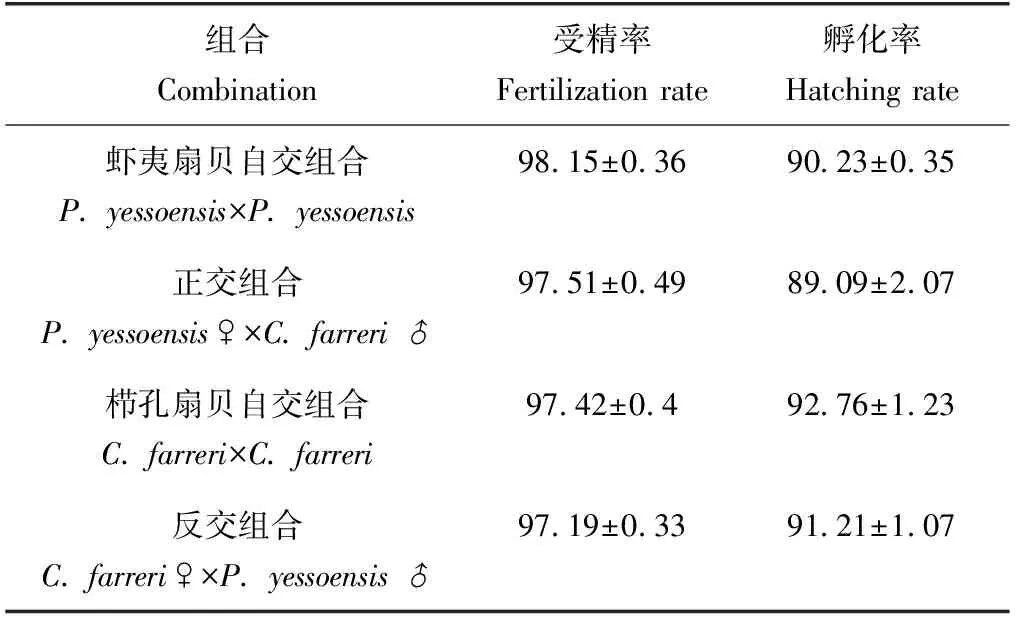
表1 各組合的受精率和孵化率 %
2.2 幼蟲期生長和存活情況比較
孵化后至7日齡時,幼蟲規格無顯著差異(P>0.05);9日齡開始幼蟲生長差異逐漸顯現出來,雜交組合明顯快于自交組合(表2、圖1)。
以櫛孔扇貝為母本的組合,櫛孔扇貝自交、反交組合在15日齡時投放附著基,反交組合的眼點幼蟲平均規格為196.27 μm,而櫛孔扇貝自交組合的眼點幼蟲的規格為189.20 μm,雜交組合較自交組合大3.74%;櫛孔扇貝自交、反交組合的生長日平均增長量為5.76、6.28 μm/d,雜交組合較自交組合提高9.03%。
以蝦夷扇貝為母本的組合,蝦夷扇貝自交、正交組在20日齡時投放附著基。正交組合的眼點幼蟲平均規格為263.10 μm,而蝦夷扇貝自交組合的眼點幼蟲的規格為252.20 μm,雜交組合較自交組合大4.32%;蝦夷扇貝自交、正交組合的生長日平均增長量為8.16、8.57 μm/d,雜交組合較自交組合提高5.02%。
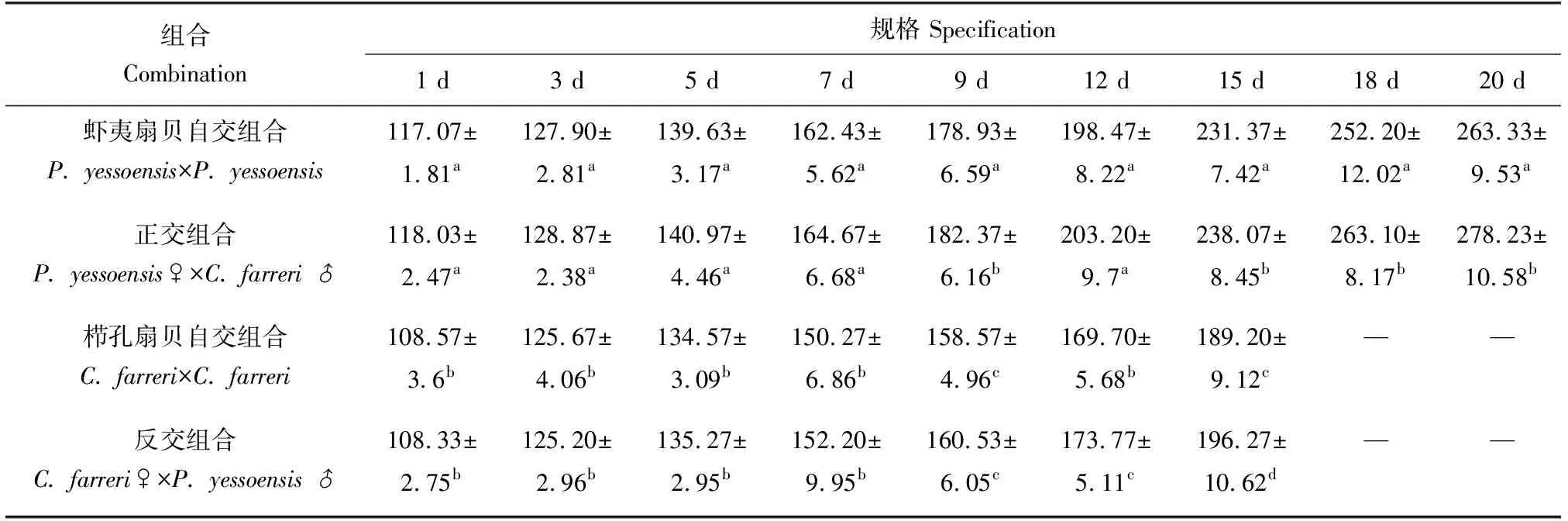
表2 幼蟲期各組合生長情況 μm
圖1 幼蟲期不同組合生長情況Fig.1 Growth at different larval stages in different combination
