獼猴桃AcNPR1 基因克隆及抗病響應表達分析
2022-07-15 06:26:38東燕飛劉欣瑞彭雪張羽秦公偉
西北農業學報 2022年6期
關鍵詞:植物
曲 東燕 飛劉欣瑞彭 雪張 羽秦公偉
(1.陜西理工大學 生物科學與工程學院,陜西漢中 723000;2.陜西理工大學 陜西省資源生物重點實驗室,果樹資源保護與開發研究所,陜西漢中 723001)
獼猴桃為獼猴桃科(Actinidiaceae)獼猴桃屬(ActinidiaLindl.)木質藤本果樹,其果實富含膳食纖維、維生素C 等益于人體健康的功能營養成分,且經濟價值較高,具有良好的市場發展前景[1]。目前,我國獼猴桃的栽培種植面積和產量越居世界第一。然而由丁香假單胞菌獼猴桃致病變種(Pseudomonas syringaepv.actinidiae,Psa)引起的獼猴桃潰瘍病正嚴重影響著中國及世界各國獼猴桃產業的發展[2],該病害傳播速度快、傳播途徑多樣、防治難度極大,病原菌主要通過危害獼猴桃樹干、枝條、葉片及花的生長發育,導致獼猴桃果實大量減產甚至毀園,因此激活植物的固有防御系統提高獼猴桃抗病能力或選育獼猴桃抗病品種是降低獼猴桃潰瘍病危害的有效辦法。目前有關獼猴桃抗病機理的研究報道較少,因此挖掘獼猴桃抗病相關的關鍵基因,對于了解獼猴桃的抗病機制具有重要作用,對于獼猴桃抗病育種的工作具有一定的現實意義。
病程相關基因非表達子1(Nonexpressor of Pathogenesis-Related genes,NPR1)隸屬于NPR1基因家族[3],是水楊酸(Salicylic acid,SA)介導的植物系統獲得性抗性(Systemic acquired resistance,SAR)以及茉莉酸(Jasmonic jcid,JA)和乙烯(Ethylene,ET)介導的植物誘導獲得抗性(Induced systemic resistance,ISR)的一個主要調控因子,是植物多種抗病性信號傳導途徑的交叉點[4]。當植物受到病原菌侵染時,NPR1 特異性結構域能夠與WRKY 等轉錄因子結合激活植物免疫系統,作用于相關抗病基因,使基因表達水平上調,從而增強植物抗病能力[5]。NPR1基因包含一個N-末端BTB/POZ結構域、一個中心錨蛋白重復結構域ANKs(Ankyrin repeats)和一個C-末端反式激活結構域,其中BTB-POZ 結構域參與NPR1蛋白的同源二聚的形成[4,6-7],而ANKs與冗余的TGACG 序列元件結合轉錄因子TGA2、TGA5和TGA6相互作用共同激活植物抗性基因的表達,從而提高植物抗病能力[8-10]。NPR1基因及其同源基因已經在擬南芥(Arabidopsis thaliana)[11-12]、水稻(Oryza sativa)[13-14]、蘋果(Malus domestica)[15]、葡萄(Vitis vinifera)[16-17]等多種植物中被分離克隆,過表達At-NPR1基因的水稻中發現當水稻感染白葉枯病、稻瘟病時,At NPR1基因表達量明顯提高[18];過表達At NPR1基因的小麥提高了其對鐮刀菌赤霉病的抗病能力[12],在水稻中過表達Os NPR1基因后,不僅提高了水稻白葉枯病抗病性[14],而且提高了水稻對草食動物的敏感性。在雙子葉植物中也發現,過表達At NPR1基因的柑橘加強了柑橘對黃龍病、潰瘍病的耐受[19],在蘋果中過表達Malus NPR1基因,蘋果中PR 基因被激活,提高了蘋果樹對細菌和真菌病原菌的抗性[15],NPR1基因參與了植物免疫系統的調節。
