西藏林芝市西藏蟾蜍種群的年齡、生長及其他種群特征的研究
2022-05-24 08:09:04李云磊王昊天陸宇燕李丕鵬
四川動物 2022年3期
李云磊, 王昊天, 陸宇燕, 李丕鵬
(沈陽師范大學兩棲爬行動物研究所,沈陽110034)
種群的年齡結構主要是指種群中不同年齡組個體的組成情況,年齡結構的類型主要分為增長型、穩定型和衰退型(Guarino & Erismis,2008)。年齡結構不僅是種群生態學研究中的核心問題,對于兩棲動物生理、生態甚至形態分類方面的研究均具有重要意義(Hasumi,2010)。
兩棲類動物年齡推斷的常見方法有體長計數法(Voris & Jayne,1979)、骨齡學方法(易祖盛等,2000)、標志重捕法(Wagner.,2011)、精巢分葉法(Sinsch,2015)等。目前,公認可靠、有效且得以廣泛使用的是骨齡學方法。
兩棲類動物作為變溫動物,具有冬眠的習性,其代謝功能隨季節變化。這使其骨骼生長發育以及礦物質沉積在不同季節表現出明顯的差異,即在生活期和冬眠期形成的骨質環密度不同。由于物質沉積量的差異,第一年冬眠期及其前后生長緩慢階段形成的骨環(染色較深)和第二年生活期形成的骨環(染色較淺)間呈現出1個明暗交界的同心環帶(畢遠閣等,2009),被稱為生長抑制線(line of arrested growth,LAG),LAG的數目可間接判斷出個體年齡(易祖盛等,2000)。李文蓉等(2010)發現,動物的指/趾骨與其他骨塊一樣具有良好且清晰的LAG結構,同時剪指/趾可避免對動物的殺戮,對動物保護具有積極意義。該法在近10種兩棲類中被用于種群年齡結構的鑒定和研究(Hemelaar,1985,1988;Bastien & Leclair,1992;Beebee & Denton,1993;Khonsue.,2000;Kumbar & Pancharatna,2002;李斌,2004;Liang & Lu,2006;毛敏等,2011;Bionda.,2018)。
西藏蟾蜍屬兩棲綱Amphibia無尾目Anura蟾蜍科Bufonidae蟾蜍屬,主要分布在海拔2 400~4 300 m較潮濕的高原草地或山區。目前對西藏蟾蜍的研究集中在消化系統(曾麟等,2011)、皮膚顯微結構和基礎組織學(高鳳娟等,2016),而其種群的年齡結構未見報道。本研究以骨齡學方法,輔以生殖腺及組織學結構的觀察,對分布于西藏自治區林芝市的一個西藏蟾蜍種群的年齡結構、體長、生殖腺形態和性成熟年齡等進行了探討,旨在積累更多生物學資料,為今后該物種的保護工作奠定基礎。
1 研究材料與方法
1.1 實驗材料
西藏蟾蜍為沈陽師范大學生命科學學院兩棲爬行動物標本室福爾馬林固定標本,共計63只(♀29只,34只),采自西藏自治區林芝市(94°56′~94°58′E,29°14′~29°17′N,海拔2 900 m),采集時間主要為2014年、2015年7月28日—8月 2日(當地西藏蟾蜍的繁殖季節)。
1.2 實驗方法
標本流水沖洗過夜,利用賽多利斯電子天平(Mettler Toledo Q/OAFA02-2009,上海精密科學儀器有限公司天平儀器廠)稱量體質量(精度0.001 g),稱量前利用吸水紙吸干標本體表液體。利用電子數顯游標卡尺(哈爾濱量具廠,GB/T 14899-94)測量體長(吻端到泄殖腔距離,SVL;精度0.01 mm)。成年個體依據是否有婚墊區分性別,對婚墊不明顯個體則解剖辨別生殖腺確定性別,生殖腺稱重(精度0.001 g)。
取第三指骨和生殖腺流水沖洗過夜(指骨先在5%硝酸溶液中脫鈣,直至軟化),常規石蠟切片,指骨切片厚度10 μm,生殖腺切片厚度7 μm,HE染色。利用Olympus BX-51顯微鏡觀察,Olympus DP70拍照。
1.3 骨齡計數標準
為了保證年齡估計的準確性,取同一指骨不同位置的8~10個切片進行LAG計數,由于所用標本均在繁殖期前后捕獲,故當年的LAG還沒有完全形成,因此在計數過程中將骨組織切片的最外緣(當年生活期形成的骨質環)也算作1條LAG(馬小艷,2009)。為了降低誤差,由2位獨立觀察者分別進行LAG數目的統計。
1.4 LAG間距離測量
利用Image-Pro Plus 6.0測量相鄰2條LAG之間的距離(Hemelaar,1985;易祖盛等,2000),同一指骨隨機選取8~10個不同部位進行測量:A~G分別代表1~2、2~3、3~4、4~5、5~6、6~7、7~8條 LAG之間的距離。
1.5 數據處理
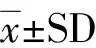
1.6 計算公式
使用von Bertalanffy模型SVL=SVL(1-e-+b)建立種群生長曲線,估算西藏蟾蜍的生長模式(Hemelaar,1986),生長速率=dd=(S-S)(Lou.,2012)比較雌、雄性生長的快慢,其中,SVL為年齡時的體長,SVL為漸進最大體長,為生長系數,b是常數,S是一個種群成體的最大體長,S是一個種群成體的平均體長。采用生殖腺系數=生殖腺質量/體質量×100%(劉海映等,2013)分析各個年齡段雌、雄性的生殖腺發育狀況。
2 實驗結果
2.1 年齡鑒定及種群年齡結構
第三指骨切片上可觀察到:中心空腔為骨髓腔,向外依次是骨質疏松的內骨板,大多呈裂開縫隙狀,為Kastschenko線(指內骨板與外骨板之間有 1層缺乏骨組織細胞的、均質的軟骨樣組織,由于制片易裂開形成縫隙),骨質致密為外骨板(圖1)。在外骨板上可見清晰的LAG,每一條代表經過了 1個冬眠期,即為1齡。由于西藏蟾蜍當年完成變態,外骨板上可見LAG數目即為個體年齡。