幼蟲階段,櫛孔扇貝自交、反交組合的存活率分別為63.09%、64.83%,相互之間差異不顯著(P>0.05);蝦夷扇貝自交、正交組合存活率相對較高,分別為70.07%、69.16%,但相互之間差異不顯著(P>0.05)(表3、圖2)。
2.3 附著變態率和海區保苗率的比較
櫛孔扇貝自交、反交組在15日齡時投放附著基,蝦夷扇貝自交、正交組合則在20日齡時投放附著基,4個組合幼蟲均在一周內完成附著變態過程。蝦夷扇貝自交、正交組合變態率較櫛孔扇貝自交、反交組合高,其中蝦夷扇貝自交附著變態率最高,達48.28%,櫛孔扇貝自交、反交組合變態率約為45%(表4),但單因素方差分析及鄧肯多重比較結果顯示相互之間差異不顯著(P>0.05)。
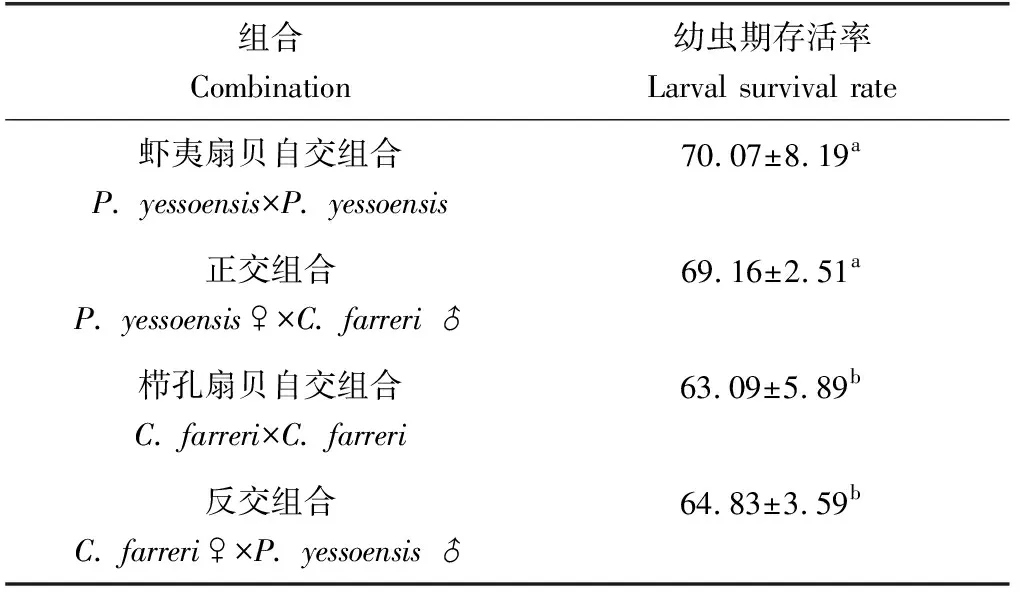
表3 幼蟲期各組合存活率情況 %
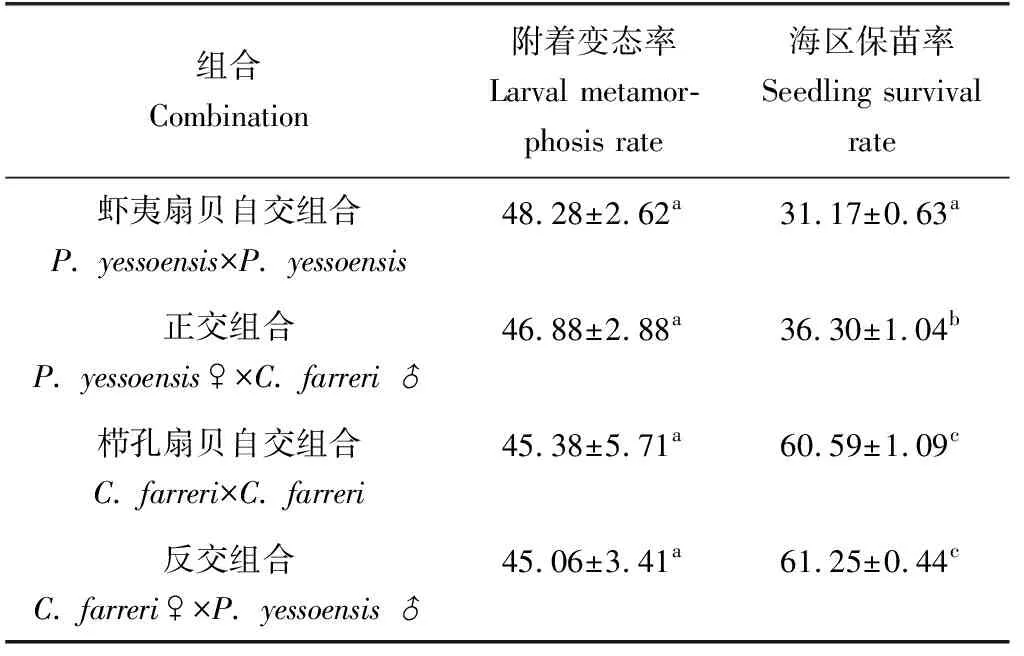
表4 各組合附著變態率與海區保苗率情況 %
以櫛孔扇貝為母本的櫛孔扇貝自交、反交組合的海區保苗率明顯高于以蝦夷扇貝為母本的蝦夷扇貝自交、正交組合,其中櫛孔扇貝自交、反交組合的保苗率分別為60.59%、61.25%,相互間差異不顯著(P>0.05),蝦夷扇貝自交、正交組合的海區保苗率分別為31.17%、36.3%,雜交組合顯著高于自交組合,提高5.13個百分點。
2.4 養成期生長和存活情況比較
