骨碎補總黃酮對缺氧環境中犬骨髓間充質干細胞成骨分化潛能的影響
2022-04-24 02:52:02龍亞麗田啟會
畜牧獸醫學報 2022年4期
龍亞麗,田啟會
(1.甘肅農業大學動物醫學院,蘭州 730070; 2.甘肅畜牧工程職業技術學院,武威 733006)
骨髓間充質干細胞(bone marrow mesenchymal stem cells, BMSCs)屬于骨髓基質干細胞,同時具有橫向向骨分化的能力,因此被認為是骨組織損傷修復的種子細胞之一[1-2]。但骨組織損傷通常伴隨著嚴重的組織腫脹,形成局部缺氧環境[3]。文獻報道,缺氧一方面可以導致BMSCs生物學特性改變,同時還會造成其分化能力的異常,這為其在應用中的有效性提出挑戰[4-5]。BMSCs骨分化能力是受損骨組織再生和修復的基礎,因此,如何避免缺氧環境中BMSCs的成骨分化能力異常改變,是BMSCs應用前必需解決的關鍵問題。
BMSCs大量存在于脊髓中,骨碎補作為補腎中藥,根據“腎主骨生髓”理論,其極可能是能夠促進BMSCs增殖和成骨分化的重要藥物[6-7]。因此,在中醫理論的支持下,骨碎補總黃酮(total flavonoids ofDrynariaRhizoma, TFDR)可能對BMSCs成骨分化能力具有影響作用。本研究擬通過建立缺氧微環境,并探究骨碎補總黃酮對缺氧環境中BMSCs成骨分化的影響作用。
1 材料與方法
1.1 材料
1.1.1 細胞來源 犬骨髓間充質干細胞(BMSCs)細胞株購自美國 ScienCell 公司(編號:7500)。
1.1.2 主要試劑及儀器 骨碎補總黃酮(北京岐黃制藥有限公司,批號:國藥準字Z20133051);間充質干細胞基礎培養基、胎牛血清(ScienCell);RT-PCR轉錄試劑盒(TaKaRa公司,批號:AKG1212A),RT-PCR熒光定量試劑盒(TaKaRa公司,批號:AL12412A),兔抗人Runx2、Osterix多克隆抗體(GeneTex),山羊多克隆抗體二抗抗兔(Alexa Fluor?488)(Abcam公司),OriCell TM 狗骨髓間充質干細胞成骨誘導分化培養基試劑盒(Cyagen 批號:CAXMX-90021),JC-10細胞線粒體膜電位(MMP)活細胞熒光染料(美國AAT公司,批號:2191362),堿性磷酸酶(AKP)檢測試劑盒(南京建成生物工程研究所,批號: 20170719);PCR引物序列見表1。

