幽門螺桿菌代謝產物腺苷通過A2A受體調控炎癥因子分泌的機制研究
2022-07-11 01:19:08羅萍王效惠向軍英劉哲
實用醫學雜志 2022年10期
羅萍 王效惠 向軍英 劉哲
成都大學附屬醫院消化內科(成都610081)
作為革蘭氏陰性菌的幽門螺桿菌(H.pylori)是第一個被正式認可的細菌致癌物,感染了世界上大約44 億(59%)的人口[1]。慢性感染產生炎癥狀態,但在大多數受試者中是無癥狀的[2]。然而,在一部分幽門螺桿菌感染人群中,胃部炎癥可能演變為慢性胃炎、消化性潰瘍、胃黏膜相關淋巴組織(MALT)淋巴瘤和胃癌[3-5]。簡而言之,幽門螺桿菌定植的階段如下:穿過胃黏液層,黏附到胃上皮,然后在避免被宿主免疫反應擊敗的同時獲得營養。對于一些幽門螺桿菌定植/毒力因子,已經提出其在炎癥發展和對宿主免疫系統的影響中的特定作用[6]。另一方面,已經發現一些宿主因素與幽門螺桿菌感染的起源(慢性胃炎癥)和隨后的病理結果有關[7]。因此,細菌作用和宿主反應都參與發病機制。宿主-病原體共同適應導致幽門螺桿菌定植,在大多數情況下可能保持無害;然而,病原體和宿主的特定基因型的同時發生可能會導致嚴重病理的發展[8]。盡管自發現H. pylori以來已經提供了大量的臨床和實驗觀察結果,但仍需要進一步了解宿主-病原體相互作用的機制以及導致對這種病原體獲得保護性免疫的機制的證據,以指導和制定有效對抗幽門螺桿菌。宿主-微生物相互作用是健康和疾病的基本組成部分[9]。維持人類與微生物組之間健康共生狀態的一個關鍵參數是微生物群產生獨特的代謝物,這些代謝物既為宿主提供營養,也參與免疫發育的調節[10]。然而,關于H. pylori代謝物是否調控宿主炎癥因子的分泌尚不清晰。基于此,本研究旨在探討H.pylori代謝產物參與調控宿主炎癥因子的潛在機制。
1 材料與方法
1.1 材料 幽門螺桿菌菌株H. pyloriSS1 獲贈于中科院微生物所。大腸桿菌E.Coli購自北京博邁德基因技術有限公司(貨號:BC102-03)。C57BL/6小鼠購自廣州銳格生物科技有限公司(貨號:C570005)。布魯氏菌肉湯購自ELITE-MEDIA 公司(貨號:M1070-01)。RPMI1640 培養基購自維森特生物技術(南京)有限公司(貨號:350-000-CL*10)。人胃癌細胞AGS 購自北京欣盛百泰科技有限公司(貨號:CL0031)。