A激酶錨定蛋白9提高ERK通路活性促進肺腺癌細胞A549增殖和遷移
2022-05-31 05:47:52傅揚揚黃曉穎王良興
溫州醫科大學學報 2022年5期
傅揚揚,黃曉穎,王良興
溫州醫科大學附屬第一醫院 呼吸與危重癥醫學科,浙江 溫州 325015
肺癌是一種最常見的惡性腫瘤,其病死率在腫瘤中位居第一[1]。近年來,分子靶向治療和免疫治療明顯改善了腫瘤患者的預后,但肺癌的5年生存率仍然只有22%[2]。因此,發現新的腫瘤相關基因和治療靶點對肺癌的診斷和治療具有重要的意義。A激酶錨定蛋白9(A kinase anchoring protein 9, AKAP9)是一種參與微管成核的高爾基體/中心體支架蛋白[3-4]。研究發現,AKAP9參與了結直腸 癌[5]、甲狀腺癌[6]、乳腺癌[7]、急性髓系白血病[8]的發生或轉移。在一項大規模的肺癌相關全基因組篩查中,AKAP9被發現是潛在的易感基因[9]。在前期的研究中,我們通過Kaplan-Meier plotter平臺分析AKAP9的表達水平與肺癌預后的關系,結果發現AKAP9表達高的患者生存時間明顯縮短。然而,AKAP9在肺腺癌細胞中的確切作用尚未明了,因此本研究通過體外實驗探討AKAP9對肺腺癌細胞A549增殖和遷移的作用,并進一步闡明其可能的分子機制。
1 材料和方法
1.1 材料
1.1.1 細胞:肺腺癌細胞A549購自中國科學院典型培養物保藏委員會細胞庫/中國科學院上海生命科學研究院細胞資源中心。
1.1.2 主要試劑:shRNA購自上海吉凱基因醫學科技股份有限公司;MTT試劑購自美國Sigma-Aldrich公司;SYBR Green購自天根生化科技(北京)有限公司;MLV-reverse transcriptase試劑盒購自美國Thermo Fisher Scientific公司;細胞周期試劑盒購自美國BD Pharmingen公司;AKAP9抗體購自美國Santa Cruz公司;P27、細胞周期素D1(Cyclin D1)、細胞周期素依賴性激酶4(cyclin-dependent kinase 4, CDK4)、表皮生長因子受體(epidermal growth factor receptor, EGFR)、磷酸化EGFR(p- EGFR)、細胞外信號調節激酶(extracellular signal- regulated kinase, ERK)、磷酸化ERK(p-ERK)、β-actin抗體購自美國Cell Signaling Technology公司。
1.2 方法