生長至8月齡時,櫛孔扇貝自交、反交組合的殼高分別達到33.49、35.97 mm(表5、圖3),組合間差異顯著且雜交組合較自交組合生長速度提高7.41%;8月齡時,櫛孔扇貝自交、反交組合生長月增長量為4.70、5.05 mm/月,組合間差異顯著且雜交組合較自交組合月增長量提高7.45%。
生長至15月齡時,蝦夷扇貝自交、正交組合的殼高分別達到64.30、75.47 mm(表5、圖3),組合間差異顯著且雜交組合較自交組合生長速度提高17.37%;15月齡時,蝦夷扇貝自交、正交組合生長月增長量明顯較高,為4.55、5.34 mm/月,組合間差異顯著且雜交組合較自交組合月增長量提高17.36%。
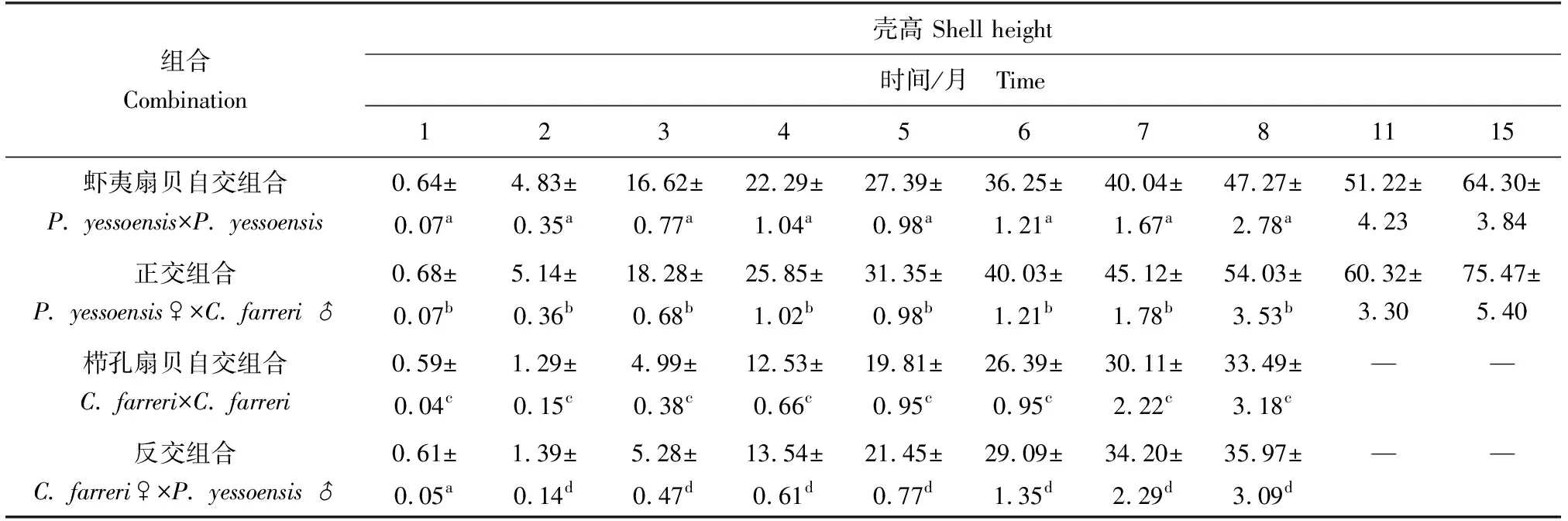
表5 養成期各組合生長情況 mm
第一次分苗時,櫛孔扇貝自交、反交組合的存活率分別為87.42%、89.35%,組合間差異不顯著(P>0.05);第二次分苗時,櫛孔扇貝自交、反交組合的存活率分別為82.69%、87.45%,雜交組合顯著高于自交組合(表6)。反交組合總的存活率為78.14%,櫛孔扇貝自交組合總的存活率為72.29%,雜交組合較自交組合提高5.85個百分點。
第一次分苗時,蝦夷扇貝自交、正交組合的存活率分別為96.44%、98.50%,組合間差異不顯著(P>0.05);第二次分苗時,蝦夷扇貝自交、正交組合的存活率分別為87.57%、92.22%,雜交組合顯著高于自交組合;第三次分苗時,蝦夷扇貝自交、正交組合的存活率分別為81.23%、89.45%,雜交組合顯著高于自交組合;第四次分苗時,蝦夷扇貝自交、正交組合的存活率分別為71.07%、82.66%,雜交組合顯著高于自交組合。