廣州一科生物科技有限公司購自(貨號:BS7772)。腺苷檢測試劑盒購自上海群己生物科技有限公司(貨號:KA4547)。抗幽門螺桿菌免疫球蛋白(IgG)檢測試劑盒購自北京科瑞美科技有限公司(貨號:QS440220-48T)。人IL-33檢測試劑盒購自深圳市文樂生物科技有限公司(貨號:JL19282)。人TNF-α檢測試劑盒購自北京江晨文軒生物科技有限責任公司(貨號:JL10208-1)。人IFN-γ 檢測試劑盒購自北京江晨文軒生物科技有限責任公司(貨號:JL28106-1)。鼠IL-33 檢測試劑盒購自廣州徠智生物科技有限公司(貨號:2MKMLJM219447m)。鼠TNF-α 檢測試劑盒購自昆明皇寶商貿有限公司(貨號:ml002098-96T(JK))。鼠IFN-γ 檢測試劑盒購自Elabscience 公司(貨號:E-EL-SR001)。SiGFP、siA2AR 由南京金斯瑞設計和合成。
1.2 方法
1.2.1 細胞培養與感染 幽門螺桿菌菌株SS1是一種適應小鼠的人類分離物,用于所有實驗。H. pyloriSS1 的甘油原液首先在5%羊血瓊脂平板上在微需氧條件下生長2 d。平板培養后,H.pyloriSS1 在5%FCS 的布魯氏菌肉湯中在37 ℃微需氧條件下溫和攪拌培養14 ~16 h。人胃癌細胞AGS 維持在補充有10%滅活FBS 的RPMI1640 培養基中。AGS 細胞血清饑餓16 h,然后以200∶1 的感染復數(MOI)感染幽門螺桿菌。當AGS 細胞感染H.pylori或E.Coli時,它們的融合水平為60% ~70%。感染后幾乎細胞附著在細胞培養皿上,并且與對照細胞相比,細胞數量也沒有減少,因此判斷MOI 為200∶1 的H.pylori感染后的對AGS 細胞的增殖無顯著影響。本研究均使用lipo3000 進行相關轉染。
1.2.2 幽門螺桿菌菌株SS1感染小鼠H.pyloriSS1由平板和液體培養物制備。每周3 次,在400 μL 5% FCS/布魯氏菌肉湯中給C57BL/6 小鼠口服3 ×108CFU 的H.pyloriSS1,持續4 周。
