貴州雷公山國家級自然保護區原住民村落與周邊保護地鳥類多樣性差異
2022-05-24 08:09:04盛尚朱蕓劉宇鄭進鳳余永富李揚張凱徐雨
四川動物 2022年3期
盛尚, 朱蕓, 劉宇, 鄭進鳳, 2, 余永富, 李揚, 張凱, 徐雨*
(1. 貴州師范大學生命科學學院,貴陽550025; 2. 西南大學生命科學學院,重慶400715;3. 雷公山國家級自然保護區管理局,貴州雷山557199)
自然保護區是生物多樣性保護的重要基石。目前,中國已經建立2 750個保護區,占陸域國土面積的15%(https://www.forestry.gov.cn/)。但由于歷史遺留問題,許多保護區內分布有一定居民點,可能對保護區的有效管理造成一定影響(徐網谷等,2016;孫潤等,2017;吳嘉君等,2020)。2019年,《關于建立以國家公園為主體的自然保護地體系的指導意見》指出要分類有序解決保護區內仍有原住民居住等歷史遺留問題(新華社,2019)。2020年,自然資源部、國家林業和草原局發布《關于做好自然保護區范圍及功能分區優化調整前期有關工作的函》(自然資函〔2020〕71號)(羅紅等,2021),明確指出要解決好原住居民生產生活與保護管理間的矛盾與沖突。
科學實施保護區的優化調整工作,有賴于對比分析居民區對生物多樣性的影響。但是,目前研究的結果尚存一定爭議。Liu等(2001)發現保護區內人口數量及家庭規模的增多導致臥龍國家級自然保護區大熊貓棲息地喪失率與破碎率增加;鄧敏等(2020)報道壺瓶山國家級自然保護區較強人為干擾樣段中蝴蝶群落的Simpson指數、Shannon-Wiener指數、Menhinick豐富度指數均顯著低于無干擾樣段;潘晨等(2021)發現升金湖國家級自然保護區內人為干擾強度持續增大導致聚集度指數、斑塊內聚力指數下降,減弱了相同區域間的連通性。但部分具有傳統民族特色的人類活動反而促進生物多樣性的保護。最大熵模型(MaxEnt model)揭示,禁忌殺生的336座藏傳佛教寺廟能有效地保護含三江源自然保護區核心區在內的8 342 km雪豹棲息地(Li.,2013);坡崗自然保護區內各類傳統文化景觀,如祭祀點、村寨等的文化多樣性保護價值與生物多樣性保護價值(含野生珍稀動、植物多樣性、生境類型等指標)呈正相關關系,可較好地維持區域生態功能的穩定(畢興等,2020)。因此,保護區內居民區對生物多樣性的影響可能與地理區域、文化氛圍等因素有關。在開展保護區優化調整工作時,需結合保護區實際情況展開調查分析。
雷公山國家級自然保護區位于貴州省東南部苗嶺山地的雷公山,區內有25個行政村,苗族人口約占90%以上(張一帆,2017)。盡管已有研究指出苗族傳統文化對生物多樣性保護具有潛在價值(劉舜青,賴力,2003;趙棟昌,劉鋒,2020),但目前并不清楚原住民村落對生物多樣性的保護效益。鳥類是生態系統的重要組成部分及指示物種,能在一定程度上反映棲息地生境的質量(Padoa-Schioppa.,2006;彭子嘉等,2020),且其多樣性水平可作為整個生態系統健康狀況的指標(Roth & Weber,2008)。為此,本研究對保護區原住民村落及其周邊保護地的鳥類多樣性差異進行研究,為保護區實施優化調整及科學管理提供理論依據,同時向其他類似自然保護區制定管理對策提供參考。
1 研究方法
1.1 研究區概況
雷公山國家級自然保護區位于貴州省東南部,屬長江水系與珠江水系分水嶺,地理坐標108°5′~108°24′E,26°15′~26°32′N,海拔650~2 178.8 m(張鳳太,蓋媛瑾,2012)。地形高聳挺拔,地勢呈西北高、東南低,屬中亞熱帶季風山地濕潤氣候區,年均降水量1 300~1 600 mm(王英等,2016)。區內分布有35種國家一、二級重點保護野生動物和 25種國家一、二級重點保護野生植物(http://lgsbhq.qdn.gov.cn/)。區內植被類型有20多個,是以保護禿杉等珍稀瀕危物種為主的森林生態系統類型自然保護區(貴州省黔東南苗族侗族自治州人民政府,1989)。