1.2.1 細胞培養:A549在含有10%胎牛血清的F12K完全培養基中,放置在37 ℃、5% CO2的恒溫培養箱中進行培養,每3天更換1次新鮮培養基。
1.2.2 實時定量聚合酶鏈反應(real-time quantitative polymerase chain reaction, RTqPCR)檢測目的基因mRNA表達量:提取肺腺癌細胞的總RNA,利用MLV-reverse transcriptase試劑盒將其反轉錄為cDNA,然后用RealMssterMix(SYBR Green)試劑盒進行RT-qPCR反應,GAPDH基因作為內參。實驗結果采用2-△△Ct相對定量分析法,并以GAPDH基因作為內參基因來計算轉錄的相對差異。AKAP9引物序列為F:5’-ACTCAAGGCACAGCATAAA CAC-3’,R:5’-GTTCTTCACTGCGTCCCAA-3’。GAPDH引物序列為F:5’-ACAGTCAGCCGCATCTTCTT-3’,R:5’-GACA AGCTTCCCGTTCTCAG-3’。
1.2.3 蛋白質印跡(Western blot)法檢測細胞周期相關蛋白和ERK通路相關蛋白的表達:利用 Bradford法檢測蛋白濃度,等量的蛋白樣品經聚丙烯酰胺凝膠電泳分離后轉印至PVDF膜上。PVDF膜用5%牛奶封閉2 h,隨后一抗4 ℃孵育過夜,二抗室溫孵育2 h。蛋白條帶用ECL系統進行檢測。
1.2.4 shRNA感染實驗:特異性shRNA及陰性對照shRNA由上海吉凱基因醫學科技股份有限公司合成。取1×105個細胞接種于6孔板,根據細胞的感染復數值感染相應的病毒量,利用Western blot、RTqPCR檢測敲減效率。經敲減處理后的細胞用于后續實驗。病毒感染細胞的兩個靶點序列為shAKAP9 1#:GAAUAUUGAUGGUACAAUATT,shAKAP9 2#:GCACA AUAAUUAUUGAAUUTT。
1.2.5 MTT實驗檢測細胞增殖:以每孔3 000個細胞接種到96孔板,設3個復孔,每3天更換一次新鮮培養基,分別培養1、3、5 d后,加入MTT溶液。繼續培養4 h后,加入DMSO融解紫色結晶產物,用分光光度計于570 nm處讀取吸收光值。
1.2.6 克隆形成實驗檢測細胞克隆形成能力:以每孔1 000個細胞接種于6孔板,設3個復孔,每3天更換一次新鮮培養基,細胞在完全培養基中培養10~12 d,甲醛固定后用0.5%結晶紫溶液對細胞進行染色,拍照并計算克隆形成數目。