圖1 西藏林芝西藏蟾蜍指骨組織切片Fig. 1 Phalangeal section of Bufo tibetanus in Nyingchi City, Tibet
骨齡學方法可將西藏蟾蜍劃分為9個年齡組,其中,雌性的最長壽命為9齡,雄性為8齡;兩性的2齡組均為個體數目最多的年齡組,其次為 3齡組、4齡組。雌性的平均年齡為(3.59±2.23)年,雄性為(3.56±1.83)年,兩性在各年齡組的數量分布上的差異不顯著(=63,=0.054,>0.05)(圖2)。隨著年齡增加,個體數逐漸減少,種群屬于增長型。

圖2 西藏林芝一個西藏蟾蜍種群的年齡結構分布Fig. 2 Age structure of one Bufo tibetanus population in Nyingchi City, Tibet
2.2 體長與年齡
西藏蟾蜍雌性的平均體長為46.44 mm±15.91 mm(23.96~70.61 mm),雄性為45.06 mm±13.00 mm(23.98~64.66 mm)。各年齡組雌、雄性的平均體長基本接近(表1),無顯著差異(﹥0.05)。其中,5~9齡組由于個體數量明顯減少,甚至一個年齡組中僅有1只個體。這雖然證實了西藏蟾蜍在骨齡學上年齡組的連續性,但由于數量不符合統計學要求,后續分析中這部分數據僅作為參考。

表1 西藏林芝一個西藏蟾蜍種群雌雄間體長的差異統計Table 1 Statistics of body length between male and female individuals of one Bufo tibetanus population in Nyingchi City, Tibet
雌、雄性個體的年齡與體長均呈極顯著正相關(Spearman相關性檢驗:雌性:=0.900,=29,<0.01;雄性:=0.878,=34,<0.01)。模擬兩性的生長模式并建立生長曲線(圖3):雌性為SVL=74.542 0(1-e-0.300 4-0.176 1),雄性的為SVL=63.210 3(1-e-0.377 5-0.135 4);兩性體長均隨年齡增長而呈現增長的趨勢,但雌性的生長速率(3.29)大于雄性(2.65)。5齡前雌、雄性的生長曲線均呈快速上升趨勢,5齡后趨于平緩,且雄性的上升趨勢明顯低于雌性。