越來越多的研究證據表明NPR1是SA 的受體[20]。在擬南芥中NPR1基因是SA 誘導PR(Pathogenesis-Related,PR)基因表達和抵抗病原體所必需的[4]。當植物未感染病原菌時,SA 水平較低,NPR1主要存在于通過分子間二硫鍵形成的低聚物中;當植物感染病原菌后,SA 水平升高,NPR1低聚物被還原為單體以響應病原體感染期間SA 積累增加引起的氧化還原變化[21]。
鑒于NPR1蛋白在植物防御反應中發揮重要的作用,而獼猴桃NPR1基因在抗病中的功能研究目前報道較少,故本研究選用高感病品種‘紅陽’和抗病品種‘徐香’獼猴桃為材料[22],克隆Ac NPR1基因序列,利用生物信息學方法對Ac-NPR1蛋白的理化性質,二、三級結構、亞細胞定位等特性進行預測分析,用DNAMAN 和MEGA 7.0 進行氨基酸多序列比對和進化關系的分析與構建。利用實時熒光定量PCR 分析獼猴桃Ac NPR1基因組織表達情況,以及獼猴桃潰瘍病致變種(Psa)、水楊酸加病原菌(SA+Psa)處理不同時間基因的表達量變化,旨在為初步明確獼猴桃Ac NPR1基因在抗病中的作用機制提供初步的理論基礎。
1 材料與方法
1.1 植物材料與處理
試驗材料源自陜西省漢中市城固縣自然好獼猴桃專業合作社獼猴桃園,移栽于陜西理工大學資源生物重點實驗室的嫁接2 a生高感病品種‘紅陽’獼猴桃(Actinidia chinensisvar.chinensis‘Hongyang’)、抗病品種‘徐香’(Actinidia deliciosavar.‘Xuxiang’)獼猴桃幼苗,試驗于2021年3月進行。
分別取‘紅陽’獼猴桃和‘徐香’獼猴桃的莖、花、葉片,裝入無菌的樣品管內標記,放入液氮速凍,并保存于-80 ℃冰箱備用。將‘紅陽’獼猴桃和‘徐香’獼猴桃幼苗分為3組,噴灑水為第1組(對照組,CK)、先水對葉片進行噴灑,1周后用丁香假單胞桿菌獼猴桃致變種(Pseudomonas syringaepv.actinidiae,Psa)菌液(陜西理工大學資源生物重點實驗室分離鑒定保存菌種,Genbank 序列登錄號 MW404385,菌液濃度108cfu/m L)進行噴灑處理為第2組(Psa)、先用水楊酸(2.5 mmol/L)對葉片進行噴灑預處理,1周后用丁香假單胞獼猴桃致變種液噴灑葉片為第3組(SA+Psa),預處理方法參照文獻[23];處理后分別在0 h、6 h、12 h、24 h、48 h、72 h 5個時間點取樣,裝入無菌的樣品管內,用記號筆標記采取樣品的處理和時間,立即放入液氮中進行速凍,轉入-80 ℃冰箱保存待用,每個樣品重復3 次。
1.2 RNA提取及cDNA合成
