角蟬觸角及其感器的掃描電鏡觀察
2022-07-15 06:26:08孫詠澤于瑞濤李澤晨胡凱楊宗武袁向群
西北農業學報 2022年6期
孫詠澤于瑞濤李澤晨胡 凱楊宗武袁向群
(西北農林科技大學 植物保護學院,陜西楊凌 712100)
觸角作為接受外來化學信號的主要器官[1-3],在昆蟲種間及種內的化學通信、聲音通信和觸覺通信中扮演著重要的角色[4-6],其感受功能主要通過著生于其上的多種感器實現[2,7]。感器由昆蟲體壁的部分表皮特化而來,包括外部表皮結構、內部神經元以及相關連的鞘細胞和一些附屬結構,可對周圍環境和昆蟲種群內部各種刺激做出反應,是昆蟲感知機械和化學信號的基本結構和功能單位,在昆蟲的雌雄交配、寄主選擇和信息通訊等行為中起著關鍵作用[8-10],觸角感器的種類、數量與分布關乎昆蟲行為及對各種環境刺激因子的反饋,尤其對昆蟲的寄主定位與辨認、取食與繁衍、棲息與遷徙影響甚大[11]。
1964年Schneider[4]首次對昆蟲觸角上不同類型感器的表面形態結構特征進行分類描述;Zacharuk[12]于1980年初次聯合掃描電鏡和透射電鏡技術研究昆蟲觸角感器的超微結構,其后伴隨電子顯微技術的蓬勃發展,掃描電鏡和透射電鏡技術被很多專家學者普及到許多不同類群的昆蟲觸角感器研究之中。40余年來,國內學者運用掃描電鏡技術圍繞鱗翅目、膜翅目以及半翅目昆蟲等的觸角感器做了諸多研究。尹文英等[13]于國內第一次使用掃描電鏡觀察棉紅鈴蟲的觸角感受器;袁向群等[14-15]利用掃描電鏡開展弄蝶科5種弄蝶的觸角超微結構研究,描述了各種感器的形態結構及其分布,并討論了感器的功能;黃珍富[16]將掃描電鏡觀察到的稻弄蝶觸角感器超微結構應用于探究稻弄蝶屬的系統學及其進化方式中;李虹雨等[17]使用掃描電鏡觀察金環胡蜂兩亞種的觸角感器。此外,使用掃描電鏡觀察半翅目葉蟬科昆蟲觸角感器的研究也不斷有報道。高景林等[18]觀察到假眼小綠葉蟬觸角上僅存在一種毛形感器,且其數量、分布在雌雄二性間相近;趙立嵚等[19]觀察到條沙葉蟬的觸角感器類型與數量在雌雄蟲間不存在差異,其觸角感器僅包括毛形感器和錐形感器兩種,梗節上僅有毛形感器;于凱等[20]對柳寬突葉蟬和黑紋片角葉蟬的雌、雄性成蟲感器進行觀察,發現兩種葉蟬觸角感器數量都較少,同種雌、雄蟲之間也不存在明顯差異。掃描電鏡技術能發現各種感器表面細微結構的差異,探究其種類、數量、分布,這不僅有助于人們進一步探索昆蟲觸角感器與昆蟲行為之間的關系,且有助于探究昆蟲與自然界間的化學通信模式,并為分類學和昆蟲系統發育分析提供更多新的線索與依據[13-20]。
角蟬科(Membracidae)屬于半翅目(Hemiptera)角蟬總科(Membracoidea),同葉蟬科(Cicadellidae)一致。中國角蟬共計281 種,均為植食性,部分種類是重要的經濟植物害蟲,目前已知寄主植物的角蟬共80 種,已明確的寄主植物達116種,主要為雙子葉植物。角蟬對植物的危害方式與大部分葉蟬科害蟲相似,主要體現在:①刺吸植物根、莖、葉或果實的汁液,使植物萎蔫、失綠,影響植株的生長;②取食同時分泌蜜露,污染植株葉面,影響植株呼吸作用和光合作用;③傳播植物病毒病;④產卵過程中刺傷植物組織,導致枝條易折。