花魔芋響應軟腐病菌侵染初期的轉錄組分析
2022-07-15 06:27:02斐馮明月王菁軒邵永春王甜王瑩盧謝敏方姝穎
西北農業學報 2022年6期
何 斐馮明月王菁軒邵永春王 甜王 瑩盧謝敏方姝穎
(安康學院 現代農業與生物科技學院,陜西安康 725000)
魔芋系天南星科魔芋屬(Amorphophallus)多年生草本植物,它是目前為止自然界唯一能大量提供葡甘聚糖的重要經濟作物[1]。這種天然高分子優質膳食纖維-葡甘聚糖,具有凝膠性、增稠性、持水性和成膜性等獨特的特性,被廣泛應用于生物醫藥、食品和化工等領域[2]。由于其較高的經濟價值,近年來魔芋已發展成為云南、貴州、四川、陜南及湖北等山區農民脫貧致富的支柱產業[3]。花魔芋是中國的主栽品種,其產量大、葡甘聚糖含量高,但抗病性較差[4]。其中,土傳和種傳細菌性軟腐病是花魔芋生產過程中危害最大且極難防控的病害,在植株生長期和采后貯藏期均可發病,可危害花魔芋葉片、葉柄及地下塊莖,輕則造成減產20%~30%,重則導致80%以上減產甚至絕收[4],嚴重制約魔芋產業的可持續發展。隨著科技的不斷進步,利用抗病基因進行分子育種已成為解決作物抗病性的有效途徑,這一途徑的關鍵是對花魔芋新型內源抗病基因的挖掘和篩選以及進一步對其抗病防御機制的解析。
目前,對花魔芋軟腐病相關的研究雖然已有一定報道,但是主要集中在病原菌鑒定與檢測[3]、病害防治[5]及傳統的抗病品種選育[6]等方面。Wu等[7]研究發現,芽孢桿菌產生的酰基高絲氨酸內酯酶(Aii A)通過干擾其群體感應系統的信號分子N-酰基高絲氨酸內酯(AHL),從而對花魔芋軟腐病菌產生抑制作用。雷珍珍等[8]從抗軟腐病花魔芋植株基因組中分離獲得一條NBSLRR 類抗病基因同源序列。上述結果為花魔芋對軟腐病菌侵染響應分子機制的研究提供了理論基礎,但這些研究僅局限于病原菌群體感應的信號分子或花魔芋中部分抗病基因的分析,而植物對病原菌侵染的抗性反應涉及多種途徑和基因的表達調控,有必要系統鑒定花魔芋中參與軟腐病脅迫響應的多種基因,以揭示花魔芋抗病的分子機制及挖掘重要的抗病基因與調控功能。
植物染病后,基因轉錄和翻譯水平通常發生明顯改變,而基因表達水平的變化則早于病癥的出現。李玲[9]在研究2個稻瘟病菌生理小種侵染時,發現在3個水稻品種上接種不同病菌后24 h和48 h分別有12 456個和4 851個基因發生差異表達,說明在染病早期草本植物的基因表達已經明顯改變。此外,軟腐病對花魔芋造成的危害嚴重且不可逆,增強植株自身染病初期的抗性十分必要。轉錄組測序技術(RNA-seq)作為一種高效快捷、準確度高、成本低的研究手段,被廣泛應用于模式植物擬南芥[10]和玉米[11]、非模式植物甘蔗[12]和花生[13]等多種植物抗病機理研究。本研究通過轉錄組測序技術探究花魔芋感染軟腐病菌初期基因表達的整體變化情況,通過對差異表達基因進行GO 功能分類和KEGG 富集性分析,篩選花魔芋響應細菌性軟腐病菌侵染的相關基因、抗病相關的信號傳導途徑和生物活性物代謝通路,以期為深入研究魔芋抗病分子機制提供理論參考,也為魔芋抗病分子育種提供候選基因。