采用TransZol Plant 試劑(TransGen Biotech)提取不同處理時期樣品RNA,經電泳檢測(1%瓊脂糖凝膠)合格,Thermo NanoDrop 2000分光光度計測定各樣品的濃度以及OD260/OD280、OD260/OD230值;后 反 轉 錄 為cDNA(TransScript One-Step gDNA Removal and cDNA Synthesis Supermix,TransGen Biotech)并保存于-20 ℃冰箱備用。
1.3 獼猴桃Ac NPR1 基因的克隆及測序
根據獼猴桃數據庫(http://kiwifruitgenome.org/)中注釋為Regulatory protein NPR1蛋白基因的序列(編號Acc15406),利用Primer 6.0軟件設計引物,Ac NPR1基因上下游引物見表1。分別為ATGGCAAATTCTGC 和TTACGACAATTTTCTAATCTTGTAATTC。以Psa菌液處理12 h的獼猴桃葉片cDNA 為模板,利用高保真酶進行PCR,退火溫度為60 ℃,PCR 產物檢測(1%瓊脂糖凝膠)、膠回收(E.Z.N.A.○RGel Extraction Kit)、載體連接等方法參照本實驗室方法[24],選取陽性克隆送北京擎科生物有限公司(西安測序部)進行測序。
1.4 AcNPR1 序列的生物信息學分析
利用ORF finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)在線分析開放閱讀框,通過SMART 在線分析Ac NPR1蛋白序列的保守結構域(https://smart.embl-heidelberg.de/);利用DNAMAN 9.0 軟件對獼猴桃Ac NPR1 蛋白質與其他物種NPR1 蛋白質進行比對分析;使用Clutsal Xv1.83 程序進行多序列比對,然后將比對結果輸入到MEGA 7.0 軟件中,利用鄰接法(neighbor-joining,NJ)構建系統發育樹,Bootstrap 值 取1 000 次;利 用ExPASy-ProtParam tool 在線軟件(http://web.expasy.org/protparam/)進行蛋白質理化性質分析;利用Expasy中的ProtScale 在線軟件(http://web.expasy.org/protscale/)分析Ac NPR1蛋白質的親水性和疏水性;利用NetPhos v3.1(http://www.cbs.dtu.dk/services/NetPhos/)在線工具對獼猴桃Ac NPR1蛋白進行潛在的磷酸化位點預測分析;利用SOPMA 數據庫(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)得 到Ac NPR1 可能的二級結構;利用SWISS-MODEL(http://swissmodel.expasy.org)進行同源建模,構建Ac-NPR1的三級結構域模型;通過TMPred(https://embnet.vital-it.ch/software/TMPRED_form.html)預測該蛋白跨膜區域;通過CELLO2GO 在線分析工具(http://cello.life.nctu.edu.tw/cello2go/)預測亞細胞的可能定位。
1.5 AcNPR1 基因實時熒光定量分析