1.2.3 液相色譜與串聯質譜鑒定幽門螺桿菌菌株SS1 的代謝物 對H.pyloriSS1 感染AGS 細胞后的上清進行3 KDa 過濾器的過濾,收集濾液液進行LC/MS 分析。LC/MS 測量使用配備兩個LC-30AD泵、DGU-20A5 脫氣機、SIL-30AC 自動進樣器、CTO-20AC 柱溫箱和 一個CBM-20A 控制模塊,連接到LCMS-8040 三重四極桿質譜儀。ChromaTOF 2.32版用于數據預處理,無需平滑、3 s 峰寬、略高于噪聲水平的基線減法,以及在整個色譜圖中以5∶1的信號/噪聲水平進行自動質譜去卷積和峰檢測。定點質量被報告用于BinBase 算法。結果以txt 文件形式導出到數據服務器,并由代謝組學BinBase數據庫中實施的過濾算法進一步處理。BinBase算法(rtx5)使用以下設置:色譜圖的有效性(<10 個峰,強度>107 個計數s-1),無偏保留指數標記檢測(MS 相似性>800,高m/z 標記離子強度范圍的有效性),并通過5 階多項式回歸計算保留指數。使用以下匹配過濾器將光譜切割至5%的基峰豐度并從最高到最低豐度光譜與數據庫條目匹配:保留指數窗口,±2 000 單位(相當于約±2 s 保留時間);驗證獨特的離子和頂點質量(獨特的離子必須包含在頂點質量中,并且以>3% 的基峰豐度存在);和質譜相似性必須符合取決于峰純度和信噪比以及最終異構體過濾器的標準。
1.2.4 酶聯免疫吸附實驗 抽取處理后小鼠尾靜脈血清或者細胞上清進行酶聯免疫吸附實驗。酶聯免疫吸附實驗檢測腺苷、抗幽門螺桿菌免疫球蛋白(IgG)、IL-33、TNF-α、IFN-γ 的水平。
1.2.5 腺苷合成酶敲除株H.pyloriΔAdA菌株的構建和驗證 使用Red/ET方法[11]在H.pyloriSS1菌株中構建AdA缺失編碼的菌株H.pyloriΔAdA。H.pyloriSS1通過電轉染PSQ 質粒轉化,攜帶PSQ 的H. pyloriSS1 細胞能夠進一步轉化靶向目標基因組基因座的重組DNA 片段。通過PCR 后瓊脂糖凝膠電泳確認AdA 被敲除,引物序列:AdA-F:5'-GGAGACCAGCGATATCCACG-3'、AdA-R:5'-ATGCGAGTCAGACCGTTGTT-3'、GAPDH-F:5'-CTGCCGCTAACGCAGAAATC-3'、GAPDH-R:5'-CCATAGACAAAGGTGGGCGT-3'。
1.2.6 統計學分析 使用Student'st檢驗分析數據,P<0.05 表示差異具有統計學意義。