1 材料與方法
1.1 材料
植物材料為1 a生的安康市本地花魔芋盆栽苗。花魔芋軟腐病菌菊果膠桿菌(Pectobacterium chrysanthemi)CZS-B6菌株由中國典型培養物保藏中心(China Center for Type Culture Collection,簡稱CCTCC,也稱武漢大學保藏中心)提供。
1.2 方法
1.2.1 病原菌接種與取樣 花魔芋軟腐病菌的培養和接種體的制備參考徐煒等[14]的方法。參照何斐等[15]的方法進行盆栽花魔芋的種植與管理,于盆栽花魔芋生長至苗高約10 cm 左右,采用澆灌法[14]將稀釋至108CFU/m L 的接種體菌懸液澆灌到健康的盆栽花魔芋植株根際,每株澆灌200 m L,以澆灌無菌水的花魔芋作為對照。根據預試驗效果,分別選取澆灌處理第5天的健康和軟腐病菌侵染初期的花魔芋球莖,于液氮中迅速冷凍后至-80℃保存。每6個生物學重復當作1個處理,每處理重復2次。
1.2.2 RNA 提取、cDNA 文庫構建和轉錄組測序 將凍存樣品在液氮中研磨,用RNApure Plant Kit(含DNaseⅠ)試劑盒(康為世紀生物科技有限公司,北京)提取總RNA,分別采用Nanodrop 2000 和Agilent 2100 生物分析儀(Agilent Technology,美國)檢測RNA 樣品的濃度和質量。經檢測合格后送至北京組學生物科技有限公司構建cDNA 文庫,采用實時定量PCR 方法庫檢合格后,使用Illumina HiSeq 2500 測序平臺(Illumina Inc.,美國)進行轉錄組測序(RNAseq)。
1.2.3 測序數據處理 花魔芋無參考基因組,采用Trinity軟件[16]對原始測序數據(Raw data)過濾后獲得高質量數據(Clean data),對其進行組裝拼接成轉錄本。去冗余后獲得unigenes,使用COG、GO[17]、KEGG[18]、KOG[19]、Pfam[20]、Swiss-Prot[21]和Nr數據庫進行基因功能注釋。
差異表達基因篩選:用FPKM 值(每千個堿基的轉錄每百萬映射讀取的片段數)[22]計算基因表達量。差異表達基因篩選標準:花魔芋健康株與軟腐病株基因表達水平差異倍數FC(Fold change)≥2,且錯誤發現率FDR(False discovery rate)<0.01[23]。
