玉米BBX基因家族鑒定及表達分析
2022-07-15 06:27:08趙長江徐尚緣都夢翔宋巨奇徐晶宇楊克軍李佐同
西北農業學報 2022年6期
關鍵詞:分析
趙長江徐尚緣都夢翔宋巨奇徐晶宇楊克軍李佐同
(黑龍江八一農墾大學 農學院/農業農村部東北平原農業綠色低碳重點實驗室/黑龍江省現代農業栽培技術與作物種質改良重點實驗室/黑龍江省秸稈資源化利用工程技術研究中心/黑龍江省普通高校寒地作物種質改良與栽培重點實驗室,黑龍江大慶 163319)
鋅指(結構)蛋白作為一類重要的轉錄因子,能夠與核酸及蛋白質互作,參與多種生物學過程調控[1]。其中一個亞家族B-box(BBX)蛋白N 端至少含有一個B-box結構域,有的成員會多一個B-box或CCT(CONSTANS,CO-like and TOC1)結構域,在植物生長發育過程中發揮重要作用[2-3]。值得一提的是,CCT 結構域也是具有轉錄因子活性的基因家族[4-5]。擬南芥(Arabidopsis thaliana)CONSTANS(CO)/At BBX1 作為該家族的典型代表,已成為光周期途徑調控開花的主導者[6-8],同時還有At BBX4[9]、AtBBX6[10]、At-BBX7[11]和At BBX28[12]等成員參與開花調控。其中AtBBX28通過與AtBBX1互作,調控開花Flowering Locus T(FT)基因表達[12];當然BBX家族對開花的調控也存在不依賴AtBBX1的通路[13]。眾所周知,光周期對開花的調控歸根結底是對光信號的響應,包括避蔭反應[13]。在光敏色素和隱花色素依賴途徑中,依據與通路中兩標志因子 CONSTITUTIVE PHOTOMORPHOGENIC1(COP1)和ELONGATED HYPOCOTYL 5(HY5)的關系,將BBX 參與光形態建成信號通路大致分為4 種類型,即雙因子依賴、依賴COP1不依賴HY5、不依賴COP1 依賴HY5 和雙不依賴[2]。當然COP1 可以激活也可以抑制BBX,BBX 亦可激活也可抑制HY5,形成復雜的信號通路實現植物對光的形態建成調控。其中,At BBX24不僅參與可見光的形態建成,也參與UV-B光介導的形態建成[14-15]。近來發現,蘋果(Malus domestica)MdBBX37直接與光信號正向調節因子Md HY5的啟動子結合并抑制其表達,從而解除Md HY5介導的下胚軸抑制[16]。
植物BBX 除參與光生長發育調控,還參與對多種逆境和激素的應答[2,17]。例如,AtBBX18和AtBBX24分別參與對熱和鹽等非生物逆境響應[18-19]。Md BBX37調控茉莉酸介導的耐冷性,通過 JAZ (JAZ MONATE ZIM-DOMAIN)-BBX37-ICE1(inducer of CBF expression 1)-CBF(C-repeat binding factor)級聯通路提高耐冷性[20]。 菊 花 (Chrysanthemum morifolium)CmBBX19通過與ABF3 模塊通過ABA(脫落酸)依賴的途徑在菊花的耐旱性調控中發揮作用[21]。