褐三刺角蟬Tricentrus brunneusFunkhouser和黑圓角蟬Gargara genistae(Fabricius)均屬角蟬科露盾角蟬亞科(Centrotinae),二者均為中國角蟬常見種。其中,褐三刺角蟬主要分布于中國陜西、甘肅、山東、廣西、貴州等6 個省(區),寄主植物為構樹、刺槐和胡頹子,成、若蟲散居;而黑圓角蟬廣布于除青海以外的中國各省,目前已傳入美國一些州,寄主植物多達17種,分屬茄科、豆科、桑科、蕓季科與菊科等10個科[21]。
自2002年至今,國內關于葉蟬科昆蟲觸角感器的研究時有報道;但國內外對角蟬的研究卻始終以系統學、行為學、生物學和生態學為主,其次為形態學,罕有關于其觸角及感器的報道。因此,本研究觀察比較中國兩種常見角蟬褐三刺角蟬T.brunneus和黑圓角蟬G.genistae成蟲雌雄二性的觸角及其感器形態、數量與分布,以期為角蟬科昆蟲后續系統發生學、行為生物學、化學生態學和電生理學等研究,以及對角蟬的生物學防治措施提供觸角及感器方面的新線索與新依據。
1 材料與方法
1.1 供試昆蟲
所用褐三刺角蟬和黑圓角蟬成蟲標本均為西北農林科技大學昆蟲博物館館藏干標本。分別取雌、雄成蟲若干頭作為供試標本。
1.2 掃描電鏡樣品制備及觀察
步驟為:①將黑圓角蟬和褐三刺角蟬分別在解剖鏡下隨機選取觸角完整的雌、雄個體各5頭;②向試蟲容器中倒入丙酮,用超聲波清洗儀振動清洗3 min,更換丙酮后再次清洗,重復3次;③清洗后將試蟲在70%酒精中浸泡15 min,再在解剖鏡下將觸角連頭一起取下;④待酒精揮發后將這些連頭觸角分不同面粘于載物臺上,將粘好的載物臺放入抽真空干燥器中干燥,備用;⑤用離子濺射儀(日立E-1045)噴金鍍膜3 min;⑥置于電子掃描儀(日立S-4800)中于20 k V 加速電壓下進行觀察并拍照。
2 結果與分析
2.1 褐三刺角蟬觸角的一般特征
2.1.1 觸角外部形態及結構組成 對褐三刺角蟬雌、雄性成蟲的觸角進行觀測發現,在外部形態方面,褐三刺角蟬雌、雄蟲的觸角無明顯差異,均為剛毛狀;在結構組成方面,雌、雄成蟲的觸角均包含柄節(scape)、梗節(pedicel)和鞭節(flagellum)3部分,整體長度約771.96μm(圖1-A)。
柄節近似倒鐘狀,最短,較粗壯,長度約64.36μm,基部包被于膜質的觸角窩內,其上有點粒狀突起,這些點狀突起大部分大小均勻,部分點狀突起合并形成較大的不規則突起(圖1-B)。梗節連接于柄節端部洼陷的窩內,形似鳳梨,長度約92.60μm,兩端較細而中部較粗,基部較端部稍粗,中部著生錐形感器和毛形感器,其表面遍布鱗片狀的突起(圖2-B)。鞭節形態細長,基部略呈紡錘形,且分為許多亞節,長度約615.00μm,遠長于柄節與梗節,約占觸角總長的80%。前8個亞節相對較膨大,其后各亞節趨于變細。第1亞節基部縊縮,同梗節頂端凹陷處相連接,端部膨大;感器集中分布于第3、4、5、6、7亞節,其余各節均無感器(圖2-E)。

圖1 褐三刺角蟬成蟲觸角的掃描電鏡觀察Fig.1 SEM of adult T.brunneus antennae
2.1.2 感器的種類、形態和分布 褐三刺角蟬的觸角感器僅存在毛形感器和錐形感器2種,數量較少,且雌雄成蟲間的觸角結構及感器分布基本相同,無明顯差異。