整個養成期間,正交組合的總存活率為67.16%,而蝦夷扇貝自交組合的存活率為48.75%,雜交組合較自交組合提高18.41個百分點。
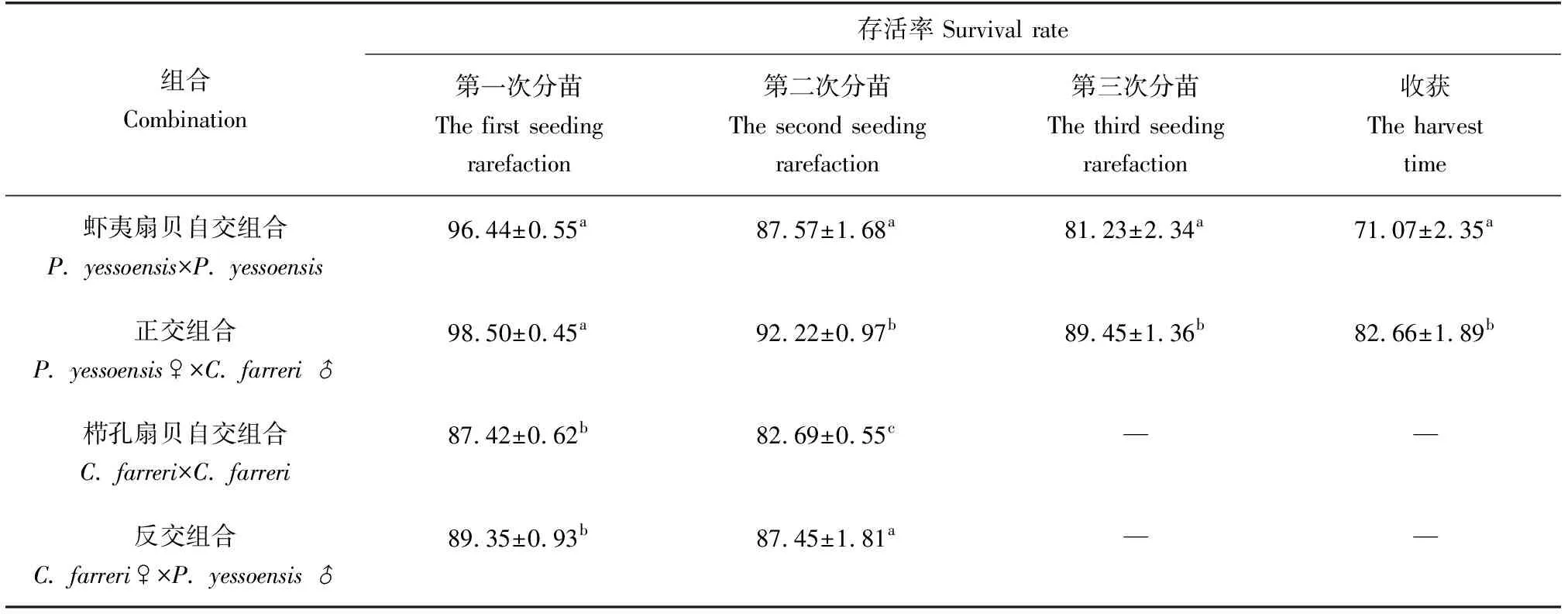
表6 不同時期各組合存活率情況 %
3 討 論
3.1 蝦夷扇貝與櫛孔扇貝雜交的可行性
蝦夷扇貝和櫛孔扇貝為扇貝科扇貝屬的兩個不同種,二者染色體同為2n=38條[22-24],這就為二者進行遠緣雜交提供了可能。周麗青等[25-27]研究表明:在染色體數目上,蝦夷扇貝和櫛孔扇貝正、反交子代早期胚胎的染色體數目為38條,與其雙親一致;在染色體構成上,絕大部分正、反交子代分別繼承了櫛孔扇貝和蝦夷扇貝各一套染色體,為真正的雜交種。本研究表明,二者正反交均可進行,后期雜交子代均可正常生長,這也與楊愛國等[20-21]的研究結果一致。
3.2 遠緣雜交對受精率、孵化率的影響
本試驗中,4個組合的受精率均在97%以上,組合間差異不顯著(P>0.05),這也與楊愛國等[20-21]的研究結果一致。劉桂龍等[28]研究蝦夷扇貝與風向標扇貝(P.caurinus)種間雜交時發現,雜交組合的受精率與自交組合有顯著差異;而南樂紅等[29]在紫扇貝(Argopectenpurpuratus)與墨西哥灣扇貝(A.irradiansconcentricus)雜交的研究中發現,雜交組的受精率與自交組差異不顯著(P>0.05),甚至部分雜交組的受精率和孵化率高于自交組。這些研究結果與陳來釗等[30]的觀點一致,即父母本親緣關系的遠近一般與受精無直接的聯系。
