綠萼鳳仙花對低溫脅迫的生理響應
2022-04-27 12:59:04劉松曹孟會李芹梅張天謠李文祥黃美娟黃海泉
山東農業科學 2022年3期
劉松 曹孟會 李芹梅 張天謠 李文祥 黃美娟 黃海泉
摘要:為探析綠萼鳳仙花對低溫脅迫的生理適應性,本研究在前期預試驗基礎上,以20℃為對照,分別在2℃和0℃處理0、12、24、48、72h,測定其細胞傷害率(CIR)、葉綠素和丙二醛(MDA)含量、過氧化氫酶(CAT)和超氧化物歧化酶(SOD)活性及脯氨酸(PRO)等滲透調節物質的含量等指標,并進行相關性分析,以期為綠萼鳳仙花開發應用提供參考。結果表明,隨溫度的降低及脅迫時間的延長,低溫脅迫對綠萼鳳仙花的傷害程度不斷加劇。2℃和0℃低溫脅迫下,綠萼鳳仙花的CIR及MDA、可溶性糖(SS)和可溶性蛋白(SP)含量均呈上升趨勢,在脅迫72h時分別高于對照3.18倍和5.09倍、80.85%和2.31倍、1.02倍和1.24倍、2.39倍和4.93倍,差異極顯著;POD活性、葉綠素含量呈下降趨勢,在脅迫72h時分別顯著低于對照43%和71%、53%和60%,差異極顯著;SOD、CAT活性及PRO含量均隨脅迫時間的延長呈先升高后降低趨勢,溫度越低,越快達到峰值。相關性分析結果表明,CIR、葉綠素含量、MDA含量、SP含量和POD活性可作為綠萼鳳仙花耐寒性鑒定的主要指標,SOD和CAT活性、PRO和SS含量可作為輔助指標。綜合來看,綠萼鳳仙花對2℃低溫具有一定的耐性,在0℃低溫下生長受抑并表現出一定的寒害特征。本研究結果可為綠萼鳳仙花的引種馴化、雜交育種及抗寒育種提供一定的理論依據。
關鍵詞:綠萼鳳仙花;低溫脅迫;細胞傷害率;抗氧化生理指標;滲透調節物質;相關性分析
隨著全球氣候變化加劇,極端天氣如極端高溫、極端低溫等氣候事件頻繁發生[1]。而溫度是決定植物地理分布、影響其品質和生長發育的主要因素,因此,探究植物對低溫和高溫的適應性及生理機制具有重要意義和作用[2]。
低溫脅迫會引起植物新陳代謝紊亂,通過探析植物對低溫的生理響應來研究和評價其耐寒性是較常見的方法。Plazek等[3]發現抗寒植株在低溫脅迫下的細胞膜變化比低溫敏感植株小,電導率增幅小。低溫脅迫下的細胞外結冰現象可使胞內水向胞外轉移,從而導致細胞受到程度更為嚴重的滲透脅迫,電導率升高[4]。Airaki等[5]將辣椒葉片在8℃低溫下脅迫1~3d發現低溫脅迫使細胞膜脂質過氧化加重,誘導了氧化脅迫。為此,植物通過啟動一系列抗氧化酶系統來清除細胞內的活性氧(ROS),從而減低氧化脅迫對其造成的傷害[6]。另外,低溫脅迫下,植物體內也能積累更多滲透調節物質如脯氨酸、可溶性糖、可溶性蛋白,從而增強其滲透調節能力,提高對逆境脅迫的耐受性[7,8]。
鳳仙花屬植物種類豐富,在我國分布范圍廣,因花型奇麗、顏色和形態多樣性高,極具觀賞價值,但目前在花卉園藝中開發利用的品種還較少。綠萼鳳仙花(ImpatienschlorosepalaHand.Mazz.)為一年生或多年生草本花卉[9],主要分布于貴州、廣東、廣西等地,常生于疏林溪旁或山谷水旁陰處[10],具有低矮、分枝能力強、花大色艷、花期長、一年多次開花等特點,是可開發為園林地被植物的優良植物材料。因此,研究綠萼鳳仙花在低溫脅迫下的生理適應性,對其引種馴化和抗寒品種選育具有重要指導意義。
目前尚未見有關綠萼鳳仙花對低溫脅迫的生理響應的研究報道。因此,本研究在前期研究基礎上,設置2℃和0℃兩種低溫處理,以20℃為對照,通過分析不同處理時間葉片中相關抗氧化生理指標的變化動態,探析綠萼鳳仙花對低溫脅迫的生理響應機制,以期為其開發利用提供理論依據。