2 結果
2.1 幽門螺旋桿菌代謝物抑制炎癥因子分泌E.Coli或H. pylori感染后,人胃癌細胞AGS 中IL-33、TNFα、IFN-γ 的分泌水平上升。相比于感染E.Coli,感染H. pylori的AGS 細胞分泌的IL-33、TNF-α、IFN-γ的水平較低,見圖1A-C。H. pylori感染AGS 細胞24 h后,將收取細胞上清進行3 kDa過濾器過濾,濾液為<3 kDa 的分子(一般為代謝物),未過完3 kDa過濾器的液體為>3 kDa的分子(一般為蛋白,圖1D),將這些組分處理E.Coli刺激過的AGS 細胞16 h后,發現小于3 kDa的分子能夠降低IL-33、TNF-α、IFN-γ的分泌水平,見圖1E-G。
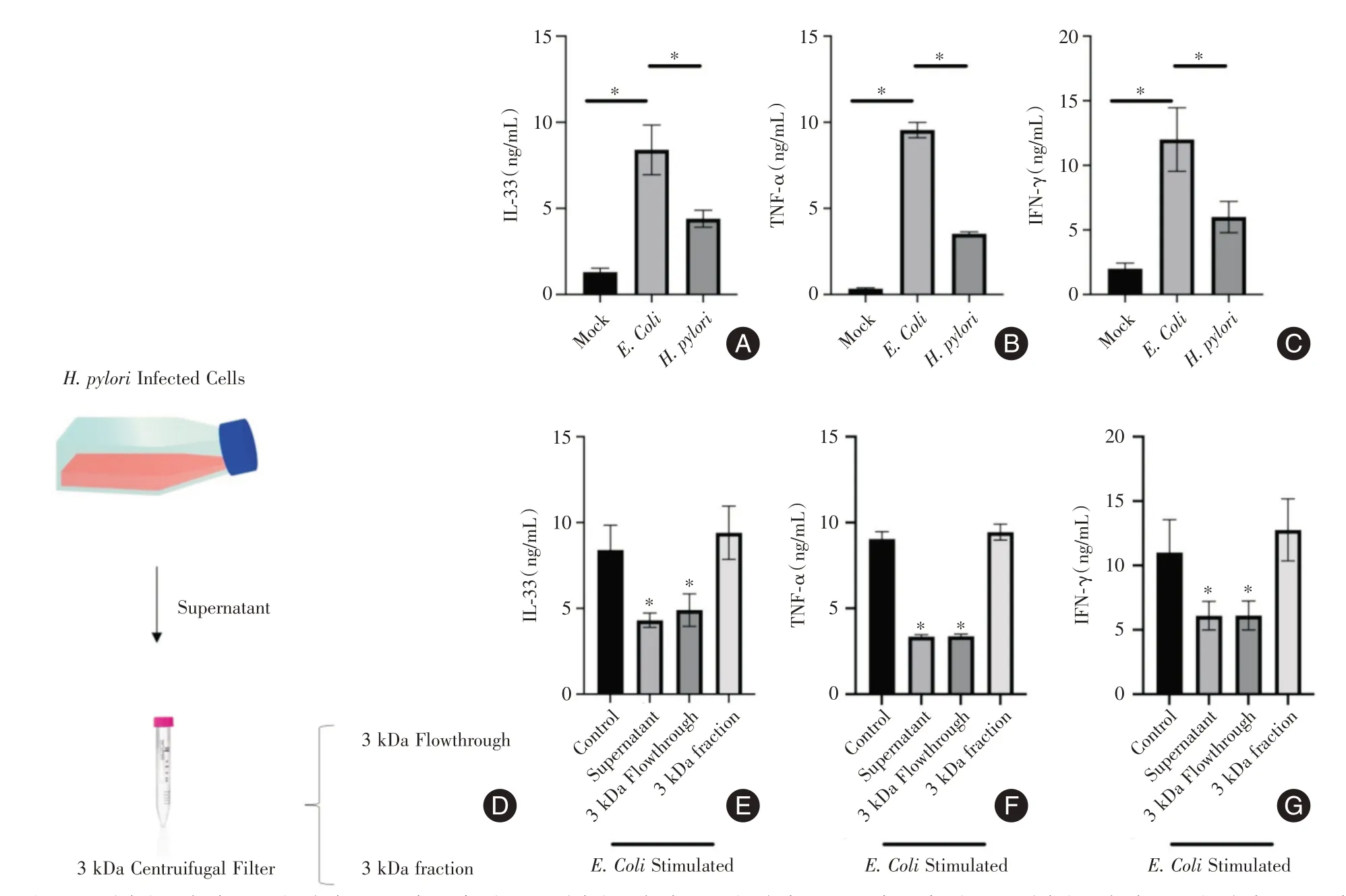
圖1 幽門螺旋桿菌代謝物抑制炎癥因子分泌Fig.1 Helicobacter pylori metabolites inhibit the secretion of inflammatory factors
2.2 腺苷抑制炎癥因子分泌E.Coli或H. pylori感染AGS 細胞24 h 后,將收取細胞上清進行液相色譜與串聯質譜鑒定H.pylori的代謝物,發現多種代謝物的表達水平存在差異,見圖2A。將上述代謝物小分子以5 μmol/L 的濃度進行篩選,發現腺苷能夠影響IL-33 的分泌水平,見圖2B。此外,腺苷降低IL-33、TNF-α、IFN-γ 的分泌水平存在梯度依賴效應,見圖2C-E。
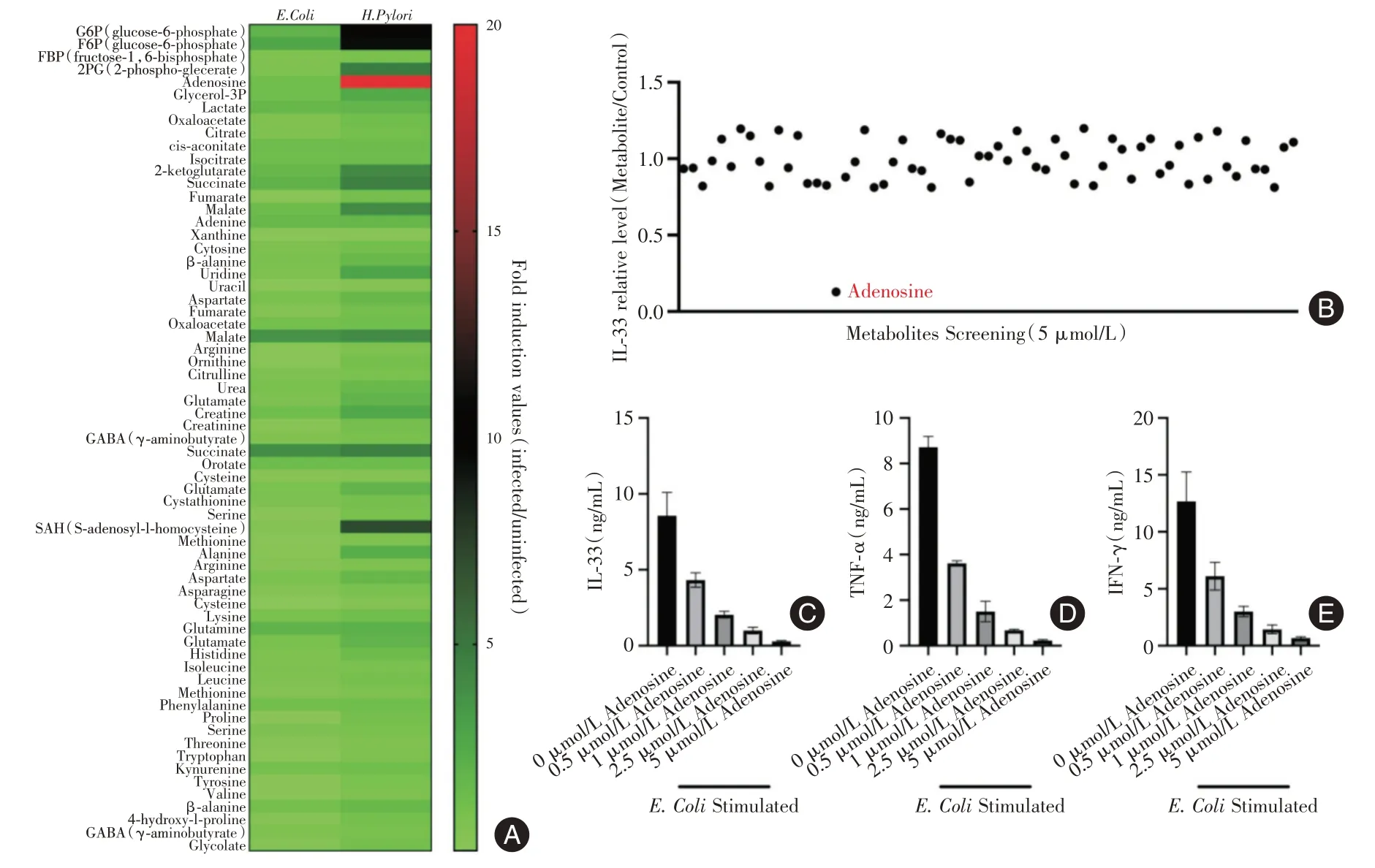
圖2 腺苷抑制炎癥因子分泌Fig.2 Adenosine inhibits the secretion of inflammatory cytokines
2.3 腺苷合成酶/腺苷/A2A 受體軸調控炎癥因子分泌 通過NCBI 數據庫blast 功能發現H. pylori存在腺苷合成酶(NCBI Reference Sequence:WP_078247416.1),見圖3A。構建腺苷合成酶敲除株H. pyloriΔAdA菌株,見圖3B。H.pyloriΔAdA感染后,AGS細胞的腺苷水平無顯著改變,見圖3C。此外,H.pyloriΔAdA感染后,AGS細胞中IL-33、TNF-α、IFN-γ的分泌水平均比H. pyloriWT感染的AGS 細胞高,見圖3D-F。A2A 受體是腺苷調控下游分子的關鍵[12],敲低A2A 受體后,H. pylori感染的AGS 細胞IL-33、TNF-α、IFN-γ的分泌水平上升,見圖3G-I。