差異表達基因GO 注釋和KEGG 富集分析:利用Blast2GO(https://www.blast2go.org/)進行差異表達基因的GO 注釋[24]。通過P值(P<0.05)的Benjamini-Hochberg矯正方法確定差異表達基因中顯著富集的GO 分類,使用Cytoscape(http://www.cytoscape.org/)注釋通路。利用KEGG 數據庫(http://www.genome.jp /kegg/),以P<0.05且Q≤0.05作為顯著富集標準,進行差異表達基因的通路富集分析[25]。
2 結果與分析
2.1 測序數據質量分析
利用Illumina測序平臺對健康和軟腐病菌侵染初期的花魔芋球莖轉錄組進行測序,經過對測序原始數據質量控制,在健康和發病魔芋中分別獲得19 236 352和21 116 175條clean reads(NCBI SRA 數據庫登錄號:PRJNA738708,各樣本序列登錄號 SAMN19759221 - SAMN 19759224),GC 含量分別為51.32%和51.36%,Q30值均高于93%。各樣品的clean reads與拼接得到的轉錄本參考序列的比對效率達到75%以上,數據顯示轉錄組測序質量良好,可進行下一步分析(表1)。

表1 花魔芋球莖轉錄組測序數據Table 1 Transcriptome data assembly for corm of A.konjac
2.2 Unigene功能注釋
將unigene序列與COG、GO、KEGG、KOG、Pfam、Swissprot和Nr數據庫比對和注釋,共獲得57 525條有注釋信息的unigenes,占全部unigenes的51.6%。其中COG 數據庫注釋信息最少,有18 357條;Nr數據庫注釋信息最多,高達53 611條,占總注釋unigenes的93.2%(表2)。
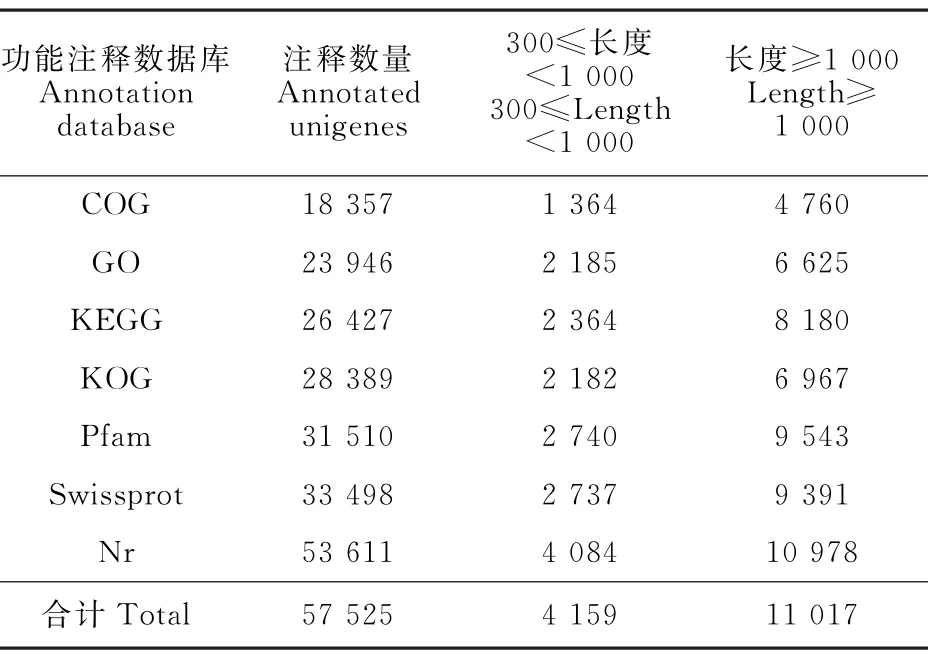
表2 Unigene注釋分析Table 2 Unigene annotation based on different databases
2.3 差異表達基因分析
轉錄組分析(圖1)表明,軟腐病菌侵染初期的花魔芋球莖中共出現差異表達基因3 222個。與健株相比,病株中有2 660 個基因表達上調,562個基因表達下調,上調基因比下調表達的基因多373.3%。差異基因表達量上調、下調最大的前10個基因中(表3),多數基因編碼特定功能的蛋白質或酶,如上調表達基因TRINITY_DN674_c0_g3 編碼Mi AMP1 抗菌蛋白,下調表達基因TRINITY_DN3252_c0_g3 編碼熱激蛋白,TRINITY_DN11231_c0_g1編碼胰蛋白酶與蛋白酶抑制劑、胰凝乳蛋白酶抑制劑。研究表明這些蛋白質、酶與抵御病菌侵染密切相關[26-27]。

表3 花魔芋在軟腐病菌脅迫下表達量上升和下降最大的前10個基因Table 3 Top 10 up-and down-regulated genes differentially expressed in A.konjac under P.chrysanthemi stress