熒光定量反應程序及反應體系按照熒光定量試劑盒(Perfectstart Green qPCR Supermix,全式金生物,北京)說明完成,采用美國ABI Step One-Plus實時熒光定量PCR 儀系統進行實時熒光定量分析。熒光定量上、下游引物分別為5′-CACAAGGCTTAGACTCAGA-3′和5′-CAAGCACTTCAGACACAAC-3′。內參基因上下游引物分 別 為5′-CTGTGAAACTGCGAATGGCTC-3′和5′-TTCCAGAAGTCGGGGTTTGT-3′。采用3次生物學重復,不同材料中目的基因的相對表達量采用2-ΔΔCT方法計算。利用SPSS 21.0統計分析軟件進行數據處理。
2 結果與分析
2.1 Ac NPR1 基因克隆以及序列分析
以病原菌Psa處理12 h后的‘紅陽’、‘徐香’獼猴桃葉片為試驗材料,提取總RNA 反轉錄后進行PCR 擴增,克隆至T 載體并送樣測序,得到‘紅陽’Ac NPR1基因序列全長為1 770 bp(Gen-Bank登錄號為MW881148)(圖1-A)。經測序發現‘徐香’Ac NPR1基因序列只有個別堿基與‘紅陽’不同(未列出)。本研究進一步對‘紅陽’序列進行生物信息學分析,利用NCBI 網站中ORF Finder在線工具分析該序列開放閱讀框,發現該基因序列開放閱讀框長為1 770 bp,可編碼589個氨基酸;利用SMART 分析該蛋白序列的保守結構域,結果表明在第64位至第190位,第266位至361位,第371位至575位區域間分別含有N 端保守的BTB-POZ、錨蛋白重復結構域An Ks、C端NPR1_like_C 植 物NPR1 蛋 白 所 共有的保守結構域(圖1-B)。
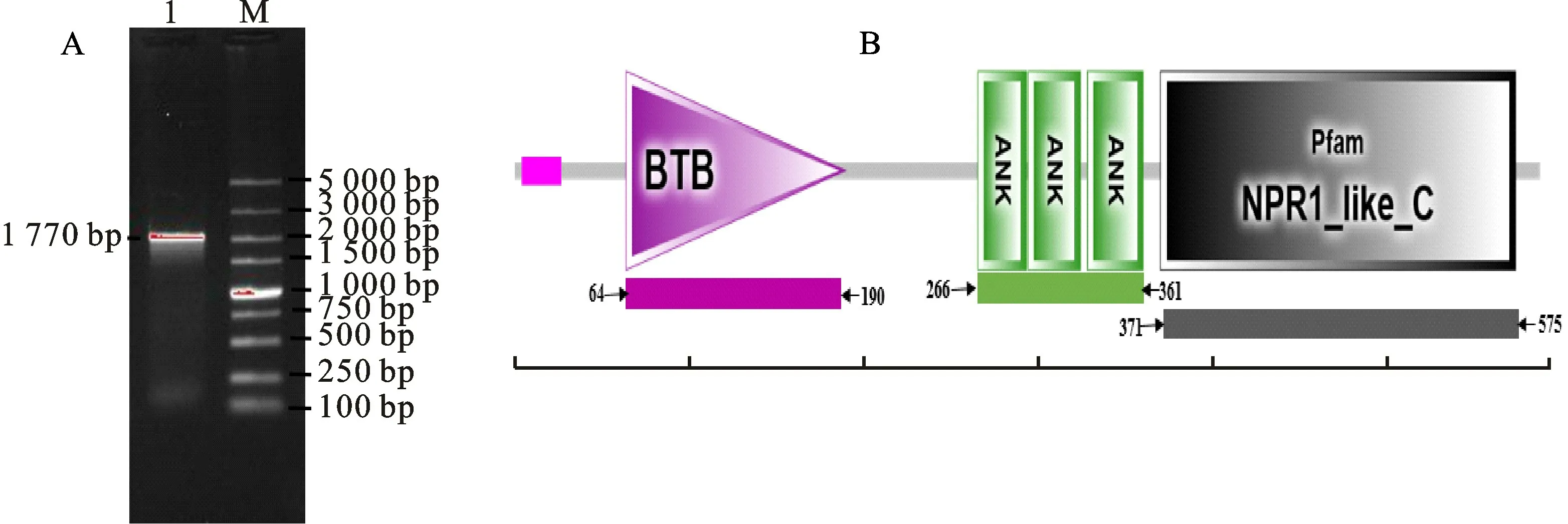
圖1 ‘紅陽’AcNPR1 基因PCR 擴增(A)和AcNPR1蛋白的保守結構域(B)Fig.1 The PCR amplified product of AcNPR1 gene(A)and the conserved domain of AcNPR1 protein(B)in‘Hongyang’
2.2 AcNPR1蛋白的理化性質及磷酸化位點分析
利用在線工具ProtParam 分析Ac NPR1基因編碼的蛋白分子式為C2895H4639N799O889S29,分子質量為65 792.24 ku,理論等電點(pI)為6.27,其中異亮氨酸和絲氨酸的使用頻率最高,均占所有氨基酸的10.9%;然后依次為丙氨酸,含量最少的為色氨酸,占所有氨基酸的0.2%。Ac NPR1蛋白的不穩定指數為43.75;總平均親水性為-0.245,脂肪系數為89.56。ProtScale在線軟件分析蛋白質的親水性和疏水性,發現該蛋白為親水性蛋白(圖2-A);利用NetPhos對該蛋白的磷酸化位點進行分析,發現Ac NPR1蛋白均具有絲氨酸(Ser)、蘇氨酸(Thr)和酪氨酸(Try)磷酸化位點(圖2-B)。通過TMPred 預測該蛋白跨膜區域,跨膜結構域預測結果顯示獼猴桃Ac NPR1蛋白可能為一個跨膜蛋白,分別在第5~26、115~134、160~182、193~216個氨基酸處含有4個從內到外的跨膜結構域(圖2-C)。