圖3 西藏林芝市西藏蟾蜍生長曲線Fig. 3 Growth curve of Bufo tibetanus by von Bertalanffy model in Nyingchi City, Tibet
2.3 性成熟年齡
雌性個體依據卵子外部形態可劃分為3類:體長62.70 mm以上為性成熟個體,體腔中可見發育良好的卵巢,左右成對,體積較大,表面可見大量黑色卵粒,為尚未進行繁殖的成熟雌性個體,還有部分個體的卵巢體積較小,卵巢呈淡黃色和黑色2個部分,在黑色部分可見飽滿的黑色卵粒,淡黃色部分未見大量成熟黑色卵粒,推測這些是已經完成了繁殖過程,排出了大部分成熟卵粒的個體;體長48.82~61.40 mm為亞成體,卵巢呈淡黃色,表面可見淡黃色顆粒結構;體長48.82 mm以下為幼體,卵巢呈淡黃色,未見明顯卵粒(圖4:A,B,C)。雄性個體均可見成對精巢,體長42.40 mm以上為性成熟個體,精巢體積較大,組織切片中均可見形態較為規則的生精小管;體長 35.17 mm以下為亞成體,精巢尚未完全發育(圖4:D,E,F)。

圖4 西藏林芝一個西藏蟾蜍種群的雌、雄性生殖腺Fig. 4 Male and female gonads of one Bufo tibetanus population in Nyingchi City, Tibet
雄性生殖腺組織切片結果顯示,1齡(圖5:A)精巢中可見形態相異、胞核大小及染色不同的大量細胞,排列松散,且未見成型的生精小管;2齡(圖5:B)精巢中可見各類細胞明顯變大,有不太明顯的區域劃分,排列呈區域性的密集;3齡(圖5:C)精巢中可見明顯的管狀結構,即已形成的生精小管,同時可見成型的精子束,但排列不是很整齊密集(圖5:D);4齡(圖5:E,F)精巢中可見較多含有精子的生精小管,其中可見處于不同發育階段的生精細胞和已經成型的精子;5齡(圖5:G)精巢中可見明顯的生精小管、界膜和間質細胞等;6齡(圖5:H)精巢中生精小管管腔明顯增大,未見正常的精子。

圖5 西藏林芝一個西藏蟾蜍種群雄性不同年齡生殖腺切片Fig. 5 Male gonad sections of different ages of one Bufo tibetanus population in Nyingchi City, Tibet
C明顯小于A和B,且差異顯著,但與D~G之間無顯著差異。全部個體的 B、C之間的差異顯著,少量個體(♀4只,4只)的A、B之間的差異顯著(表2)。這種現象提示:骨骼的生長存在著不均衡性,骨質環厚度在發育初期(3齡,完成變態后的第2~4個冬眠期)的增長較快,而在 4齡(性成熟年齡)以及之后的增長明顯減緩,但也存在個體差異。

表2 西藏林芝一個西藏蟾蜍種群性成熟個體生長抑制線間距離顯著性分析Table 2 Significance analysis of the distance between arrested growth lines in sexually mature individuals of one Bufo tibetanus population in Nyingchi City, Tibet
雌雄兩性個體生殖腺系數均表現為3齡組和4齡組之間具有顯著性差異,而其他年齡組均無顯著性差異(表3)。

表3 各個年齡段雌雄西藏蟾蜍生殖腺系數差異性比較Table 3 Comparison of male and female gonad coefficient of Bufo tibetanus at different age groups
3 討論
3.1 骨齡學方法
骨齡學方法是目前鑒定兩棲爬行類動物年齡廣泛使用的一種方法(Vences & Jovanovic,2010;Hjernquist.,2012),但是也存在著一些干擾因素。最明顯的是雙重線,有些兩棲動物以夏眠的方式度過酷熱缺水的夏季,此時在長骨切面上將出現2條清晰而非常靠近的類似LAG的結構,即雙重線(Liang & Lu,2006)。其次是假線,又稱附加線,是由于個體受傷、或者遇到短暫的氣候劇烈變化或者食物缺乏在長骨切面上的反映,一般假線僅在局部出現,未見其形成閉合環狀結構,且線條結構不清晰(Castanet & Smirina,1990)。雙重線和假線在一定程度上均會干擾對個體年齡的準確計數,出現高估或者低估年齡的可能(Leclair & Castanet,1987)。第三是由于骨內膜的重吸收導致1齡線被局部損壞的現象,但很少涉及到2齡線(Hemelaar,1980)。在長骨切片結構完整時,并不會影響LAG計數。
本研究中有5%的個體指骨切片存在雙重線現象,僅1只個體指骨切片上觀察到骨內膜的重吸收現象,但最內圈1齡線并未被完全吸收,可以清晰識別。假線或LAG模糊不清等現象均未觀察到。因此,鑒定生活于高海拔地區的西藏蟾蜍種群年齡時,骨齡學方法是有效、可靠的。
3.2 西藏蟾蜍的性成熟年齡