甜薯(Ipomoea batatas)IbBBX24可以通過調控茉莉酸合成和信號途徑增強植物對紋枯病的抗性并提高產量[22]。目前,已有研究對擬南芥[23]、水稻[24](Oryza sativa)、番茄[25](Solanum lycopersicum)、蘋果[26-27]、葡萄[28](Vitis vinifera)和矮牽牛[29](Petunia hybrida)等多個物種中BBX基因家族進行鑒定。然而關于高光效C4作物玉米(Zea mays)BBX 基因家族系統研究鮮有報道。本研究通過生物信息學方法鑒定出玉米BBX 所有成員,對家族基因特征、全生育期和非生物逆境下表達情況,并對生育期組織基因的共表達進行分析,為揭示玉米BBX 基因家族成員的生物學功能提供依據。
1 材料與方法
1.1 蛋白鑒定及系統進化分析
通過Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)下載玉米、擬南芥、水稻、二穗短柄草(Brachypodium distachyon)和番茄基因組數據,以擬南芥和水稻BBX 家族成員蛋白序列為模板,利用TBtools軟件獲取候選ZmBBX 蛋 白 序 列,并 上 傳 到 NCBI CD-Search、PFAM、SMART 進一步驗證。利用Expasy(https://web.expasy.org/protparam/)預測等電點、相對分子質量、氨基酸數量,利用PSORT 網站(http://psort1.hgc.jp/form.html)預測亞細胞定位。
利用MegaX(https://www.megasoftware.net/)進行ClustalW 多重對比,構建系統進化樹,生成算法為ML(Maximum Likelihood),校驗參數Bootsrtrap重復1 000次。
1.2 基因結構及其啟動子順式作用元件分析
利用TBtools繪制基因在染色體上的分布圖及外顯子-內含子結構圖;取翻譯起始點ATG 上游2 000 bp和1 000 bp啟動子序列提交Plant-CARE (http://bioinformatics.psb.ugent.be/webtools/)進行順式作用元件分析。利用MCScan X 進行物種間共線性分析。
1.3 蛋白結構及互作蛋白預測分析
利用 MEME (https://meme-suite.org/meme/tools/meme)分析蛋白保守基序,并利用TBtools繪制保守基序圖。使用STRING (https://string-db.org/)數據庫對蛋白互作關系進行預測。
1.4 逆境和全生育期組織表達譜
逆境(低溫、高溫、鹽、紫外)的表達數據來源于SRA 數據庫,登錄號為低溫(SRR1238717,SRR1819204,SRR1819205)、高溫(SRR1238715,SRR1819196,SRR1819198)、鹽(SRR1238719)和紫外(SRR1238720)。玉米全生育期的表達數據來源于Maize eFP Browser(http://bar.utoronto.ca/efp_maize/)數據庫。從NCBI-GEO 下載GSE50191轉錄組表達譜數據,提取其中的FPKM 值轉化為TPM 值作為基因表達譜數據。