毛形感器(Sensilla trichodea,St):整體形態呈纖毛狀,端部較為尖細,略有彎曲,著生于梗節中部,其基部位于表皮上隆起的圓形火山狀凹窩中。梗節上的毛形感器長28.77~34.50μm,端部與觸角表面的夾角小于30°(圖2-B~2-D)。
錐形感器(Sensilla basiconica,Sb):整體形態呈圓錐狀,其較毛形感器稍短稍粗,端部較為圓鈍,不彎曲,在梗節和鞭節上都有分布。梗節上的錐形感器長約25.94μm,端部與觸角表面的夾角不足30°,鞭節上的錐形感器長12.76~16.47 μm,端部與觸角表面的夾角為25°~50°,位于梗節上的錐形感器較鞭節上的更長,鞭節上的錐形感器集中在鞭節的一側,自表皮隆起的圓形火山狀凹陷中發生(圖2-B~2-G)。
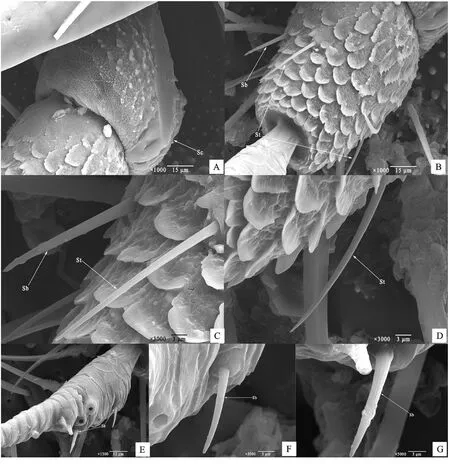
圖2 褐三刺角蟬成蟲觸角感器的掃描電鏡觀察Fig.2 SEM of adult T.brunneus antennal sensilla
2.2 黑圓角蟬觸角的一般特征
2.2.1 觸角外部形態及結構組成 對黑圓角蟬雌、雄性成蟲的觸角進行觀察發現,雌、雄蟲的觸角形態上無差異,均呈剛毛狀;在結構組成方面,雌、雄成蟲的觸角均包含柄節,梗節和鞭節3 部分,長度約660.4μm(圖3-A)。柄節近似鐘形,最短,較粗壯,長度約89.5μm,基部包被于膜質的觸角窩內,其上具點粒狀和刺狀突起,其中點狀突起大部分大小相近,部分刺狀突起由2個突起合并而成。梗節連接于柄節端部洼陷的窩內,呈較為規則的鳳梨狀,長度約100.0μm,兩端較細,近基部較粗,中部著生有錐形感器和毛形感器,其表面似洋薊花蕾,遍布鱗片狀的突起(圖4-B)。鞭節形態細長,基部似紡錘形,長度約為470.9 μm,遠長于柄節與梗節長度之和,且分為許多亞節。基部10 個亞節較膨大,其后各亞節逐漸變細。第1亞節基部縊縮,同梗節頂端凹陷處相連接,端部膨大;感器集中分布于第7、8、9、10亞節,其余各節均無感器(圖4-F)。

圖3 黑圓角蟬成蟲觸角的掃描電鏡觀察Fig.3 SEM of adult G.genistae antennae
2.2.2 感器的種類、形態和分布 黑圓角蟬的觸角感器僅存在毛形感器和錐形感器2種,數量較少,而且雌雄成蟲間的觸角結構及感器分布基本相同,無明顯差異。
毛形感器(Sensilla trichodea,St):整體形態呈纖毛狀,端部較為尖細且存在不同程度的彎曲,長度也存在差異,僅觀測到在梗節各面分布,主要著生于梗節中部。毛形感器基部位于表皮上隆起的圓形火山狀凹窩處,長24.19~35.80μm(圖4-C,4-E)。
錐形感器(Sensilla basiconica,Sb):整體形態呈圓錐狀,其較毛形感器稍短稍粗,端部較為圓鈍,幾乎不彎曲,其在梗節和鞭節上均有分布,鞭節上僅有錐形感器分布且偏向集中在鞭節的一側,著生于表皮隆起的圓形火山狀凹窩中。其中梗節表面的錐形感器長約30.22μm,鞭節表面的錐形感器長19.41~33.06μm(圖4-D、4-G、4-H和4-I)。

圖4 黑圓角蟬成蟲觸角感器的掃描電鏡觀察Fig.4 SEM of adult G.genistae antennal sensilla