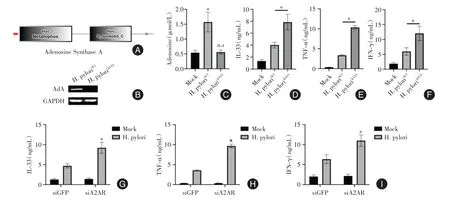
圖3 腺苷合成酶/腺苷/A2A 受體軸調控炎癥因子分泌Fig.3 Adenosine synthase/adenosine/A2A receptor axis regulates the secretion of inflammatory cytokines
2.4 幽門螺旋桿菌腺苷合成酶調控小鼠免疫反應H.pyloriΔAdA或H.pyloriWT感染小鼠4周后,與Mock組比較,H. pyloriWT感染和H. pyloriΔAdA感染胃組織出現囊性擴張,排列不規則和炎癥細胞浸潤,說明感染后胃組織有炎癥發生見圖4A。另外,發現小鼠血清中的IL-33、TNF-α、IFN-γ 的水平均上升。相比于H.pyloriWT感染,H.pyloriΔAdA感染后小鼠血清中的IL-33、TNF-α、IFN-γ 的水平上升更明顯,見圖4B-D。此外,相比于H.pyloriWT感染,H.pyloriΔAdA感染后小鼠產生的抗幽門螺旋桿菌抗體的滴度上升,見圖4E。

圖4 幽門螺旋桿菌腺苷合成酶調控小鼠免疫反應Fig.4 Helicobacter pylori adenosine synthase regulates the immune response in mice
3 討論
H.pylori感染會引起多種胃腸道疾病,如無癥狀慢性胃炎、消化性潰瘍病、胃腺癌和胃淋巴瘤,以及相關的分子機制在對宿主和細菌的研究中開始闡明這些疾病的基礎[13]。然而,很少有研究使用代謝組分析評估幽門螺桿菌感染產生的代謝物。在一項研究中,從已感染幽門螺桿菌(一種已知可在沙鼠中長期適應的臨床分離物)的沙鼠中收集尿液,并使用質子核磁分析尿液中的代謝物共振光譜法[14]。此外,研究表明,幽門螺桿菌感染會改變腸道微生物群,如微生物相關代謝物的變化所示[15]。然而,幽門螺桿菌的代謝物如何影響宿主并實現免疫逃逸尚未明確。本研究通過代謝組學和遺傳學操作鑒定了幽門螺桿菌代謝物腺苷對炎癥因子IL-33、TNF-α、IFN-γ 的分泌水平的影響。