1.5 組織表達數據的共表達分析
使用TBtools進行加權共表達網絡構建,以至少6成樣本中該基因的TPM 值>1為標準對基因表達矩陣進行篩選,構建無尺度網絡與劃分相關模塊,閾值為0.8軟閾值為β=22,并繪制模塊-性狀熱圖。利用TBtools 軟件進行GO 和KEGG 富集分析,以玉米基因組為參考數據庫,利用TBtools進行可視化,設置數量參數為30。
1.6 鹽脅迫表達分析
以玉米自交系合344為供試材料,1/2 Hoagland營養液培養至兩葉一心期進行處理。在營養液中添加150 mmol/L NaCl,分別于處理0 d、1 d、3 d和7 d取樣,將地上和地下部分離,用液氮冷凍存放于-80 ℃冰箱中保存。每組樣品3個生物學重復。采用TRIZOL 試劑提取玉米葉片和根組織總RNA,純化后構建c DNA 文庫,通過聚合酶鏈反應(PCR)富集cDNA 文庫,然后使用Illumina HiSeq 2000平臺對構建的文庫進行RNA-seq測序(SRP176441 和SRP307694)。抽取BBX基因構建鹽處理下根和葉表達譜,相對表達量計算方法為:首先將3次生物學重復處理的FPKM 平均值與對照(0 d)平均值相比,然后對二者比值進行log2FC轉換,轉換值即為相對表達量。
2 結果與分析
2.1 ZmBBX基因家族成員的鑒定與命名
獲得34個ZmBBX 基因家族成員,按照基因在染色體的位置依次命名為ZmBBX1-ZmBBX34(表1)。家族成員中蛋白氨基酸序列長度為142~498 aa,分子質量為15.38~53.77 ku;等電點為4.55~10.52。預測亞細胞定位發現,除ZmBBX5、ZmBBX19、ZmBBX26定 位 于 細 胞質,Zm BBX9、Zm BBX24、ZmBBX25、Zm BBX30和ZmBBX32定位于線粒體,ZmBBX33定位于內質網外,其余成員均定位于細胞核內。
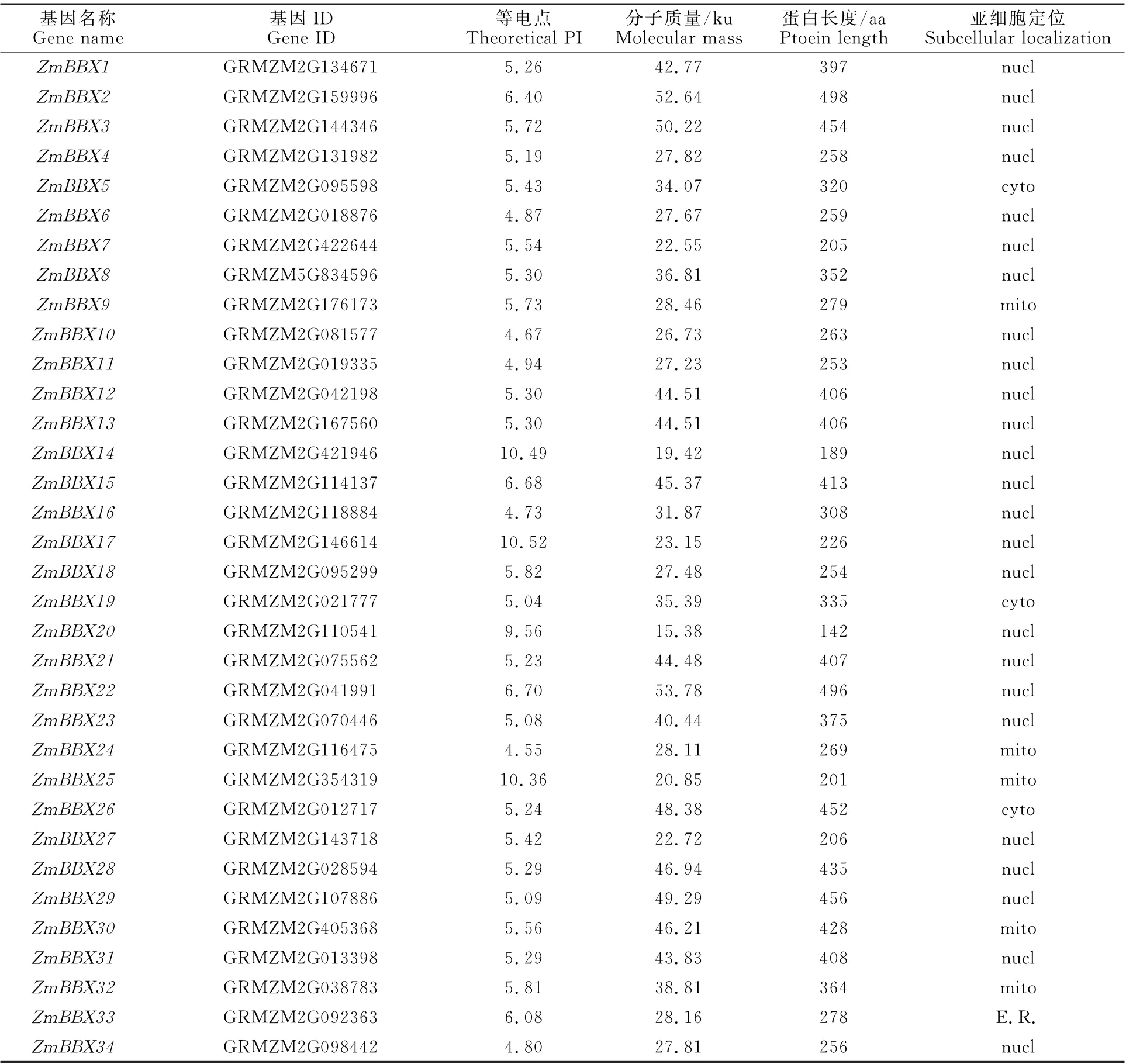
表1 ZmBBX 基因編碼蛋白序列理化性質Table 1 Physicochemical properties of protein sequence encoded by ZmBBX gene
2.2 ZmBBX家族成員蛋白進化分析
選取鑒定的34個玉米、32個擬南芥、30個水稻、22個二穗短柄草和29個番茄BBX 家族成員進行系統聚類分析,根據含有B-box 和CCT結構域的情況,ZmBBX家族可分為5個亞家族(Ⅰ~Ⅴ)(圖1),第Ⅰ亞族只含有2個B-box結構域,包括ZmBBX4、ZmBBX6、ZmBBX7、ZmBBX8、ZmBBX11、Zm BBX16、Zm BBX18、Zm BBX20、ZmBBX23、ZmBBX24、ZmBBX27、ZmBBX28和Zm BBX34等13個家族成員;第Ⅱ和Ⅴ亞族含有2個B-box結構域和1個CCT 結構域,但其B-box 在氨基酸序列上有差異,分別包括ZmBBX1、ZmBBX2、ZmBBX5、ZmBBX9、ZmBBX19、ZmBBX30、Zm BBX32和ZmBBX33等8個成員和Zm BBX12、ZmBBX13、Zm BBX21 和ZmBBX31等4個成員;第Ⅲ亞族含有1個B-box結構域和1個CCT 結構域,包括ZmBBX3、Zm BBX15、ZmBBX22、ZmBBX26 和ZmBBX29 等5個成員;第Ⅳ亞族只含有1個B-box結構域,包括ZmBBX10、ZmBBX14、ZmBBX17 和ZmBBX25等4個成員。