1材料與方法
1.1試驗材料
試驗用綠萼鳳仙花于2020年7月分株于西南林業大學樹木園。2021年1月選取長勢一致且健康的成年植株進行試驗。
1.2試驗方法
將所選綠萼鳳仙花植株分別在20、2℃和0℃的光照培養箱內處理0、12、24、48、72h,以20℃為對照(CK),共計15個處理,每個處理3盆植株。選取當年生莖上的成熟葉洗凈并擦干,用于測定相關指標,每次試驗重復3次。
1.3生理指標及測定方法
1.3.1細胞傷害率(CIR) 參考Chen等[11]的方法進行測定,計算公式:
CIR(%)=[(R-CK)/(R0-CK)]×100。式中,R為組織殺死前外滲液的電導率值;R0為組織殺死后外滲液的電導率值;CK為處理0h的外滲液電導率值。
1.3.2葉綠素含量 采用直接浸提法進行測定[12]。
1.3.3抗氧化物酶活性和丙二醛(MDA)含量 MDA含量采用硫代巴比妥酸(TBA)法測定,超氧化物歧化酶(SOD)活性采用氮藍四唑(NBT)光還原法測定,過氧化物酶(POD)活性用愈創木酚法測定,過氧化氫酶(CAT)活性用過氧化氫還原法測定[13]。
1.3.4滲透調節物質含量 脯氨酸(PRO)含量采用酸性茚三酮法測定,可溶性糖(SS)含量用蒽酮比色法測定,可溶性蛋白(SP)含量用考馬斯亮藍G-250比色法測定[14]
1.4數據處理
用MicrosoftExcel2010進行數據處理和作圖,用SPSS26.0軟件進行多重比較(Duncan’s法)和Spearman相關分析。
2結果與分析
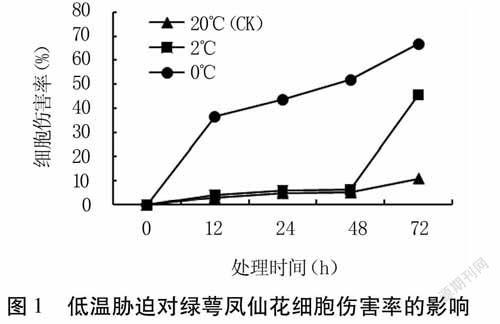
2.1低溫脅迫對綠萼鳳仙花細胞傷害率的影響
由圖1可知,20℃條件下,CIR隨處理時間延長緩慢上升,但上升幅度很小,處理72h的CIR僅11%。2℃低溫下,48h內的CIR與對照相當,但48h后急劇上升,72h達到46%,顯著高于對照。0℃低溫下,CIR在處理后即急劇上升,至12h已上升至37%,之后增幅略緩,至72h達到67%,與其他兩處理間差異顯著(P<0.05)。可見,綠萼鳳仙花的細胞傷害率受脅迫溫度和時間的共同影響。
2.2低溫脅迫對綠萼鳳仙花葉綠素含量的影響
由圖2可知,2℃和0℃低溫處理顯著降低綠萼鳳仙花的葉綠素含量(P<0.05),但兩種低溫處理間差異不顯著;低溫脅迫12~48h內各溫度處理的葉綠素含量無顯著差異,至72h降至最低,分別低于CK(20℃)53%和60%。CK處理下,綠萼鳳仙花葉片中的葉綠素含量波動較小,均在12~15μmol·g-1FW。
2.3低溫脅迫對綠萼鳳仙花葉片保護酶活性的影響
由圖3A可知,除2℃脅迫12h的SOD活性與CK差異不顯著外,兩種低溫脅迫均能使綠萼鳳仙花SOD活性顯著升高(P<0.05),且2℃處理的SOD活性顯著高于0℃處理;隨脅迫時間延長,2℃和0℃處理的SOD活性先升高,并分別于24、12h達到最高值,之后逐漸下降,72h時仍顯著高于CK57%和31%。CK處理的綠萼鳳仙花SOD活性在72h內呈下降趨勢,但降幅很小,為0.89%~17.86%。
與CK相比,0℃低溫脅迫顯著降低綠萼鳳仙花的POD活性,且隨脅迫時間的延長呈逐漸降低趨勢,至72h時低于CK71%;2℃低溫脅迫下48h內的POD活性與CK差異不顯著,呈先升后降變化趨勢,72h時大幅降低,顯著低于CK43%(P<0.05)。CK的POD活性在72h內呈緩慢下降后又緩慢上升狀態,降幅為0.43%~16.52%,處理48h的值最低(圖3B)。