1.2.7 流式細胞術檢測細胞周期:取1×105個細胞接種于6孔板,次日用慢病毒感染,置于孵箱培養96 h后離心收集細胞。細胞沉淀用PBS洗滌2次后,用冰冷的70%乙醇固定,-20 ℃存儲過夜。離心并用PBS洗滌后,細胞沉淀重懸于500 mL含50 mg/mL 碘化丙啶,0.1 mg/mL RNase A,0.05% Triton X-100的PBS中,室溫下避光孵育15 min。細胞周期用Becton Dickinson FACS Caliburr進行檢測。
1.2.8 Transwell實驗檢測細胞遷移:將含5×104個肺腺癌細胞A549的100 μL無血清培養基加入上室,下室加入600 μL含10%胎牛血清的培養基,放置培養箱中孵育。24 h后,用甲醇固定細胞,0.1%結晶紫染色,棉簽拭去上室細胞,顯微鏡下隨機選擇5個區域用于細胞計數。
1.3 統計學處理方法 采用SPSS17.0軟件進行分析。計量資料用±s表示,兩組間比較采用t檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 AKAP9對肺腺癌細胞A549增殖的影響 利用針對兩個不同靶點的shRNA感染肺腺癌細胞A549,RTqPCR結果顯示,shAKAP9 1#組(0.221±0.043)和shAKAP9 2#組(0.331±0.034)AKAP9 mRNA的表達水平與shNC組(1.00±0.049)比較顯著降低(P<0.01),見圖1A。Western blot結果顯示,相較shNC組(1.00±0.033),shAKAP9 1#組(0.387±0.037)和shAKAP9 2#組(0.52±0.025)AKAP9蛋白水平明顯下降(P<0.01),見圖1B。MTT結果顯示,從第3天開始,shAKAP9 1#組(0.383±0.048)和shAKAP9 2# 組(0.405±0.057)細胞數目較shNC組(0.765± 0.016)顯著減少(P<0.01),見圖1C。克隆形成實驗結果顯示,相較shNC組(190.667±26.652),shAKAP9 1#組(14.333±6.658)和shAKAP9 2#組(50.333±4.163)克隆形成能力明顯減弱(P<0.01),見圖1D、圖1E。
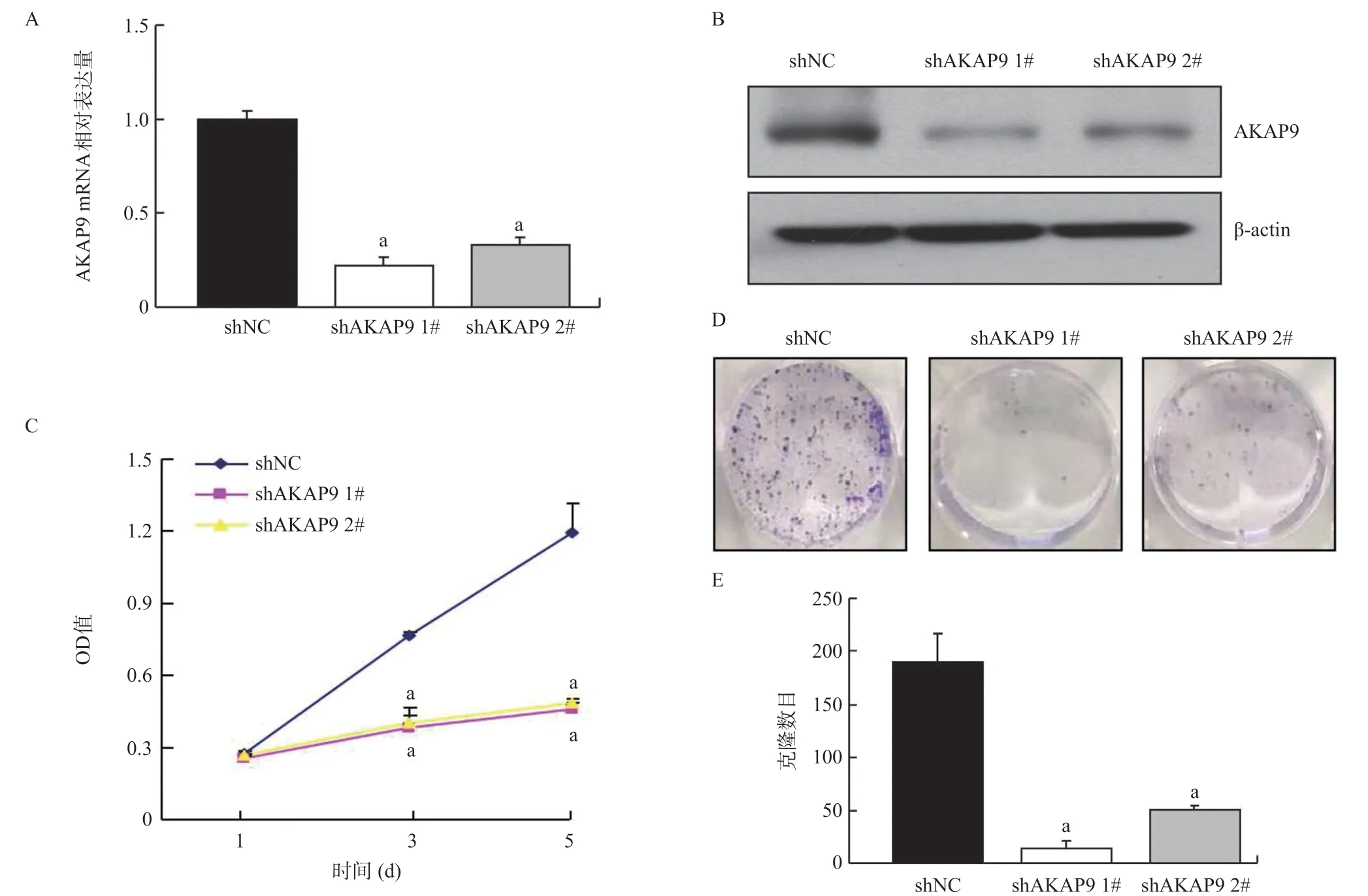
圖1 沉默AKAP9對肺腺癌細胞A549增殖的影響
2.2 AKAP9對肺腺癌細胞A549遷移的影響 Transwell實驗結果顯示,相較shNC組(319.8± 22.039),shAKAP9 1#組(123.8±14.618)和shAKAP9 2#組(164.2±9.338)遷移細胞數目減少(P<0.01),見圖2。