表1 PCR引物序列
CO2細胞低氧培養箱(日本SANYO公司),酶標儀(美國BIO-RAD公司),光學顯微鏡、激光共聚焦顯微鏡FV10-ASW 2.1 Viewer(日本Olympus公司),FACS CelestaTM型流式細胞儀(美國BD公司),RT-PCR儀(美國BIO-RAD公司)。
1.2 方 法
1.2.1 試驗分組 試驗分為空白組(Control)、低氧組(Hyp)、TFDR干預組(Hyp+TFDR),低氧組和TFDR組細胞培養氧濃度為10%[8],TFDR的濃度為50 μg·mL-1(前期預試驗篩選出能夠促進BMSCs最大增殖的濃度)。
1.2.2 細胞培養 取第3代對數期生長的BMSCs細胞,并調整成細胞濃度為1×104個·mL-1的單細胞懸液,接種于包被明膠的六孔板內,每組定容為2 mL,并設3個平行重復。將細胞置于常氧環境培養箱中培養,至細胞匯合達到60%~70%時,低氧組和TFDR干預組置于10%的氧濃度環境中培養4周,TFDR干預組加入含有50 μg·mL-1TFDR的培養基。在第2周結束后,向六孔板中加入骨髓間充質干細胞成骨誘導分化誘導劑,繼續在10%的氧濃度環境中成骨誘導分化14 d,TFDR干預組誘導培養基中含有50 μg·mL-1TFDR。
1.2.3 細胞形態觀察 細胞成骨誘導過程中,每天將各組細胞置于顯微鏡下進行形態觀察,并采集圖像,重點觀察鈣結節形成情況。
1.2.4 茜素紅染色 成骨誘導結束后,棄掉六孔板中剩余誘導液,PBS清洗;每孔加入 2 mL細胞固定液,固定 30 min后,棄掉細胞固定液,PBS清洗兩次。每孔加入1 mL茜素紅染色液,染色3~5 min。PBS清洗兩次,于顯微鏡下觀察。
1.2.5 堿性磷酸酶(ALP)活性測定 誘導完成后,棄培養瓶中誘導液,PBS反復搖晃清洗,用細胞裂解液裂解細胞,反復吹打后,12 000 r·min-1離心6 min,取上清,檢測ALP活性。在96孔板中分別加入標準品和各組樣品與底物的混合液,37 ℃條件下孵育30 min。每孔加入終止液終止反應,于酶標儀檢測每孔OD405 nm值。ALP活性單位為“U·g-1”。
1.2.6 流式細胞術檢測線粒體膜電位 誘導完成后,用胰酶消化細胞,用PBS清洗1遍后,用PBS將細胞重懸,并移入流式上樣管,將JC-10熒光探針工作液加入細胞懸液,37 ℃避光孵育20 min后,通過流式細胞儀進行檢測。當膜電位水平低時,激發波長527 nm,通過FITC熒光通道收集;當膜電位水平高時,激發波長590 nm,通過PE熒光通道收集,每組細胞進行3次重復。
1.2.7 激光共聚焦顯微鏡檢測成骨分化關鍵基因熒光表達 將蓋玻片平鋪入24孔細胞培養板中,將干預后的細胞消化、吹散并接種于6孔板中,匯合度控制在20%左右,進行培養。待細胞匯合度達到60%~70%時,每個孔中加1 mL 4%多聚甲醛,室溫固定20 min,棄多聚甲醛,再加入1 mL甲醇繼續固定5 min。棄甲醇,加入1 mL免疫熒光用Blocking buffer,室溫封閉10 min。棄封閉液,加入含兔抗Runx2抗體(1∶500)和Osterix抗體(1∶500)的Blocking buffer,慢搖過夜。PBS液,洗3遍后,加入1 mL含Alexa Fluor? 488 F(ab)羊抗兔熒光二抗(500∶1)的Blocking buffer,避光慢搖1 h。PBS液潤洗細胞5遍,每遍5 min。載玻片上滴加含DAPI染料的封片液,將蓋玻片取出并倒扣在封片液上,于激光共聚焦顯微鏡下觀察,每組細胞隨機挑選3個視野,每個視野隨機選擇10個細胞進行熒光定量,熒光表達水平通過FV10-ASW 2.1 Viewer軟件進行分析。
1.2.8 RT-PCR檢測成骨分化關鍵基因表達水平 干預完成后,提取各組細胞中RNA并進行濃度測定。以20 μL的體系反轉錄合成cDNA,并進行實時熒光定量檢測,檢測成骨分化關鍵基因Runx2、Osterix的轉錄水平,反應條件:95 ℃變性1 min;[95 ℃ 10 s,57 ℃ 20 s,72 ℃ 20 s收集熒光]40個循環;熔解曲線制備65~95 ℃,0.5 ℃·s-1。

2 結 果
2.1 TFDR對低氧環境中BMSCs形態學的影響
分別于誘導成骨分化第4、7、14天在倒置相差顯微鏡下觀察細胞變化。空白對照組細胞生成較多高密度鈣結節,成片分布;與空白對照組細胞相比,在分化前期(第0~7天),低氧條件下BMSCs高密度鈣結節較少,后期(第8~14天)更明顯;與低氧組相比,加TFDR組細胞誘導分化鈣結節均明顯增多(圖1)。

圖1 TFDR對低氧環境中 BMSCs形態學的影響
2.2 TFDR對低氧環境中BMSCs鈣鹽茜素紅染色的影響
誘導各組細胞向成骨方向分化14 d,茜素紅染色后倒置顯微鏡下觀察,空白對照組BMSCs呈鮮紅色,著色均勻,緊密連接成片;而低氧組細胞紅色鈣結節分布較為分散,結節明顯較小;相較于低氧組,加TFDR組細胞生成鈣結節沉積數量增多(圖2)。

圖2 TFDR對低氧環境中BMSCs 茜素紅染色的影響(10×4)
2.3 TFDR對低氧環境中BMSCs ALP活性的影響
在405 nm波長條件下,檢測各組細胞ALP活性值,與空白對照組細胞相比,低氧組細胞ALP活性明顯降低,差異極顯著(P<0.01);與低氧組相比,TFDR組細胞ALP活性升高,差異顯著(P<0.05)(圖3)。