各個亞家族中玉米與水稻、二穗短柄草聚成一支,表明雙子葉與單子葉植物的進化存在差異。

圖1 ZmBBX家族成員蛋白系統進化樹Fig.1 Protein phylogenetic tree of ZmBBX family member
2.3 ZmBBX家族成員基因進化分析
對玉米、水稻、二穗短柄草的BBX 家族基因進行共線性分析(圖2),34個Zm BBX基因不均勻地分布在10條染色體上,其中5號染色體上分布的成員最多,涉及Zm BBX15~Zm BBX21共8個,8號染色體上不存在BBX 家族成員。另外,18個ZmBBX同源基因出現在二穗短柄草和水稻染色體中,供試禾本科作物基因組BBX 具有較好的共線性,這與進化樹的結果一致。同時,對Zm BBX 家族基因進行串聯重復事件分析,共有11對重復基因,分別是ZmBBX3~Zm BBX15、ZmBBX4~Zm BBX20、Zm BBX7~Zm BBX27、ZmBBX11~ZmBBX18、ZmBBX12~Zm BBX21、ZmBBX14~Zm BBX17、ZmBBX16~ZmBBX24、Zm BBX17-Zm BBX25、Zm BBX23~ZmBBX28、ZmBBX34~Zm BBX4、Zm BBX34~ZmBBX20,其Ka/Ks 值 介 于0.187 856~0.570 824之間,二者比值小于1說明其受到純化選擇。
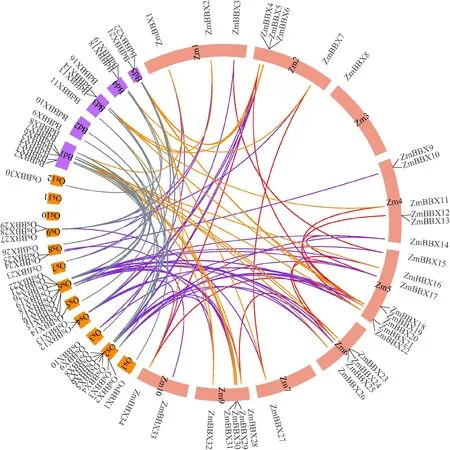
圖2 ZmBBX家族染色體定位(A)及二穗短柄草、玉米、水稻間共線性(B)Fig.2 Chromosome location of ZmBBX family(A)and collinearity analysis among Brachypodium distachyon,Zea mays and Oryza sativa(B)
2.4 ZmBBX家族基因結構及編碼蛋白保守結構分析
保守結構域分析發現成員含有2~8個保守基序(圖3-A、3-C),所有成員均含有B-box結構域核心序列(Motif 1)。其中ZmBBX4、Zm BBX5、ZmBBX6、Zm BBX7、Zm BBX8、Zm BBX11、ZmBBX12、ZmBBX13、ZmBBX16、ZmBBX18、Zm BBX19、Zm BBX20、Zm BBX21、Zm BBX23、Zm BBX24、ZmBBX27、ZmBBX28、ZmBBX31和Zm BBX3419 個 成 員 含 有 另 一 個B-box 保 守 域(Motif 3);ZmBBX1、ZmBBX2、ZmBBX3、ZmBBX5、ZmBBX9、ZmBBX12、ZmBBX13、ZmBBX15、ZmBBX19、ZmBBX21、ZmBBX22、ZmBBX26、ZmBBX29、ZmBBX30、Zm BBX31、ZmBBX32和ZmBBX3317 個 成 員 除 了 具有典型的B-box 結構域外,還含有CCT 結構域(Motif 3)。