處理0~12h,2℃和0℃低溫處理對綠萼鳳仙花CAT活性的影響與CK無顯著差異,但均在24h顯著提高CAT活性,達到峰值;處理48h時,兩低溫處理的CAT活性仍高于CK,其中2℃處理仍顯著高于CK;至72h,兩低溫處理的CAT活性分別低于CK10%和53%,其中,0℃處理與CK間差異顯著(P<0.05)。兩低溫處理間比較,0~12h兩者間CAT活性無顯著差異,24~72h0℃處理的CAT活性顯著低于2℃處理(P<0.05)。20℃條件下,處理72h內綠萼鳳仙花的CAT活性為1.09~1.23U·g-1FW,呈小幅上升趨勢(圖3C)。
2.4低溫脅迫對綠萼鳳仙花葉片MDA含量的影響
由圖4可知,2℃和0℃低溫處理能夠促進綠萼鳳仙花的MDA積累,且積累量隨處理時間的延長而增加。2℃條件下處理72h的積累量顯著高于CK80.85%;而0℃條件下處理12h的積累量即顯著高于CK(P<0.05),72h時則高于CK2.31倍。CK處理的MDA含量變化很小,在1.28~1.41μmol·g-1FW范圍內波動。
2.5低溫脅迫對綠萼鳳仙花葉片細胞滲透調節物質含量的影響
由圖5A可知,2℃脅迫12~72h的PRO含量均高于CK,并在脅迫24h后達到差異顯著水平;0℃脅迫12~24h的PRO含量顯著高于CK,之后逐漸下降,72h時略低于CK,差異不顯著。隨脅迫時間的延長,2℃和0℃處理的PRO含量均先升高后下降,分別于處理48h和12h達到峰值。CK的PRO含量在72內無明顯變化,穩定在13~14μg·g-1。
由圖5B可知,2℃脅迫0~72h的SS含量先下降后上升,24h時最低,顯著低于CK24.31%,48~72h則顯著高于CK,尤其72h時,比CK高1.02倍;0℃處理下,隨脅迫時間的延長,SS含量呈先上升后下降再上升的變化趨勢,12~24h顯著高于CK,48h時則顯著低于CK,之后快速上升,72h時顯著高于對照1.24倍。CK的SS含量在0~72h內先降低后上升,在218~255μmol·g-1FW范圍內小幅變化。
由圖5C可知,低溫脅迫能夠增加綠萼鳳仙花葉片中的SP含量,2℃處理48h后差異達顯著水平(P<0.05),尤其處理72h時達到CK的3.39倍;0℃條件使SP含量在處理12h時即顯著高于CK,之后隨處理時間延長逐漸升高,24h后增幅加大,72h時達到CK的5.93倍。CK的SP含量在處理的72h內變化很小,保持在5.4~6.9μmol·g-1FW。
2.6綠萼鳳仙花葉片各項生理指標的相關性分析
由表1可知,CIR與脅迫時間始終呈極顯著正相關,表現為隨著脅迫時間的延長顯著上升,且溫度越低,兩者的相關性越高。2℃和0℃低溫脅迫下,CIR與SP和MDA含量一直呈極顯著正相關,與葉綠素含量呈極顯著負相關;POD在2℃低溫下與CIR顯著相關,在0℃下則與CIR極顯著相關;PRO在2℃低溫下與CIR呈現極顯著正相關;SS和CAT在2℃和0℃低溫脅迫下與CIR相關不顯著。表明MDA、SP和葉綠素含量及POD活性在綠萼鳳仙花受低溫脅迫的不同階段對細胞損傷進行有規律的響應,而SOD和CAT活性及PRO和SS含量對綠萼鳳仙花響應低溫脅迫無明顯規律。
3討論