2.3 AcNPR1蛋白二、三級結構特征和亞細胞定位預測分析
通過SMOPA 數據庫得到二級結構預測結果表明,AcNPR1蛋白質二級結構主要是由α螺旋、β折疊、延伸鏈、無規則卷曲所組成,占據比例分別為53.14%、3.57%、7.47%、35.82%(圖3-A);其中α螺旋和無規則卷曲所占比例較高,為其主要的二級結構元件。通過SWISS-MODEL軟件預測Ac NPR1蛋白質三級空間結構模型,含有α螺旋至少35個,β折疊至少7個,無規則卷曲至少25個,主要是以螺旋為基本結構單元形成的平行或反向平行螺旋結構(圖3-B)。利用CELLO2GO 在線分析工具對Ac NPR1蛋白亞細胞的可能定位進行預測分析,結果顯示,獼猴桃Ac NPR1 蛋白亞細胞定位最可能位于細胞核(表1)。
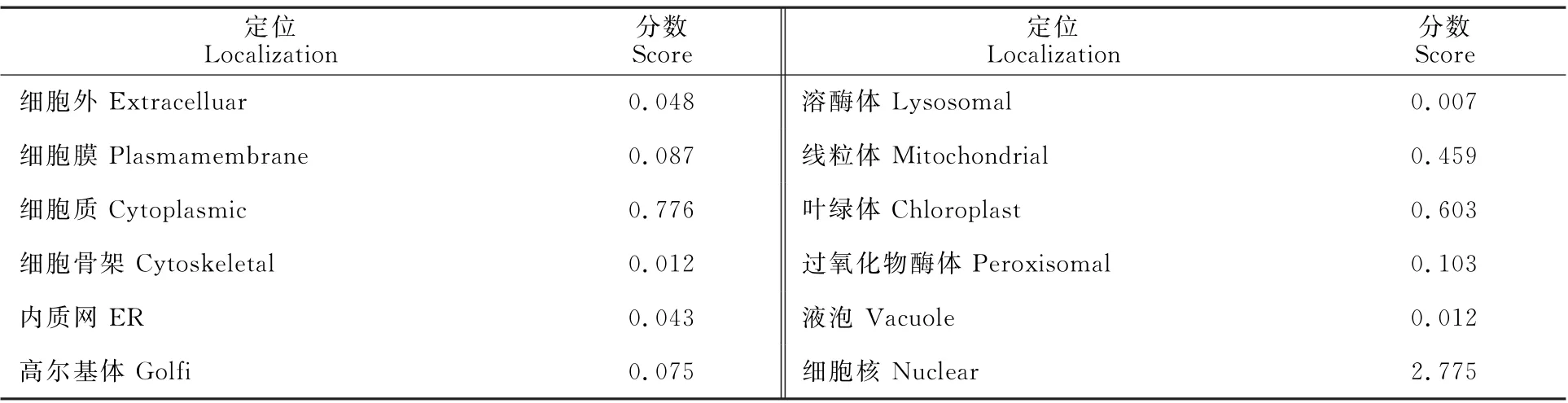
表1 亞細胞定位預測Table 1 Prediction of AcNPR1 protein in‘Hongyang’
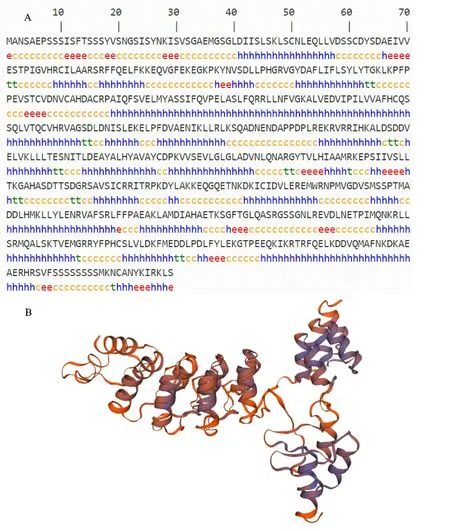
圖3 ‘紅陽’獼猴桃AcNPR1蛋白質的二級結構(A)、三級結構(B)預測Fig.3 Prediction of secondary(A)and tertiary structure(B)of AcNPR1 protein in‘Hongyang’
2.4 AcNPR1 基因編碼的氨基酸序列的比對分析
利用DNAMAN 7.0軟件,將克隆得到的‘紅陽’獼猴桃Ac NPR1基因編碼的氨基酸序列與在NCBI網站上獲取其他物種的NPR1氨基酸序列進行序列比對(圖4),結果表明獼猴桃Ac NPR1氨基酸序列與茶樹(XP_028062623.1)、葡萄(XP_002274045.1)、銀白楊(XP_034927243.1)、酸棗(XP_015882955.1)、煙草(XP_016442273.1)和胡桃(XP_018835840.1)的NPR 類轉錄因子保守結構域一致性較高。在N 端包含BTB_POZ結構域、Ank2保守域,C 端有NPR1_Like_C 結構域,與大多數物種NPR1 蛋白序列相似,蛋白相似性分別為83.94%、79.63%、78.10%、76.27%、75.62%和74.28%。

圖4 ‘紅陽’獼猴桃AcNPR1 與不同物種NPR1 轉錄因子氨基酸序列多重比對Fig.4 Multiple alignment of amino acid sequences of AcNPR1 in‘Hongyang’and NPR1 transcription factors in different species
采用 MEGA 7.0 軟件分析8 個物種的NPR1蛋白序列與‘紅陽’獼猴桃Ac NPR1 蛋白序列進行系統進化分析并構建進化樹(圖5),結果顯示NPR1蛋白被大致分為兩組,‘紅陽’獼猴桃Ac NPR1蛋白與銀白楊、簸箕柳、大豆、葡萄、酸棗、茶樹聚為一組,‘紅陽’獼猴桃Ac NPR1 蛋白與茶樹CsNPR1 蛋白親緣關系最近。
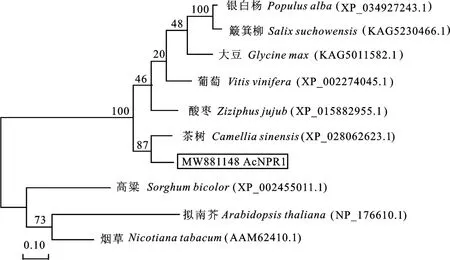