對Zm BBX 家族成員的基因內含子和外顯子分析(圖3-B),第Ⅰ組成員內含子數量為1~5個,第Ⅱ組為0~3個,第Ⅲ組和Ⅳ均為3~4個,第Ⅴ組為0~2個。其中,Zm BBX9、ZmBBX14、ZmBBX17、ZmBBX25和ZmBBX33沒有內含子,ZmBBX28內含子數目最多為5個。

圖3 ZmBBX家族保守結構域(A和C)及基因結構(B)Fig.3 ZmBBX family conserved domain(A and C)and gene structure(B)
2.5 ZmBBX家族基因啟動子區順式作用元件分析
通過基因上游2 000 bp啟動子區元件分析發現,除啟動子核心元件TATA-box、CAAT-box以外,Zm BBX 家族基因啟動子還存在四大類作用元件(圖4-A),如光響應相關元件G-box、Sp1、GT1-motif等20 種,環境脅迫相關元件ARE、MBS等7種,以及ABRE、P-box等5種激素應答相關元件,同時還有生長發育相關元件CATbox、O2-site等5種。光響應、激素應答、環境脅迫相關元件均分布在所有Zm BBX 成員中,除ZmBBX1、ZmBBX23和ZmBBX27外其余家族成員均含有生長發育相關元件。ZmBBX家族基因上游1 000 bp啟動子區域部分元件的數量及位置統計如下(圖4-B),其中光響應元件(G-box)以及脫落酸應答元件(ABRE)數目較多分布廣泛,且在5′-3′鏈上數目較多;參與低溫響應的元件(LTR)在ZmBBX11上數目最多;其余元件(GT1-motif、TGA-element、CAT-box、O2-site、ARE)在部分成員中也有分布但數目較少。

圖4 ZmBBX家族基因啟動子區順式作用元件分布(A)及數目(B)Fig.4 Distribution(A)and number(B)of cis-acting elements in promoter of ZmBBX gene family
2.6 ZmBBX家族基因全生育期組織表達譜分析
ZmBBX 家族(除ZmBBX24)在玉米全生育期的基因表達模式分為3 大類,進一步細分為5小 類(圖5)。其 中,ZmBBX 7、ZmBBX 2 7和ZmBBX28等3個基因在玉米全生育期的表達量都很高并在表達模式中聚為第一大類;第二大類包括在全生育期具有表達但表達量較低的Zm BBX2、Zm BBX4、ZmBBX8、Zm BBX9、ZmBBX10、ZmBBX16、ZmBBX17、ZmBBX25、ZmBBX26、ZmBBX29、Zm BBX31、ZmBBX33和ZmBBX3413個基因在內的第一類和表現出較為明顯的組織特異性,在玉米葉片中表達量較高的ZmBBX3、ZmBBX5、Zm BBX14、ZmBBX15、Zm BBX19、ZmBBX20和ZmBBX227 個基因的第二類。第三大類包括3 個亞類,第一亞類涉及ZmBBX6、ZmBBX11、Zm BBX18和Zm BBX324 個 成 員,在玉米葉片、種子形成前期和第一莖節間等組織部位特異表達;第二亞類涉及ZmBBX1、Zm BBX12、ZmBBX13和ZmBBX234個成員,在葉片、苞葉、胚胎、種子、胚乳中表達量較高,且Zm BBX12和Zm BBX13在各組織中表達量完全一致;第三亞類涉及Zm BBX21和Zm BBX302 個成員,在全生育期均有表達但整體表達量較低。