通常,孵化率的高低取決于卵子的成熟度,對雜交胚胎來說,還取決于雙親之間差異的程度,即親和性大小[30]。呂振明等[27]研究表明,櫛孔扇貝與蝦夷扇貝雜交子代中兩套染色體具有較強的親合性,與雜交子代具有較高的孵化率結果吻合。
3.3 子代性狀表現
楊愛國等[20]研究表明,蝦夷扇貝(♀)×櫛孔扇貝(♂)的雜交子一代僅表現出存活率優勢,生長速度差異不顯著(P>0.05)。于瑞海等[21]研究表明,蝦夷扇貝(♀)×櫛孔扇貝(♂)的雜交子一代表現出一定的生長優勢,而成活率則差異不顯著(P>0.05)。
本試驗結果顯示:蝦夷扇貝(♀)×櫛孔扇貝(♂)的雜交子一代在幼蟲期的存活率未表現出顯著性差異(P>0.05),但在9日齡后表現出一定的生長優勢;附著變態率雖然無顯著差異(P>0.05),但海區保苗率提高5.13個百分點;在養成階段,雜交子一代表現出明顯的雜種優勢,生長速度提高了17.37%。
3.4 遠緣雜交對子代存活率的影響
存活率是良種選育的另一個重要指標,存活率的高低反映了子代對環境的適應能力,對最終產量也有很大的影響[31]。雜交除了對子代生長速度有一定的影響,子代間的存活率也存在一定的差異。