圖1 軟腐病菌侵染初期花魔芋球莖中的差異表達基因分析Fig.1 Differential expression analysis of genes in corm of A.konjac with early response to P.chrysanthemi infection
2.4 差異表達基因功能分類和富集分析
對差異表達基因進行GO 功能分類(圖2),共分為3大類53個功能組。在生物學過程、細胞組分和分子功能3大類中,2個差異表達最多的功能組分別是新陳代謝過程(706個差異表達基因)和細胞進程(620個)、細胞組分(627個)和細胞(626個)、催化活性(644個)和結合(589個)。

圖2 基于轉錄組測序數據的差異表達基因GO 功能分類Fig.2 GO functional classifications of differentially expressed genes(DEGs)based on transcriptome sequencing data
通過KEGG 對花魔芋各通路進行注釋分析(圖3),發現1 148個差異表達基因在細胞過程、環境信息處理、基因信息處理、代謝和有機系統5大類的信號轉導及植物-病原菌互作等通路中富集。在29 條通路中,與代謝有關的通路最多(17條),占比58.6%。

圖3 差異表達基因的KEGG 通路分析Fig.3 KEGG pathway enrichment analysis of differentially expressed genes
在顯著富集的7條通路(表4)中,有3條與生物合成相關,2條與代謝有關,剩余的2條分別是糖酵解/糖異生和氧化磷酸化通路。富集程度最大的2 條通路分別是類黃酮生物合成[ko00941]和苯丙烷生物合成通路[ko00940]。在對花魔芋健株與軟腐病株差異表達基因分類和途徑分析的基礎上,推測軟腐病菌侵染花魔芋誘導的代謝途徑中的差異表達可能與苯丙烷和類黃酮生物合成等抗性有關,對防御病菌侵染發揮重要作用[28]。

表4 差異表達基因中顯著富集的通路Table 4 Significantly enriched pathways of differentially expressed genes(DEGs)
2.5 趨勢表達基因通路分析
為解析花魔芋抵御軟腐病菌侵染的關鍵通路和基因,逐條分析差異表達基因映射在KEGG 數據庫中的參考通路,結果發現一些通路的差異基因存在特定的表達變化趨勢(表5)。其中,硒化合物代謝和維生素B6代謝通路中,所有差異基因相比健株均呈現表達量上升的趨勢。在氨基糖和核苷酸糖代謝,甘氨酸、絲氨酸和蘇氨酸代謝,類黃酮生物合成等14條通路中,60%以上的差異基因相比健株表達量上調。但甾類激素生物合成(57.14%)和類胡蘿卜素生物合成(66.67%)通路中的大部分差異表達基因相比健株呈現下調趨勢。