圖5 ZmBBX家族基因全生育期表達分析Fig.5 Expression of ZmBBX family gene during whole growth period
2.7 生育期共表達的GO 和KEGG 富集分析
以GEO 數據庫的GSE50191轉錄組數據為基礎構建全生育期基因共表達網絡,通過對篩選的18 000個基因進行分析獲得8個共表達模塊(圖6-A、6-B)。在所有模塊中有19 個Zm BBX基因在列,其中MEbrown 和MEblue模塊中的數目最多,分別包括ZmBBX1、ZmBBX4、Zm BBX10、ZmBBX16、ZmBBX32和Zm BBX7、Zm BBX19、ZmBBX27、Zm BBX28、Zm BBX34,MEturquoise 中 包 括ZmBBX11、ZmBBX18、Zm BBX30和Zm BBX31,MEblack 中 包 括ZmBBX17和ZmBBX33,MEpink、MEred、MEmagenta中分別包括ZmBBX2、ZmBBX5和Zm BBX24。依據模塊與表型性狀的相關性,選擇有Zm BBX基因參與的且與GK 2相關性最高的MEbrown模塊進行GO 和KEGG 富集分析(圖6-C、6-D),其模塊與性狀的相關系數為0.9和P值為0.002。GO 富集分析發現,基因分子功能主要富集于蛋白質異二聚活性(GO:0046982)和磷酸甘油酸變位酶活性(GO:0004619),細胞組分富集于細胞質(GO:0005737)和微體(GO:0042579),生物過程富集于有機酸代謝過程(GO:0006082)、羧酸代謝過程(GO:0019752)和含氧酸代謝過程(GO:0043436)等。KEGG 分析發現,該模塊主要涉及代謝和碳水化合物代謝等代謝過程,內質網蛋白質加工的遺傳信息過程,過氧化物酶體等細胞過程,外泌體和脂質合成蛋白等蛋白功能。
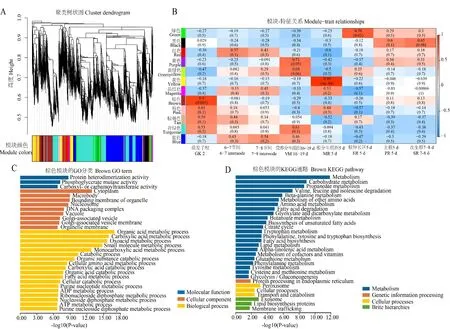
圖6 共表達網絡構建(A和B),關鍵模塊GO 功能富集分析(C)和KEGG 富集分析(D)Fig.6 Co-expression network construction(A and B),GO enrichment analysis of key modules(C)and KEGG enrichment analysis(D)
2.8 鹽處理下ZmBBX家族基因表達譜
Zm BBX 家族成員在NaCl處理玉米幼苗根和葉中表達不盡相同(圖7)。玉米幼苗葉片中差異表達(log2FC≥1)的ZmBBX基因共有17個,上調的有13個和下調的4個;其中log2FC≥2的有10個,上調的8個和下調的2個。同時,玉米幼苗根中差異表達(log2FC≥1)的Zm BBX基因共有19個,上調的有13 個和下調的6 個;其中log2FC≥2的有2個,均上調表達。