楊愛國等[20]研究表明,蝦夷扇貝(♂)×櫛孔扇貝(♀)的雜交子一代的存活率在夏季高溫季節明顯提高;鄭懷平等[32]研究表明,兩個海灣扇貝(A.irradians)養殖群體雜交組的存活率都高于相應的自交組;豐瑋等[33]研究表明,紫扇貝與海灣扇貝雜交后代的存活率顯著高于自交組。關于貝類遠緣雜交的研究也表明,雜交種不一定全部具有雜種優勢,雜交過程中會出現配子不兼容、幼蟲存活率低和幼蟲不變態等遠交衰退現象[34]。張躍環等[35]進行的長牡蠣(Crassostreagigas)與近江牡蠣(C.ariakensis)種間雜交結果表明,在存活率方面,以長牡蠣為母本的雜交組具有顯著的優勢,而反交組則表現為劣勢。本研究結果表明,幼蟲期各組合的存活率和附著變態率無顯著差異(P>0.05),而養成期反交、正交組合的存活率較櫛孔扇貝自交、蝦夷扇貝自交組合分別提高5.85、18.41個百分點,雜交組合表現出明顯的雜種優勢。
4 結 論
筆者建立了蝦夷扇貝、櫛孔扇貝自交、雜交組合,比較了每個組合的生長和存活性狀,結果顯示,正反交組合較自交組合均具有一定的雜種優勢,且蝦夷扇貝(♀)與櫛孔扇貝(♂)雜交獲得的子一代雜種優勢更加顯著。雖然雜交子一代外部形態與蝦夷扇貝相同,但顯著提高了蝦夷扇貝的生產性能,遠緣雜交育種是解決蝦夷扇貝大面積死亡的重要途徑之一,正交子一代已經具有生產使用價值,適于大面積推廣,提高蝦夷扇貝養殖的良種覆蓋率和養殖效益。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