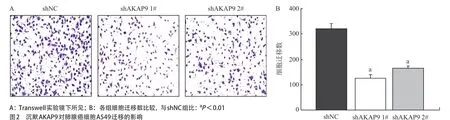
圖2 沉默AKAP9對肺腺癌細胞A549遷移的影響
2.3 AKAP9對肺腺癌細胞A549細胞周期的影響 流式細胞術結果顯示,相較shNC組(59.7±0.854)%,shAKAP9 1#組(70.2±0.557)%和shAKAP9 2#組(65.9±0.625)%細胞停滯在G1期顯著增加(P<0.01),見圖3A、圖3B。同時,細胞周期負性調控基因P27表達增加,正性調控基因Cyclin D1和CDK4表達減少,見圖3C、圖3D。
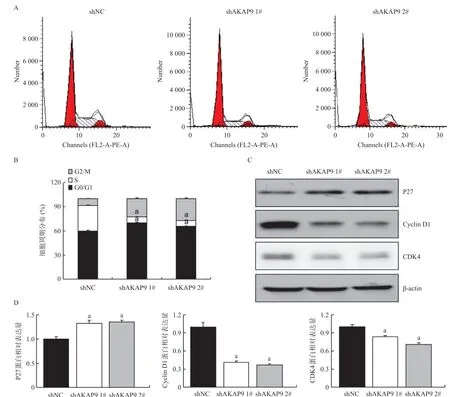
圖3 沉默AKAP9對A549細胞周期的影響
2.4 AKAP9對ERK通路的影響 Western blot結果顯示,相較shNC組,沉默AKAP9組EGFR和其下游基因ERK的磷酸化水平明顯降低(見圖4)。
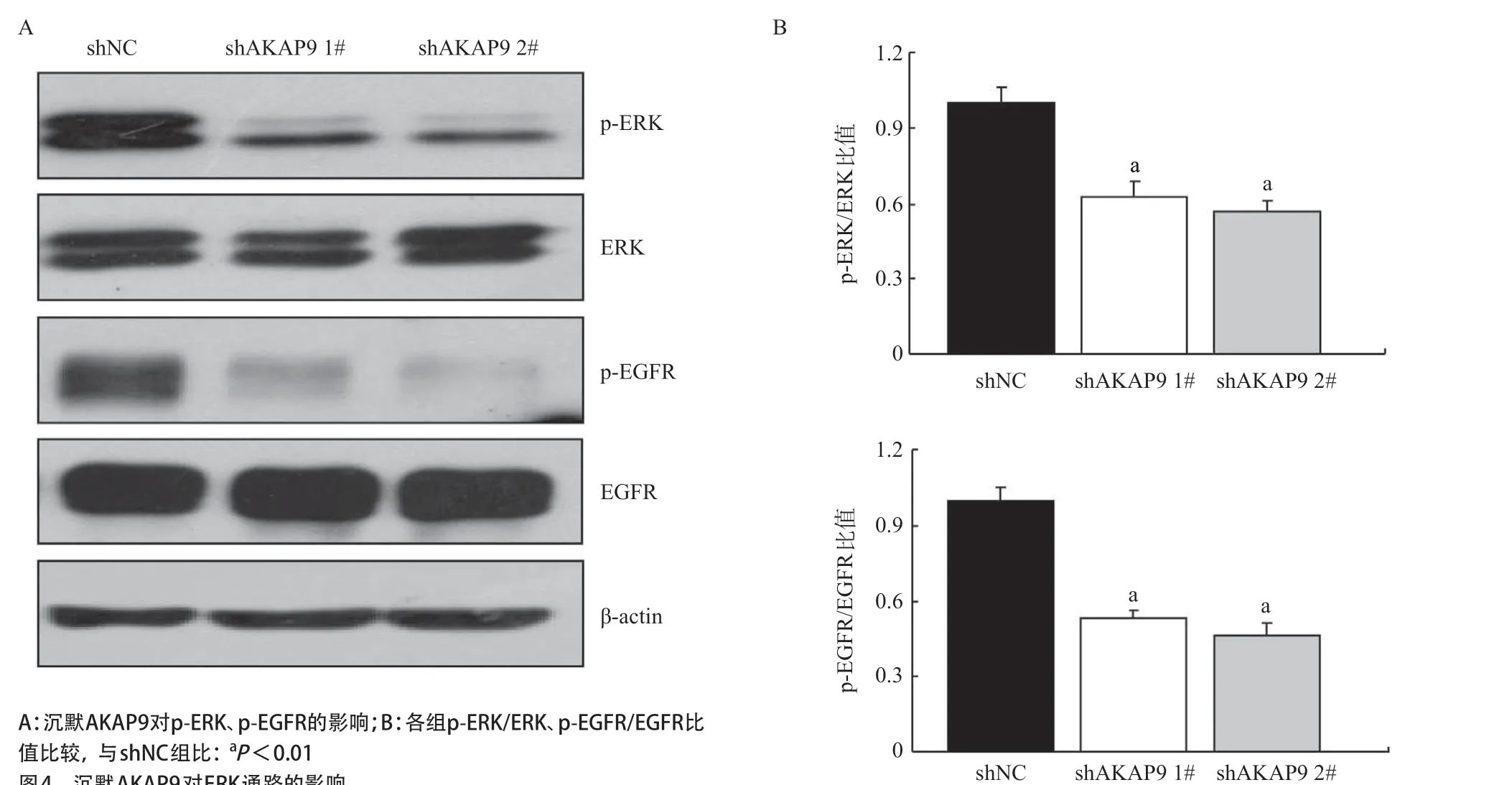
圖4 沉默AKAP9對ERK通路的影響
3 討論
AKAP9是一種蛋白激酶A(protein kinase A, PKA)錨定蛋白,它位于染色體7q21,富含coilcoil結構,可與激酶或磷酸酶相互作用,從而介導細胞內多種信號通路傳導[10]。AKAP9與CDK5RAP2、 Myomegalin、磷酸二酯酶4D、PKA等蛋白在高爾基體和中心體上形成一個復合體,來調節微管的形成和肌動蛋白的動態變化[11-13],進而參與細胞分裂、細胞分化和細胞動力等過程。研究報道,在結直腸癌細胞中,AKAP9可以與細胞分裂周期因子42相互作用蛋白4結合促進腫瘤的上皮間質轉化[14];在急性髓系白血病中,AKAP9可通過Wnt/β-catenin通路促進細胞的干性[8]。
前期研究中,我們通過Kaplan-Meier plotter 平臺分析AKAP9的表達水平與肺癌預后的關系,以AKAP9表達水平的中位值為截斷值,將1 925例肺癌患者分為AKAP9高表達組和AKAP9低表達組,Kaplan- Meier生存曲線表明AKAP9表達水平高的肺癌患者預后更差[HR=1.19(1.05~1.35),P=0.008],提示AKAP9可作為肺癌患者預后的標志物。有研究報道AKAP9與DNA合成和中心體復制有關[15],從而參與細胞周期進程。細胞周期是一個受特定細胞周期素(Cyclins)和細胞周期素依賴性激酶(CDKs)嚴格調控的步驟。