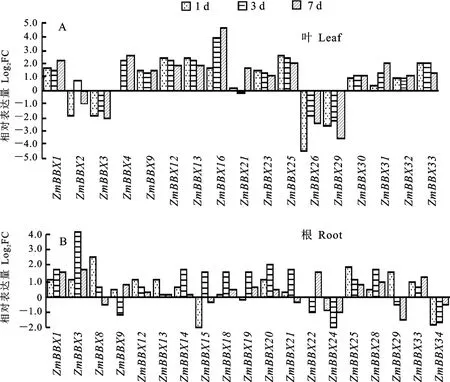
圖7 ZmBBX家族基因葉(A)和根(B)響應鹽脅迫的表達分析Fig.7 Expression analysis of ZmBBX family in leaves(A)and roots(B)under salt stress
地上和地下都差異表達(log2FC≥1)的ZmBBX基因有9個,包括ZmBBX1、ZmBBX3、ZmBBX9、Zm BBX12、ZmBBX13、Zm BBX21、ZmBBX25、ZmBBX29和Zm BBX33,其 中 除ZmBBX3在葉片中下調和根中上調,其余ZmBBX 在葉片和根中基本都上調表達。值得一提的是,差異表達倍數較大(log2FC≥4)的基因有3個,其中葉片中Zm BBX16上調表達和Zm BBX26下調表達,根中ZmBBX3明顯上調表達。
2.9 ZmBBX蛋白互作及逆境表達譜分析
利用STRING 數據庫預測BBX 的蛋白互作特性(圖8-A)。整個蛋白互作網絡有23個節點,節點之間共存在45 組蛋白互作關系。其中Zm BBX14、Zm BBX17 和ZmBBX25 位 于 整 個BBX 基因家族蛋白互作網絡的中心,分別與Zm BBX2、Zm BBX4、ZmBBX7、ZmBBX16、Zm BBX20、ZmBBX24、ZmBBX27 和ZmBBX34 這8個家族成員互作,還發現其他家族蛋白MYBR20、glk45、GRMZM2G181030 都 與 Zm BBX12、ZmBBX13、ZmBBX21 和ZmBBX31 有 互作關系,且glk45還與ZmBBX19和Zm BBX30存在互作關系。結合逆境基因共表達分析(圖8-B)ZmBBX21與GRMZM2G181030、MYBR20表 達模式相似,都在低溫脅迫下高表達,其中MYBR20在低溫下表達量大幅上調,在轉錄水平上支持蛋白互作。
3 討論
植物轉錄因子通過其特有結構域與DNA 及其他蛋白相互作用,激活或抑制基因的表達[30],其中植物BBX 轉錄因子在調節多種生命活動中發揮著重要的作用[2]。本研究利用生物信息學方法鑒定出玉米34個Zm BBX,依據蛋白進化該家族可以劃分為5個亞族。其中,劃歸第Ⅰ亞族的8個(AtBBX18~At BBX25)擬南芥家族成員均已報道參與光(暗)的形態建成,且AtBBX22、At-BBX24和At BBX25均可以與COP1和HY5 互作[2],推測同組Zm BBX4、ZmBBX6、ZmBBX7、ZmBBX8、ZmBBX11、ZmBBX16、ZmBBX18、ZmBBX20、ZmBBX23、ZmBBX24、ZmBBX27、ZmBBX28和ZmBBX34這13 基 因 可 能 也 參 與玉米對光形態建成的調控。