在保護區內選擇7個苗族原住民村落(平祥村、雀鳥村、陡寨村、水寨村、提香村、桃香村和格頭村)及其周邊保護地作為研究地點。這些村落交通較為閉塞、經濟發展較差(張鳳太,蓋媛瑾,2012;蓋媛瑾,2013),產業結構主要由養殖業(以養豬、雞、鴨等為主)、種植業(以種植水稻、玉米等為主)和旅游業組成(余志彪,2006)。村落內保存有受苗族傳統文化保護、年代較為久遠的“神樹”(比如,古老而高大的禿杉)和“風水林”等圣境區域;且族內傳統習慣法約束著村民對森林濫砍濫伐的行為(薛達元,2014)。
1.2 調查方法
在研究中,規定原住民村落是以人口聚集居住地頭端至末端的距離為直徑的圓形區域;在圓形區域外選擇與原住民村落具有相似海拔和地形特征的區域作為周邊保護地。采用樣點法(Bibby.,1998)調查村落和周邊保護地的鳥類多樣性。共設置鳥類調查樣點208個:村落44個,周邊保護地 164個;村落與周邊保護地內樣點間的直線距離100~10 000 m。
野外鳥類調查時間為2018年7月和2019年 8月,選擇無雨、無霧的晴好天氣進行,每日調查時間為鳥類日活動相對比較頻繁的 07∶00—11∶00 和 15∶00—19∶00。調查隊伍一般由2人組成。在每個樣點,以調查人員為中心,使用10×42雙筒望遠鏡觀察樣點半徑50 m范圍內的鳥類(為減少人為干擾,觀察時間為到達2 min后至10 min內),并根據《中國鳥類野外手冊》(約翰·馬敬能等,2000)、《中國香港及華南鳥類野外手冊》(尹璉等,2017)鑒定物種,記錄鳥類數量。為避免重復記錄,相鄰樣點間的直線距離通常為 200~300 m。
1.3 數據分析
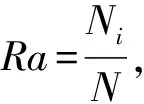
從樣點尺度與景觀尺度對比分析村落內及其周邊保護地內鳥類物種多樣性差異。在樣點尺度上,統計每個樣點上的鳥類物種豐富度、Shannon-Wiener指數()、Chao1指數()。其中,物種豐富度是統計一個樣點或群落中物種的數量(牛翠娟等,2015);Shannon-Wiener指數可描述個體出現的紊亂與不確定性,不確定性越高時多樣性越高(錢迎倩,馬克平,1994);Chao1指數對稀有物種敏感,可估算群落物種總數,值越高代表群落鳥類物種越豐富(Chiu.,2014)。采用Wilcoxcon秩和檢驗(Rosner.,2003)確定村落內及其周邊保護地間鳥類物種多樣性差異情況。
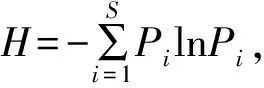
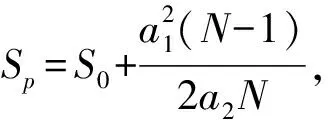
在景觀尺度上,使用物種累積曲線分析村落與其周邊保護地鳥類物種多樣性差異。若二者物種累積曲線95%置信區間重疊,說明多樣性沒有顯著差異;反之,則具有顯著差異。
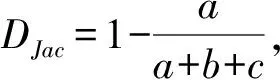
統計分析在R.4.0.3中執行;使用vegan(Oksanen.,2020)插件完成物種多樣性指數、物種累積曲線、相異性指數分析;使用ggpubr(Kassambara,2020)插件完成顯著性檢驗;使用ggplot2(Wickham.,2020)插件繪制物種豐富度、Shannon-Wiener指數、Chao1指數與物種累積曲線圖。
2 研究結果
2.1 物種組成
2次調查共記錄到鳥類64種,隸屬于7目 30科,其中鴉科Corvidae和鹟科Muscicapidae占比最高(7.8%)。從居留型來看,留鳥48種(75%)、夏候鳥11種(17.2%)、冬候鳥2種(3.1%)、旅鳥3種(4.7%);從區系上看,東洋界物種占67.2%、古北界物種占21.9%、廣布種占10.9%。中國特有鳥類 2種:灰胸竹雞和黃腹山雀。