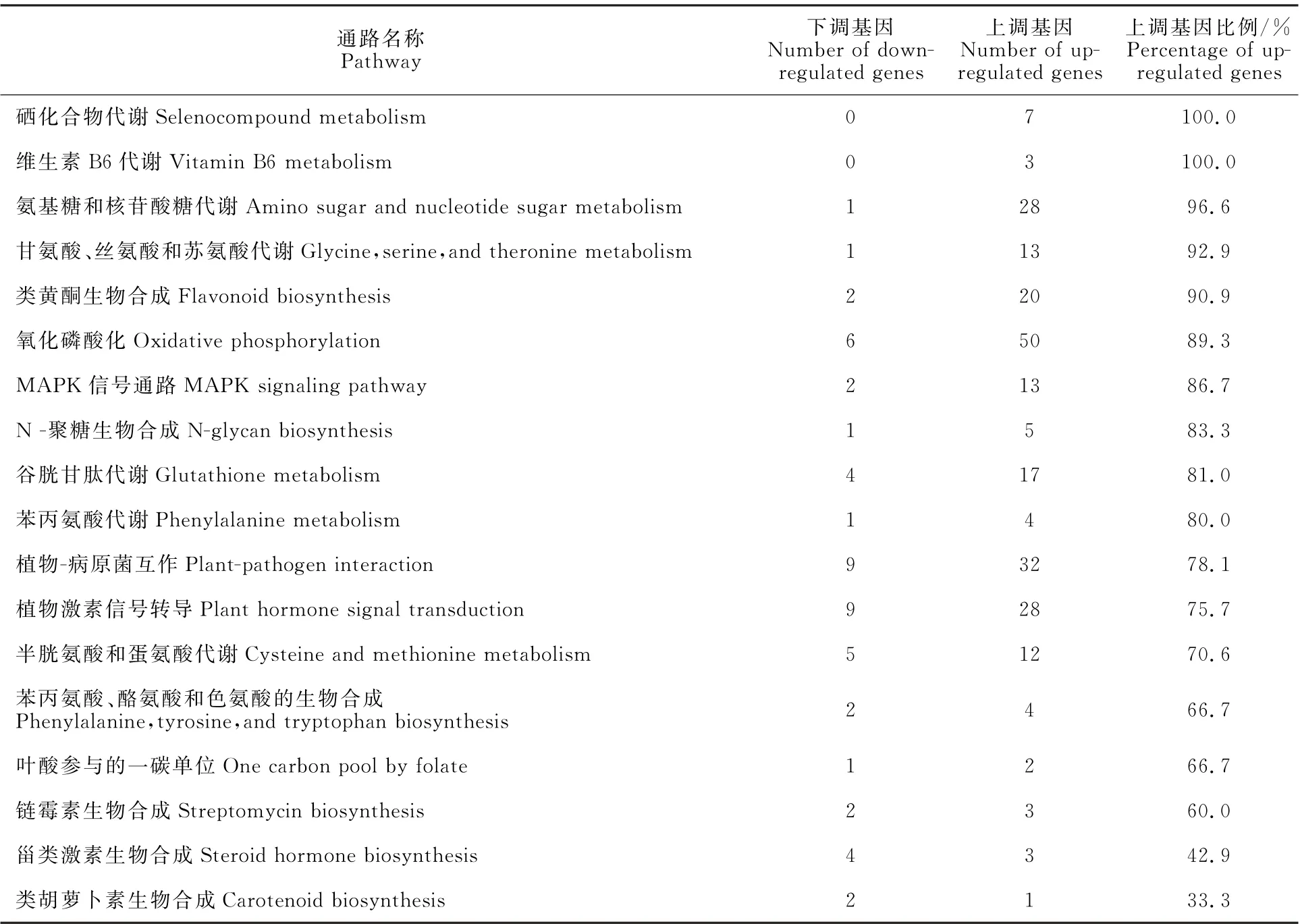
表5 趨勢表達基因KEGG 通路富集分析Table 5 Enrichment analysis of down-and up-regulated genes based on KEGG database
3 討論與結論
本研究對健康和軟腐病菌侵染初期的花魔芋球莖進行轉錄組測序分析,獲得111 404條unigenes和3 222個差異表達基因,其中2 660個基因(82.6%)上調表達,562個基因(17.4%)下調表達(病株/健株)。劉夢洋[29]研究發現,大白菜抗軟腐病突變體受軟腐病菌侵染初期12 h 時,412 個基因(81.9%)表達量上調,91 個基因(18.1%)表達量下調。于秋佟[30]通過轉錄組測序發現,軟腐病菌侵染24 h的擬南芥有579個基因(89.0%)表達量上調,72個基因(11.1%)表達量下調。這些研究與本結果類似,即上調表達基因數量明顯多于下調表達基因的數量,說明更多的基因被誘導表達來抵御軟腐病菌的入侵和發展。本試驗結果為后續鑒定花魔芋抵御軟腐病菌侵染初期發揮關鍵作用的基因縮小了篩選范圍,也為魔芋抗病育種奠定基礎。
在分析軟腐病菌脅迫下花魔芋中差異表達變化較大的基因時(表3),鑒定到部分響應軟腐病菌脅迫的差異表達基因。例如,在上調表達的基因中TRINITY_DN674_c0_g3編碼MiAMP1抗菌蛋白,已有研究表明其與植物抗病有關,Mi-AMP1抗菌蛋白重組畢赤酵母能誘導梨對擴展青霉產生抗性[31]。TRINITY_DN 1022_c0_g3編碼類枯草桿菌蛋白酶家族,有研究顯示類枯草桿菌蛋白酶在病原物侵染后被特異性誘導并參與宿主植物細胞程序性死亡,同時在病原物識別和信號轉導級聯反應中發揮重要作用[32]。值得關注的是,筆者發現也存在與抗病相關的下調表達基因,如TRINITY_DN3252_c0_g3編碼熱激蛋白,已有報道顯示眾多熱激蛋白參與調控植物抗逆和防衛反應[33]。TRINITY_DN11231_c0_g1 編碼胰蛋白酶和蛋白酶抑制劑,多種植物的蛋白酶抑制劑具有抗病蟲功能[34]。據此,推測花魔芋可能根據病菌脅迫的特點,通過上調表達具有針對性的抗性基因,同時下調表達廣譜抗性基因來響應軟腐病菌的脅迫。
本研究在分析KEGG 代謝通路時,發現花魔芋中16條通路的差異表達基因在軟腐病菌脅迫下全部或大部分上調表達(表5)。經逐條分析,可按照功能將其分成兩類:第1類與宿主植物抗逆有關;第2類與細胞合成原料相關。在第1類通路中,硒化合物和維生素B6代謝通路中差異表達基因全部上調。已有研究證明,硒是植物生長發育不可缺少的微量元素,其通過維持植物體內較高的抗氧化酶活性來誘導宿主產生抗病性[35];維生素B6作為多種代謝酶的重要輔助因子,參與植物氨基酸、乙烯合成等多種基本代謝活動,亦能通過調整細胞內抗氧化能力從而調控番茄植株對灰霉病的抗性[36]。