第Ⅱ亞族中包括6個(AtBBX1~AtBBX6)擬南芥家族成員,該亞族基因涉及開花調控、氣孔開放、逆境響應等生理作用[2],推測同組玉米ZmBBX1、Zm BBX2、Zm BBX5、ZmBBX9、ZmBBX19、ZmBBX30、ZmBBX32和Zm BBX33這8個基因也具有類似功能,其中ZmBBX30與OsBBX18/Hd1[31]直系同源,參與花期調控,值得一提的是,本組Zm BBX19/Zm-COL3已被鑒定在玉米花期調控中發揮重要作用[32]。第Ⅴ亞族中包括9 個擬南芥AtBBX 成員,其中AtBBX7、AtBBX9、AtBBX11和AtBBX13主要參與對生物逆境和非生物逆境的響應[2],推 測 同 組Zm BBX12、Zm BBX13、Zm BBX21和ZmBBX31這4 個 基 因 參 與 玉 米 對 逆 境的應答,而且本研究也發現上述4個基因均可以被鹽處理誘導表達,其中Zm BBX12、ZmBBX13和ZmBBX21具有相同組織表達模式。此外,與ZmBBX31聚于一個分支的OsBBX7/DTH2為水稻抽穗調控基因[33],推測該基因還參與植物生長發育調控。
本研究中對鹽脅迫下玉米幼苗ZmBBX 家族基因的表達進行分析,發現有24 個Zm BBX基因參與玉米幼苗對鹽脅迫的應答,包括地上和地下都差異表達的9 個Zm BBX基因,分別為ZmBBX1、ZmBBX3、ZmBBX9、ZmBBX12、ZmBBX13、Zm BBX21、ZmBBX25、ZmBBX29和Zm BBX33,其中根部ZmBBX3明顯上調表達。此外葉片ZmBBX16上調和Zm BBX26下調亦十分明顯。另一方面,從全生育期組織表達結果來看ZmBBX7、Zm BBX27和Zm BBX28在玉米全生育期的表達量都很高,ZmBBX21和ZmBBX30在全生育期均有表達但整體表達量較低,且部分基因具有較明顯的組織表達特異性。上述基因的表達支持了本研究Zm BBX基因啟動子存在大量光、生長發育、環境脅迫以及激素應答等相關順式作用元件的結果。進一步通過全生育期共表達分析 發 現,ZmBBX1、ZmBBX4、ZmBBX10、ZmBBX16和ZmBBX32等基因可能參與光響應的萌發籽粒的代謝和生長發育[34],而且可能通過蛋白二聚化分子功能,調控細胞質和過氧化物酶體中糖和酸代謝等生物學功能來實現。
通過互作蛋白預測分析發現,同屬第Ⅴ類的ZmBBX12、ZmBBX13、ZmBBX21 和ZmBBX31等BBX 蛋白與已知的生長發育調控轉錄因子家族成員MYBR20[35]和glk45[36]構成一個互作組;同時,多種逆境表達譜分析也發現ZmBBX21和MYBR20共聚一類,該共表達模式也支持了蛋白互作的預測結果。此外,Zm BBX14、ZmBBX17和ZmBBX25居于BBX 家族同源互作的核心,多種逆境表達模式下Zm BBX14和ZmBBX17相近,ZmBBX17和ZmBBX25全生育期組織表達模式相近,不僅支持蛋白互作預測,而且表明ZmBBX 同源互作可能具有分工,用于協調逆境與生長發育。
4 結論
本研究在玉米基因組水平共鑒定出BBX 基因家族成員34個,Zm BBX 家族基因生育期組織表達和逆境表達分化明顯,在鹽等非生物逆境和光相關的生長發育調控中發揮重要作用。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