圖5 ‘紅陽’獼猴桃AcNPR1蛋白系統進化樹分析Fig.5 Phylogenetic tree analysis of AcNPR1 protein in‘Hongyang’
2.5 AcNPR1 基因表達分析
利用實時熒光定量PCR 對‘紅陽’和‘徐香’不同組織中的莖、葉、花Ac NPR1基因表達模式進行分析。結果表明,Ac NPR1基因在‘徐香’品種不同組織的表達量從大到小依次為葉、花、莖(圖6-A);在‘紅陽’品種同樣是在葉片中的表達量最高,其次是莖,表達量最小的為花(圖6-B);兩品種Ac NPR1基因均在葉片中表達量最高。
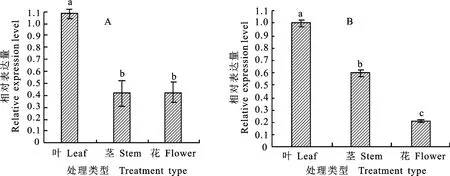
圖6 ‘徐香’獼猴桃(A)和‘紅陽’獼猴桃(B)AcNPR1 基因組織表達分析Fig.6 Expression analysis of AcNPR1 gene in different tissues of‘Xuxiang’(A)and‘Hongyang’(B)
以正常生長處理組(CK)為對照,分析Psa以及SA+Psa處理下不同抗性品種獼猴桃在不同處理時間下Ac NPR1基因表達水平變化情況。結果表明,在Psa處理下,0 h 時‘紅陽’品種中Ac NPR1基因相對表達量低于對照,而‘徐香’中Ac NPR1基因相對表達量迅速上升至對照的3.7倍;6 h后兩品種中Ac NPR1基因相對表達量均呈現先升高后下降的趨勢(圖7),‘紅陽’和‘徐香’品種分別在48 h和24 h時基因相對表達量達到最大,Psa處理下‘紅陽’Ac NPR1基因相對表達量始終低于‘徐香’(圖7-A)。在SA+Psa處理下,0 h時‘紅陽’品種中Ac NPR1基因相對表達量略高于對照,而‘徐香’中Ac NPR1基因相對表達量迅速升高至對照的2.9倍;之后‘徐香’Ac NPR1基因相對表達量均呈現先升高后下降的趨勢,在24 h時基因相對表達量達到最大為對照的7.7倍,‘紅陽’Ac NPR1基因相對表達量在6 h至48 h內波動變化,72 h達到最大,為對照的1.6倍,且該處理下‘紅陽’基因表達量始終低于‘徐香’(圖7-B)。不同處理下,‘徐香’品種Ac-NPR1基因相對表達量高于‘紅陽’,且‘徐香’中基因相對表達量達到最高的時間早于‘紅陽’。
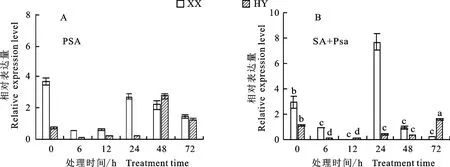
圖7 病原菌(A)、SA加病原菌(B)處理下兩品種葉片AcNPR1 基因表達量Fig.7 Expression of AcNPR1 gene in leaves of two varieties under Psa(A)and(SA+Psa)(B)treatment
3 討論
NPR1基因是介導植物免疫系統的重要調控因子[20],病原菌侵染時其特異性結構域與轉錄因子相結合激活植物免疫系統,相關抗病基因如PR 基因表達量上調,從而提高植物抗病能力。本研究以病原菌侵染12 h 后的高感病品種‘紅陽’和抗病品種‘徐香’獼猴桃葉片為試驗材料,克隆得到兩個品種獼猴桃AcNPR1基因序列,對‘紅陽’Ac NPRN1基因進行生物信息學分析發現,該基因完整開放閱讀框(ORF)大小為1 770 bp,編碼589個氨基酸,含有典型的BTB/POZ結構域、ANK 錨蛋白重復序列和NPR1_like_C 3個結構域,多重序列比對發現其與已報道的茶樹、葡萄、煙草等物種中NPR1類蛋白保守結構域一致,系統進化樹分析顯示Ac NPR1蛋白與茶樹親緣關系最近。