黃酮類化合物是植物自身抗病、抗逆的主要成分[37],在本研究中,涉及黃酮類化合物合成的類黃酮生物合成通路中有91.0%差異基因上調表達。N-聚糖生物合成通路合成的寡糖是一類植物抗逆誘導劑[38],其中木寡糖能誘導擬南芥對半活體病原細菌丁香假單胞菌番茄致病變種、活體病原煙草花葉病毒和死體病原真菌核盤菌產生抗性[39]。在本研究中N-聚糖生物合成通路83.3%差異基因上調表達。而鏈霉素生物合成通路合成的鏈霉素主要用于防治由革蘭氏陰性細菌引起的煙草野火病和青枯病、大白菜軟腐病、柑橘潰瘍病、水稻白葉枯病和條斑病等植物病害[40],在本研究中鏈霉素生物合成通路60%差異基因上調表達,推測該通路合成的鏈霉素可能在抵御軟腐病菌侵入過程中發揮重要作用。第1大類參與宿主植物抗逆相關的信號轉導和物質運輸的主要通路還包括氧化磷酸化、MAPK 信號通路、氨基糖和核苷酸糖代謝通路、植物-病原菌互作和植物激素信號轉導[41],這些通路75%以上的差異基因相比于健株表達量上調。
在第2類通路中,半胱氨酸和蛋氨酸代謝,甘氨酸、絲氨酸和蘇氨酸代謝,谷胱甘肽代謝,苯丙氨酸代謝,苯丙氨酸、酪氨酸和色氨酸的生物合成均與氨基酸的代謝或合成相關,這些通路均有超過66.7%的差異基因受誘導上調表達,這可能引起相應的氨基酸合成加快,從而為合成抗病相關的蛋白和酶提供充足原料[42]。然而甾類激素和類胡蘿卜素生物合成通路的大部分差異基因表達下調(表5)。廣泛存在于植物體內的類胡蘿卜素和甾類激素主要參與調控植物生長發育及生理功能[43],且類胡蘿卜素含量也與植物抗病性相關[44]。類胡蘿卜素和甾類激素生物合成通路中基因表達的下調可能引起細胞內甾類激素和類胡蘿卜素供應緊缺,從而影響細胞包括抗軟腐病相關基因表達等在內的正常生理功能,這可能是染病花魔芋無法有效抵御軟腐病菌入侵的原因之一。
本研究通過RNA-seq測序得到3222個在花魔芋感染細菌性軟腐病初期出現差異表達的基因,篩選出與抗病相關且表達量差異大的基因。進一步進行GO 注釋和KEGG 通路分析發現2條通路的富集程度最大且合成物質具有抗病性-苯丙烷生物合成和類黃酮生物合成通路。通過這2條通路合成的物質可能在花魔芋抵御軟腐病菌侵入過程中發揮關鍵作用。同時,研究發現16條通路中60%以上的差異基因上調表達,而2條通路中大部分差異基因下調表達。其中,硒化合物代謝、維生素B6代謝、類黃酮生物合成、N-聚糖生物合成及鏈霉素生物合成這5條通路生成的相關物質與花魔芋抗軟腐病菌脅迫密切相關。這些抗病相關通路和差異表達基因的分析結果為花魔芋抗病機制的研究奠定了基礎,也為抗病相關基因的功能研究提供一定參考。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