在G1期,Cyclin D和CDK4/6形成復合體,磷酸化視網膜母細胞瘤蛋白(retinoblastoma protein, Rb),使細胞從G1期向S期轉換,促進細胞分裂。P27是Cyclin-CDK復合體活性的抑制劑,可以使Cyclin-CDK復合體磷酸化Rb的功能失活,從而阻斷細胞周期進程[16]。本研究發現沉默AKAP9可以通過增加P27的表達以及抑制Cyclin D1和CDK4的表達來誘導肺癌細胞G1-S期阻滯,從而抑制肺癌細胞增殖和克隆形成能力。有研究發現,敲減AKAP9可干擾中心體和非中心體的微管成核,促使高爾基體結構破碎,從而抑制T細胞的遷移[17]。本研究也發現沉默AKAP9抑制肺腺癌細胞A549的遷移能力。
ERK通路屬于絲氨酸/蘇氨酸激酶家族,是影響細胞增殖最重要的信號通路,許多關鍵的生長因子和原癌基因通過該通路發揮促進腫瘤生長的作 用[18]。異常激活ERK通路可促進肺癌細胞生長和轉移[19],而且還是肺癌細胞對藥物產生耐藥性的重要原因[20]。目前,許多針對ERK通路的抑制劑正在進行臨床試驗[19]。研究報道ERK通路激活可增加Cyclin D1表達水平[16],從而促進細胞周期進程。本研究發現,沉默AKAP9可以通過抑制ERK通路減少Cyclin D1的表達,進而誘導A549細胞G1-S期阻滯。EGFR屬于酪氨酸激酶家族,它的生物學功能是調節上皮組織發育和穩態[21]。EGFR突變在非小細胞肺癌基因突變中位居第二,尤其在亞洲非小細胞肺癌患者中其發生率高達50%[22]。異常激活EGFR可以通過傳導下游ERK、AKT、蛋白激酶C等多條通路來促進肺癌的發生發展[21]。EGFR抑制劑對不適合手術、EGFR突變的肺腺癌患者具有顯著的治療作用,能延長患者的生存期[23]。本研究發現,沉默AKAP9可以減弱肺腺癌細胞A549的EGFR和ERK活性,為研究肺腺癌細胞A549異常增殖和遷移提供了一個新的分子機制。
綜上所述,本研究發現沉默AKAP9可以通過抑制EGFR和ERK磷酸化,抑制肺腺癌細胞的增殖和遷移,并誘導G1-S期阻滯;AKAP9可能成為臨床治療肺腺癌的新靶點。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
海峽科技與產業(2016年3期)2016-05-17 04:32:12
癌變·畸變·突變(2016年3期)2016-02-27 06:15:34
醫學研究雜志(2015年12期)2015-06-10 06:57:46