在擬南芥、水稻、番茄以及小麥中過量表達At NPR1基因能夠提高植物對細菌和真菌的抗性[4,11-13];在水稻中過量表達OsNPR1基因提高了水稻對細菌性枯萎病的抗病性[14];與之類似,在兩個蘋果栽培品種中過表達MalusNPR1基因后,發現其激活了PR基因增強了其對火疫病及兩種蘋果的重要真菌病原體的抗性[15]。研究表明,毛花獼猴桃(Actinidia eriantha)接種獼猴桃潰瘍病病原菌后發現,Ae NPR1基因被誘導上調表達,從而提高了毛花獼猴桃的抗病性[25]。本研究中在獼猴桃潰瘍病致變菌侵染獼猴桃后,抗病品種‘徐香’和高感病品種‘紅陽’中Ac NPR1基因表達量均被上調,表明Ac NPR1基因在獼猴桃抵御潰瘍病過程中發揮了一定作用;并且‘紅陽’品種Ac NPR1基因表達量始終低于抗病品種‘徐香’,抗病性不同的獼猴桃品種在響應病原菌脅迫的抗病機制方面存在差異,抗性品種為了抵御病原菌造成的傷害,會迅速啟動抗性反應,在抗病品種‘徐香’中AcNPR1基因被迅速誘導表達,參與了植物對病原菌的早期抗性反應,從而啟動下游與抗病相關基因的表達,這與水稻、百合和猴面花中NPR1基因表達結果一致[14,26-27]。
水楊酸(SA)是一種酚類植物激素,能夠作用于植物的生長發育過程,也是植物免疫系統的主要調控激素之一,在植物抵御生物營養性和半生物營養性病原菌方面起著重要作用[28]。SA 介導的系統獲得性抗性(Systemic Acquired Resistance,SAR)是植物的一種廣譜抗病機制,NPR1是其關鍵調節因子之一,是將SA 信號轉化為PR相關基因表達所必需的[29]。研究表明,外源SA、JA 等可以誘導NPR1基因或其同源基因表達從而激活植物抗病反應[30-32]。本研究中經SA 預處理后,在抗病品種‘徐香’和高感病品種‘紅陽’中Ac NPR1基因表達水平均升高,表明SA 對誘導Ac NPR1基因表達從而激活獼猴桃抗病反應具有一定作用,與上述報道中結果一致。此外,侯琿等[33]的研究表明輪紋病抗病蘋果品種的抗性基因更容易受到SA 的誘導表達,這與本研究結果類似,在抗病品種‘徐香’中Ac NPR1基因表達變化受SA 誘導影響較大。SA 預處理過的抗病品種‘徐香’Ac NPR1基因表達量約是未做SA 預處理組的2倍,而感病品種‘紅陽’則相反;這表明不同抗性獼猴桃品種間NPR1響應SA 誘導表達的調控機制可能存在差異。研究表明,SA 含量影響植物抗病反應的發生、強度及快慢,SA、病原菌的誘導可以使植物體內SA 水平升高,不同抗性品種間SA 水平的差異可能造會成抗性反應及NPR1等相關抗性基因表達的差異。但是,抗病信號傳遞途徑是一個非常復雜的體系,不同抗病性品種間Ac NPR1基因表達為什么會呈現出差異,需要后續進一步深入研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