原住民村落內共調查到鳥類33種,主要由雉科Phasianidae、雨燕科Apodidae、翠鳥科Alcedinidae、伯勞科Laniidae、鴉科、山雀科Paridae、燕科Hirundinidae、鵯科Pycnonotidae、柳鶯科Phylloscopidae、林鹛科Timaliidae、噪鹛科Leiothrichidae、鹟科、雀科Passeridae和鹡鸰科Motacillidae等19科組成。優勢種6種,分別是金腰燕、黃臀鵯、白腰文鳥、山麻雀、麻雀和灰鹡鸰,常見種16種,少見種或偶見種11種。周邊保護地內共調查到鳥類 56種,主要由鷹科Accipitridae、鴟鸮科Strigidae、佛法僧科Coraciidae、翠鳥科、擬啄木鳥科Capitonidae、啄木鳥科Picidae、山椒鳥科Campephagidae、鴉科、玉鹟科Stenostiridae、扇尾鶯科Cisticolidae、林鹛科、幽鹛科Pellorneidae、鹟科、啄花鳥科Dicaeidae、花蜜鳥科Nectariniidae、梅花雀科Estrildidae、燕雀科Fringillidae和鹀科Emberizidae等29科組成。優勢種4種,分別是煙腹毛腳燕、領雀嘴鵯、黃臀鵯和栗耳鳳鹛,常見種34種,少見種或偶見種18種。
2.2 物種多樣性
在樣點尺度上,原住民村落與周邊保護地的鳥類物種豐富度分別為3.11和1.51;Shannon-Wiener指數分別為0.91和0.36;Chao1指數分別為3.92和1.79(圖1)。
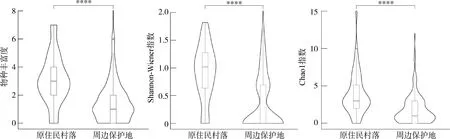
圖1 雷公山國家級自然保護區原住民村落與周邊保護地鳥類多樣性指數差異Fig. 1 Differences of bird diversity indexes between indigenous communities and the surrounding protected areas in the Leigongshan National Nature Reserve, Guizhou
在景觀尺度上,原住民村落與周邊保護地的物種累積曲線皆隨著樣點數的增加而呈上升趨勢。兩者95%置信區間有較大重疊區域,表明在景觀尺度上原住民村落與周邊保護地間的鳥類物種多樣性無顯著差異(圖2)。
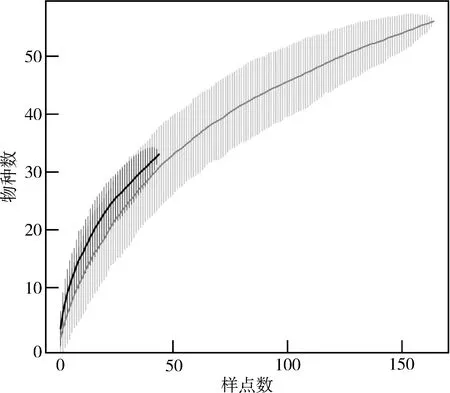
圖2 貴州雷公山國家級自然保護區原住民村落與周邊保護地鳥類物種累積曲線Fig. 2 Bird species accumulation curves in indigenous communities and the surrounding protected areas in the Leigongshan National Nature Reserve, Guizhou
2.3 群落相異性
原住民村落與周邊保護地僅有25種鳥類相同,如普通翠鳥、大山雀、金腰燕、黃臀鵯、灰鹡鸰等。Jaccard距離、Bray-Curtis距離分別為0.61、0.54(表1),表明原住民村落與周邊保護地鳥類群落間物種組成具有一定差異。
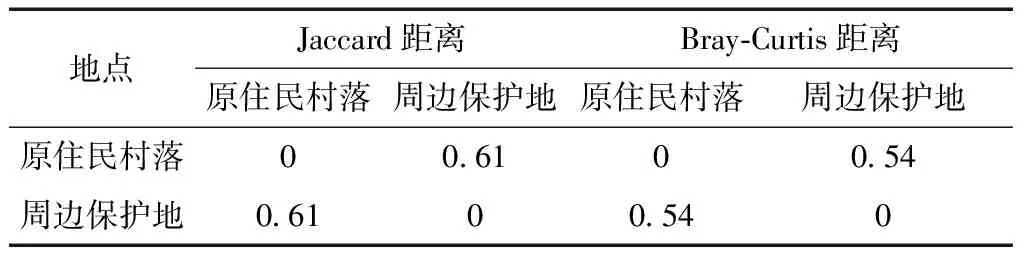
表1 雷公山國家級自然保護區原住民村落與周邊保護地鳥類群落間相異性指數Table 1 Dissimilarity indexes of bird communities in indigenous communities and the surrounding protected areas in the Leigongshan National Nature Reserve, Guizhou
3 討論
目前,已有不少學者以自然保護地或保護區為對象,對區內居民區與周邊保護地的動物多樣性開展了對比分析,但研究結果不盡相同(符建榮等,2009;那順得力格爾等,2011)。為了豐富該方面的研究內容,本研究對雷公山國家級自然保護區原住民村落與周邊保護地的鳥類多樣性進行了對比分析,結果表明村落的鳥類物種多樣性在樣點尺度上顯著高于周邊保護地,但兩者在景觀尺度上的物種累積結果無顯著差異。