與Control組相比,*.P<0.05,**.P<0.01;與Hyp組相比,#.P<0.05
2.4 TFDR對低氧環境中BMSCs線粒體膜電位的影響
通過流式細胞術檢測各組細胞膜電位變化情況(圖4A~C),結果顯示,與空白對照組BMSCs相比,低氧組細胞線粒體膜電位降低,差異顯著(P<0.05);與低氧組比較,TFDR干預組細胞線粒體膜電位增強,差異顯著(P<0.05)(圖4D)。

A.Control組;B.Hyp組;C.Hyp+TFDR組;D.各組線粒體膜電位;與Control組相比,*.P<0.05;與Hyp組相比,#.P<0.05
2.5 TFDR對低氧環境中BMSCs成骨關鍵基因Runx2和Osterix熒光表達的影響
激光共聚焦顯微鏡觀察各組細胞Runx2和Osterix的表達,結果顯示,與空白對照組BMSCs相比,低氧組細胞Runx2和Osterix熒光表達強度明顯減弱,差異顯著或極顯著(P<0.05或0.01);與低氧組比較,TFDR干預組細胞Runx2和Osterix熒光表達強度增強,差異顯著(P<0.05)(圖5~7)。

圖5 激光共聚焦顯微鏡觀察各組BMSCs細胞Runx2的表達(10×40)
2.6 TFDR對低氧環境中BMSCs Runx2和Osterix基因水平表達的影響
通過RT-PCR檢測各組細胞Runx2和Osterix的轉錄水平,結果顯示,與空白對照組相比,低氧組細胞Runx2和Osterix基因轉錄水平降低;與低氧組比較,TFDR干預組細胞Runx2和Osterix基因轉錄水平增強(表2)。