膜系統在植物抵御低溫冷害中起重要作用,冷害會使植物體內產生大量的活性氧,導致細胞膜脂過氧化,增加細胞膜透性,使電解質滲透率提高[15,16]。有研究表明,隨低溫持續時間的延長,電解質滲透率呈“S”形變化[17]。SOD、POD和CAT是植物抗氧化系統中清除活性氧的主要酶,在遭受逆境脅迫后,植物體首先啟動SOD,將O·-2轉化為H2O2和O2,POD和CAT則進一步將H2O2分解為H2O和O2,從而減輕活性氧對膜系統的傷害[18-21]。丙二醛(MDA)是細胞膜脂過氧化的產物,隨著脅迫溫度的降低其含量會不同程度地上升,是評價植物抗寒性強弱的參考指標之一[22-24]。本研究結果表明,與20℃對照相比,2℃和0℃低溫處理能提高綠萼鳳仙花葉片中的SOD和CAT活性,溫度越低提高越快,說明受到低溫脅迫后,綠萼鳳仙花快速啟動活性氧清除系統,以保護膜系統免受活性氧的傷害,此時MDA積累量較低,細胞傷害率(CIR)上升緩慢;但隨著脅迫時間的延長,SOD、POD和CAT活性均降低,且溫度越低其活性越低,MDA含量顯著提高,CIR急劇上升,說明長時間的低溫脅迫打破了植物體內的活性氧“產生—清除”平衡,造成大量的活性氧積累,對膜系統造成嚴重傷害。這與在金花茶[25]、南天竹[26]、冰草屬植物[27]、大葉鐵線蓮[28]等中的研究結果基本一致。
葉綠素含量的高低可反映植物光合能力的強弱[29]。低溫脅迫會抑制葉綠素合成相關酶的活性,從而使植物體內葉綠素含量降低,影響其光合作用[30]。本試驗結果也表明,2℃和0℃低溫脅迫顯著降低綠萼鳳仙花葉片中的葉綠素含量,且隨脅迫時間的延長呈下降趨勢,可能是因為綠萼鳳仙花對低溫的應激反應使體內葉綠素合成相關酶活性快速降低,導致葉綠素含量急劇下降,而長時間低溫脅迫使傷害進一步加劇,導致葉綠素含量繼續降低。
脯氨酸(PRO)、可溶性糖(SS)和可溶性蛋白(SP)是細胞內重要的滲透調節物質。SP在植物體內起著維持滲透調節及防止原生質體脫水的作用[31,32],低溫冷害會引起細胞蛋白質的一系列變化,主要表現在SP含量增加,是植物產生抗寒性的重要過程[33];而冷馴化下,植物的抗寒性可因SS和PRO的累積而提高[34-36]。本研究結果表明,2℃和0℃低溫脅迫能提高綠萼鳳仙花葉片中的PRO、SP和SS含量。其中,2℃脅迫期間的PRO含量均高于對照,24h后差異顯著,而0℃脅迫24h內的PRO含量顯著高于對照,之后與對照差異不顯著;兩低溫處理的PRO含量分別于脅迫12、48h達到峰值,溫度越低,達到峰值的時間越短,PRO含量下降越快。這與李慶華等[34]對不同基因型日本柳杉的抗寒性研究結果相似。SS含量在兩種低溫脅迫期間波動變化,但均在脅迫72h時顯著高于對照,總體呈現溫度越低,SS積累越快,含量越高,這可能是因為低溫脅迫會影響淀粉及蛋白質分解酶的活性,進而影響細胞內的SS含量[37]。SP含量隨低溫脅迫時間的延長而增加,均在脅迫72h時最高,且溫度越低,SP積累越快,含量越高,可能是因為低溫脅迫使部分抗寒蛋白質合成增加,從而提高植株內的滲透調節作用以減輕低溫對其的傷害。
對不同溫度下綠萼鳳仙花各生理指標間的相關性進行分析,發現CIR與脅迫時間始終呈極顯著正相關,且相關性隨溫度的降低而升高。進一步分析CIR與其他指標的相關性,發現MDA、SP和葉綠素含量及POD活性在綠萼鳳仙花受低溫脅迫的不同階段對細胞損傷響應明顯。
4結論
2℃和0℃低溫處理72h能對綠萼鳳仙花造成一定傷害,且溫度越低、脅迫時間越長,傷害越重,0℃脅迫下生長受抑并表現出一定的寒害特征,因此,綠萼鳳仙花引種時應做好防寒保護措施,預防低于0℃的極端溫度傷害。CIR、葉綠素含量、MDA含量、SP含量和POD活性可作為綠萼鳳仙花耐寒性鑒定的主要指標,SOD和CAT活性、PRO和SS含量可作為次要指標。本研究結果可為綠萼鳳仙花的引種馴化、花期調控、抗寒育種等提供參考。