在生物進化過程中,鳥類往往將人類視為潛在天敵(韋福民等,2007;唐慶圓,2009),對人類活動比較敏感(Frid & Dill,2002;Blumstein.,2005),人類聚居地或人類活動頻繁區域的鳥類多樣性通常較低(劉吉平等,2008;劉子成等,2012)。但在一些村落內,農業環境可為鳥類提供豐富的食物資源與良好的棲息場所(S?derstr?m.,2001;Tschamtke.,2005),吸引一些性情勇敢的鳥類前來覓食和繁殖(Jeliazkov.,2016;盧訓令等,2019)。除此之外,裴盛基和龍春林(2008)、薛達元(2014)等提出,某些有傳統民族特色的人類活動可能對生物多樣性的保護有積極作用。本研究涉及的村落中苗族傳統文化保存比較完好,村民受苗族習慣法的約束禁止對森林濫砍濫伐(薛達元,2014),且村落內保存有年代較為久遠的“神樹”(如古老而高大的禿杉)和“風水林”等圣境區域,可能為特定鳥類提供了適宜的棲息地(Bhagwat.,2005),使樣點上的鳥類多樣性較高。
但是,原住民村落與周邊保護地的鳥類物種多樣性在景觀尺度上無顯著差異,該結果可能與景觀異質性有關。生境異質性是物種數量的重要決定因素(Boecklen,1986),多類型植被與復雜結構可吸引不同鳥類集團并增加物種多樣性(張征愷,黃甘霖,2018)。此外,植物豐富度(彭子嘉等,2020)、樹木多樣性(武欣慧等,2016)也可能影響鳥類多樣性。本研究發現在原住民村落區域內除分布有部分苗族傳統文化中視為“神樹”的禿杉(王云,2005)外,以人工林和農田為主,植被類型相對單一;村落附近農田內農作物以小麥、玉米等單一經濟作物為主。生境內僅栽種指定樹木與經濟作物,能吸引麻雀、山麻雀、黃臀鵯、灰胸竹雞、栗耳鹀等“伴人鳥類”,但并不能滿足多種鳥類的取食與營巢(龔浩林等,2019)。村落內單一的生境或同質化的景觀可能使樣點間的物種組成相似度較高,導致村落與周邊保護地在景觀尺度上的鳥類物種多樣性無顯著差異。
進一步分析揭示,原住民村落與周邊保護地的鳥類物種組成具有一定差異。這可能與村落、周邊保護地景觀特征、植被組成、人類干擾強度不同有關。不同的景觀或生境間可能存在各自所特有的鳥類(Pithon.,2015)。對波蘭南部地區教堂和周邊農場的鳥類群落研究指出,兩者因樹木數量、建筑數量、建筑高度等組成特征不同導致鳥類物種組成不同(Skórka.,2018)。村落內農田生境可能吸引一些伴人鳥類,苗寨建筑與古樹可能為偏好該棲息地類型的某些鳥類提供避難所。相反,周邊保護地內較弱人為干擾、河流、多種植被類型等組合的環境可能吸引偏好安靜、復雜生境的鳥類,如紅嘴相思鳥、藍翅希鹛、藍喉太陽鳥等。
綜上所述,保護區原住民村落的鳥類物種多樣性在樣點尺度上高于周邊保護地,但在景觀水平上兩者鳥類物種多樣性無明顯差異。兩者在鳥類群落組成有一定差異。本研究認為村落內農田和傳統文化所營造出的特殊環境可能有助于一些鳥類的生存和繁殖,但同質化的景觀不能有效促進鳥類多樣性。因此,建議保護區合理考慮原住民村落對生物多樣性的影響,重視農業景觀中不同類型植被的配置,改善因人類活動產生的生境結構單一、景觀同質化等情況。對于未來工作,建議從植被群落結構等方面開展分析,以進一步揭示生境特征與鳥類多樣性間的關系。
感謝貴州師范大學的唐蓉、賀霜、邰合、冉小東、龍盛堯等同學在野外調查工作中所付出的努力。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
音樂探索(2022年2期)2022-05-30 21:01:37
現代裝飾(2021年6期)2021-12-31 05:27:54
小學科學(學生版)(2020年12期)2021-01-08 09:28:10
少年漫畫(藝術創想)(2020年12期)2020-06-09 05:50:08
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