此前,有研究基于代謝物譜評估了幾種代謝途徑,包括糖酵解途徑、TCA 循環、膽堿途徑、尿素循環、谷胱甘肽循環、嘌呤途徑、嘧啶途徑和氨基酸代謝,并在一些中觀察到H.pylori感染誘導的改變宿主代謝途徑[16]。本研究發現H.pylori感染后,AGS 細胞中腺苷水平顯著上升。腺苷具有免疫抑制特性,并且與癌癥發展有關。因此,H. pylori可能通過代謝物腺苷參與宿主免疫逃逸。
本研究鑒定并表征了H.pylori的腺苷合成酶,一種具有5'-核苷酸酶特征序列的H. pylori細胞壁錨定蛋白。構建腺苷合成酶敲除株H. pyloriΔAdA菌株,發現H. pyloriΔAdA感染后,AGS 細胞的腺苷水平無顯著改變。此外,H. pyloriΔAdA感染后,AGS 細胞中IL-33、TNF-α、IFN-γ 的分泌水平均比H. pyloriWT感染的AGS 細胞高。此外,相比于H.pyloriWT感染,H. pyloriΔAdA感染后小鼠產生的抗幽門螺旋桿菌抗體的滴度上升。因此,腺苷合成酶是感染期間腺苷合成所必需的,并且此蛋白有助于H.pylori在動物體內存活。
A2A 受體是腺苷調控下游分子的關鍵,敲低A2A 受體后,H. pylori感染的AGS 細胞IL-33、TNFα、IFN-γ 的分泌水平上升。有研究發現腺苷能夠與A2A 受體結合并且調控NF-κB 通路,抑制細胞因子的表達[17]。但是本研究僅檢測了細胞因子的分泌水平,其表達水平是否受到了調控值得進一步探討。并且,本研究仍然具有一定的局限性,腺苷合成酶/腺苷/A2A 受體調控軸是在細胞水平中鑒定的,其是否在動物水平也發揮相同的作用值得進一步探討。
綜上所述,H. pylori表達的腺苷合成酶能夠促進腺苷的產生,隨后腺苷通過A2A 受體調控炎癥因子IL-33、TNF-α、IFN-γ 的分泌,并且降低適應性免疫水平。腺苷合成酶/腺苷/A2A 受體可能是對抗幽門螺桿菌的潛在靶點。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
學苑創造·A版(2015年11期)2016-01-14 09:03:27
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