3 討論
本研究對角蟬科褐三刺角蟬和黑圓角蟬的觸角超微結構進行了掃描電鏡觀察。結果表明,兩種角蟬觸角形態相似,均為典型的剛毛狀觸角,包括柄節、梗節、鞭節3部分,在梗節中部的一周各面均著生有少量毛形感器和錐形感器,鞭節的前10個亞節部分側面上著生有部分錐形感器,與已報道的同為角蟬總科下部分葉蟬科昆蟲的觸角形態和分節及感器類型和分布有一定相似之處[20,22-25]。另外,黑圓角蟬和褐三刺角蟬雌雄成蟲的觸角感器差異不明顯,這點也同之前對葉蟬科昆蟲的研究結論一致[10]。該發現可能表明在未來對角蟬觸角的研究中,可對目前報道更多的葉蟬觸角的研究方法進行一定程度的借鑒。褐三刺角蟬和黑圓角蟬觸角柄節上有許多刺突狀突起,梗節上均有鱗片(耳)狀突起,這在其他半翅目頭喙亞目的昆蟲中也有發現[23,26],與山打根擬穎蠟蟬[27]和條沙葉蟬[23]等昆蟲相比較,黑圓角蟬和褐三刺角蟬的多種突起同樣缺少感覺臼形窩這一構造,這表明這些突起并不是感覺器,而應為表皮突起特化而成的特殊構造。
黑圓角蟬和褐三刺角蟬的梗節上均有毛形感器分布。毛形感器是昆蟲觸角上分布最為廣泛、數目最多的感器,其毛發長度變化很大,而且可以自由活動,在本試驗所得的掃描電鏡圖中也可見不同程度的彎曲,且這些差異在所有已報道的昆蟲觸角上均有發現[28-31]。相關研究表明,毛形感器在功能上可以是機械性的、接觸性的、熱敏感的或嗅覺的,是昆蟲感受性信息素的重要器官[12,32],其中表面無孔的一般被認為是機械感器[33],表面分布有納米微孔的被認為是嗅覺感器,這些納米孔由生物膜形成,孔徑為1~100 nm,起到過濾氣味分子的作用[34],目前已有在半翅目異翅亞目昆蟲口器的毛形感器上發現納米孔的報道。本研究中黑圓角蟬梗節的毛形感器上可見部分點狀凹陷,可能意味著這種毛形感器具嗅覺感器的特點,未來也可對其進一步探究。結合本研究結果,黑圓角蟬和褐三刺角蟬的梗節上均有毛形感器分布,考慮到毛形感器的功能,這可能意味著其在此兩種角蟬雄蟲搜尋雌蟲的過程中起重要作用。
與毛形感器不同,錐形感器是由一個到多個神經元支配的短毛(小釘)或錐體,表面多孔且樹突分支豐富,功能上主要起機械(接觸)和化學感器作用,與嗅覺刺激傳導密切相關,氣味結合蛋白在其中表達,保證其嗅覺受體的作用得以發揮[35]。另外,錐形感器對植物氣味刺激存在特殊的感受作用,對昆蟲在寄主的尋找與選擇等方面提供幫助[36]。兩種角蟬鞭節上僅觀測到錐形感器分布,且錐形感器均分布于鞭節膨大弧度較大的一側,這種特殊的分布方式可能有利于其功能的表達。
角蟬科與葉蟬科同屬角蟬總科,目前的分子系統發育研究證據也表明二者親緣關系較近,二者的觸角形態和感器類型與分布表現出相當的相似性,觸角感器可能具有作為這兩個類群間系統分類方面形態學依據的潛力。褐三刺角蟬和黑圓角蟬的觸角感器數量較少,類型單一,毛形感器均僅分布于觸角梗節,而錐形感器在二者觸角梗節和鞭節均有分布,且錐形感器僅分布于鞭節基部膨大弧度較大的一側,這種分布方式的成因及其特定功能暫不明確。為了探明這兩種角蟬觸角感器的內部結構、功能及其特異性分布的原因,為生物學防治和系統發育等相關研究提供參考,未來需要結合透射電鏡進一步觀察,并結合行為學、生理學以及分子生物學等學科,進行更加深入的觀察和探索。