兩棲動物達到性成熟年齡的時間是影響兩棲類動物種群生態學的重要因素之一,而決定性因素是動物的生長速率及性成熟時的個體大小(Ryser,1996)。研究顯示高海拔地區的物種性成熟年齡高于低海拔地區(Brockleman,1975)。Licht(1975)研究的西部多斑蛙種群中,低海拔區域的雌性1~2年即可達到性成熟,高海拔區域的雌性則需要5~6年。在對華西雨蛙川西亞種(Liao & Lu,2010a)雄性個體和岷山蟾蜍(王冬麗,2017)的研究中呈現了相同的規律。但也有物種性成熟年齡與海拔變化沒有明顯的相關性,Liao等(2011)發現中國西南部不同海拔(310 m和 880 m)的澤蛙性成熟年齡均為 1齡,不隨海拔變化而變化。而西藏蟾蜍性成熟年齡(4齡)符合高海拔物種需要更長時間的生長以達到滿足生殖的最小體長的研究結果。
Morrison等(2004)研究發現,所有澳大利亞昆士蘭州的紅眼雨濱蛙種群繁殖年限(2年)與生活區域海拔(110 m/800 m)、性成熟年齡(2齡/4齡)無關,這表明盡管很多物種的性成熟年齡與海拔有關,但其潛在繁殖壽命并沒有顯著變化。在對西藏蟾蜍精巢生精小管的觀察中也發現,在初步性成熟的3齡,性成熟的4齡和5齡個體中,均存在著可見的成熟精子,但在6齡個體中未見成型的精子。由于個體數量的限制,還不足以判斷西藏蟾蜍的潛在繁殖壽命是否僅有2年,需要進一步積累資料。
3.3 西藏蟾蜍壽命
分布于不同海拔的兩棲類在壽命上有明顯的差別,表現為高海拔個體壽命長于低海拔個體(Elmberg,1991)。分布于低海拔地區的棕背蛙個體年齡為 7齡,而高海拔地區個體最大年齡可達11齡(Lai.,2005)。在對華西雨蛙(Liao & Lu,2010a)和岷山蟾蜍(陳建斌,2016)的研究中也顯示了同樣的規律。在對不同岷山蟾蜍種群研究時還發現,雌性壽命隨著海拔的升高而變長,而雄性的壽命不受海拔的影響(王冬麗,2017)。在兩棲動物中,雌性個體的壽命比雄性個體長的現象普遍存在,例如雌性瓦拉鋤足蟾最長壽命為10齡,而雄性為7齡(Aadreone.,2011);雌性四川湍蛙最長壽命為9齡,而雄性為7齡(Liao & Lu,2010b)。本研究中,雌性西藏蟾蜍最長壽命為9齡,雄性為8齡,雌性的壽命長于雄性。可見西藏蟾蜍屬于高壽群體,但由于樣本量的限制,有待后續的積累。
3.4 年齡結構
年齡結構作為種群生態學研究的重點問題,學者們進行了較為廣泛的研究,積累了豐富的資料,研究結果顯示秦嶺雨蛙(任楨,2015)、隆肛蛙和倭蛙(鮮盼盼,2017)等的年齡結構屬于增長型,花背蟾蜍、黑斑側褶蛙、合征姬蛙屬于穩定型,而分布在太白山地區的中國林蛙則為衰退型(鮮盼盼,2017)。本研究所涉及的林芝市西藏蟾蜍種群在被采集的2014—2015年的年齡結構屬于增長型,說明該種群具有健康發展潛能。本研究結果豐富了西藏蟾蜍部分種群特征的生物學信息,為進一步全面深入了解西藏蟾蜍提供了可靠的依據。