表2 TFDR對低氧環境中BMSCs成骨相關轉錄因子的影響
3 討 論
多向分化潛能是BMSCs的重要特點,在經過誘導后,其可以向多個胚層的組織細胞進行分化,其中包括骨細胞[1-2]。因此,BMSCs在組織再生科學方面的應用是現在的研究熱點之一。目前,大量研究證實,BMSCs能夠修復骨組織損傷,因而,BMSCs在骨損傷的修復方面具有開發潛力[1-2, 9]。但是,骨損傷通常也會引起肌肉組織的水腫或者是血腫的形成,腫脹將造成局部壓迫,最終導致損傷局部形成缺氧環境[3]。盡管BMSCs的再生修復能力已經得到認可,但通過文獻可發現,缺氧環境十分容易導致其生物學特性的改變,在不同的低氧濃度下均能造成大鼠間充質干細胞成骨、成脂分化能力的降低[10],這也可能成為制約BMSCs進一步應用于臨床的關鍵。因此,有效地維持低氧環境下BMSCs成骨分化潛能,是其運用于臨床損傷修復的前提之一。
ALP廣泛分布于人體骨骼中,對骨形成具有重要意義,是BMSCs早期成骨分化能力評估的關鍵指標之一[11];Runx2是骨細胞的特異性轉錄因子,能夠調節骨組織的形成和重建,并與破骨細胞分化及細胞外基質的形成密切相關[12-13]。Osterix(Osx)處于Runx2 的下游位置,受到Runx2的表達影響,Osterix是干細胞向成骨細胞分化過程中的重要標志基因。Runx2主要是在成骨細胞早期未成熟分化時期進行調節,而成骨細胞在成熟分化時期主要由 Osterix 進行調控[14-15]。而在成骨分化終末期,鈣結節形成的多少是評估骨分化程度最關鍵的效應性指標。本研究以10%的氧濃度為缺氧標準進行低氧環境的模擬,并發現缺氧導致BMSCs成骨分化的早期調控酶ALP和早期調節基因Runx2的降低,成熟分化時期的調節基因Osterix表達降低,同時也引起關鍵的終末效應性標志物鈣結節形成的明顯減少。通過對成骨分化早期、成熟期及終末期相關標志物和基因的檢測結果均提示,在模擬的低氧環境中BMSCs成骨分化的能力降低明顯。細胞的分化需要能量供給,線粒體是細胞通過呼吸鏈進行能量代謝的關鍵場所,而在缺氧條件下,線粒體會因為產生的能量不足,無法持續支持BMSCs成骨分化,導致了成骨分化能力減弱[16-17]。通過檢測細胞線粒體膜電位來評估線粒體的活性,發現低氧條件下,BMSCs線粒體膜電位明顯下降,證實缺氧可以導致線粒體活性降低。
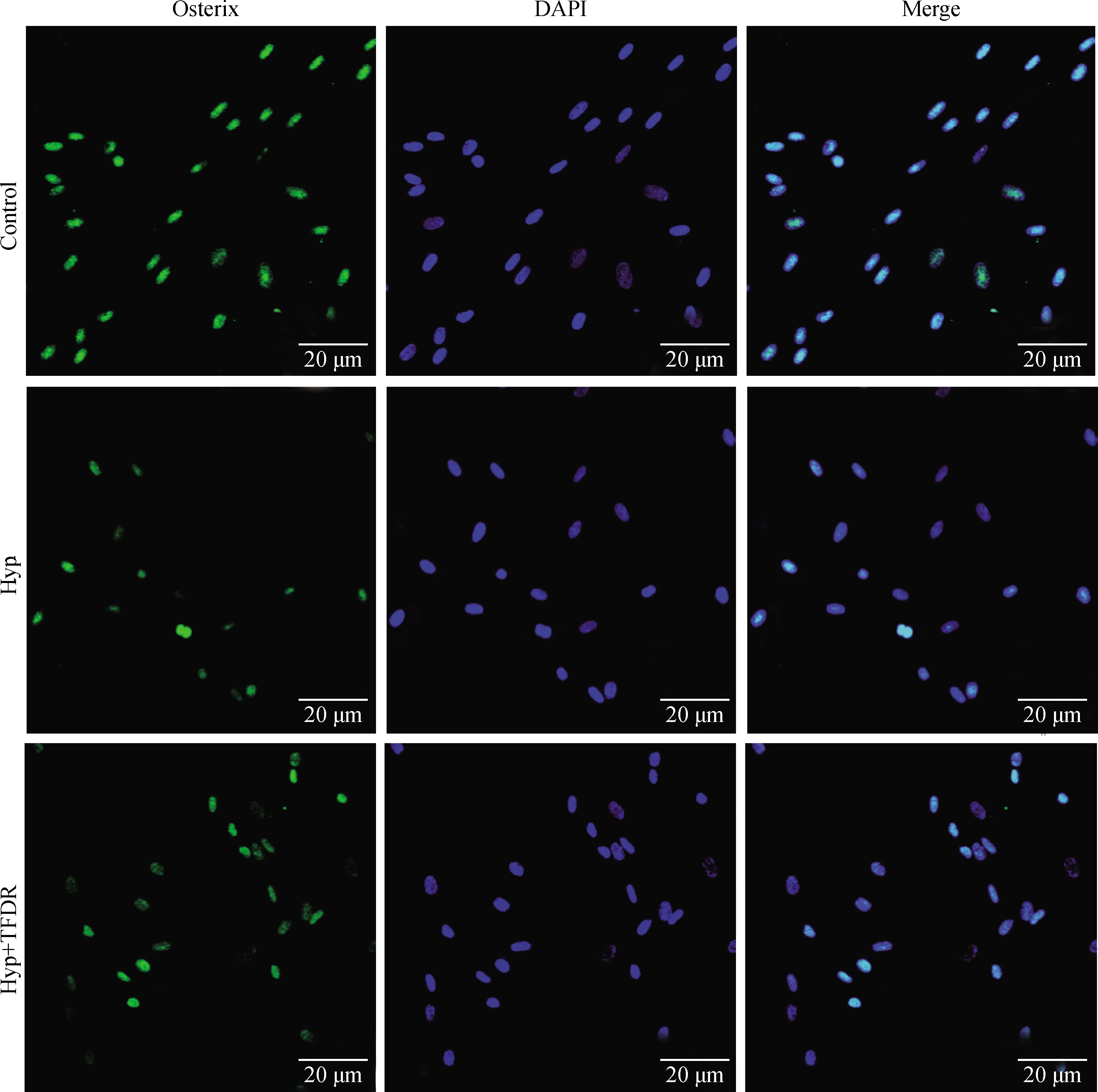
圖6 激光共聚焦顯微鏡觀察各組BMSCs細胞Osterix的表達(10×40)

與Control組相比,*.P<0.05,**.P<0.01;與Hyp組相比,#.P<0.05
傳統中藥骨碎補具有補腎強骨、活血止痛的功效,根據“腎主骨生髓”理論,骨碎補對骨相關疾病具有較好的治療作用[18]。而骨碎補的主要成分骨碎補總黃酮(TFDR)在免疫調節、抗炎、骨修復等方面具有明顯功效[5-6,19]。本研究用濃度為50 μg·mL-1的TFDR對低氧環境中BMSCs進行干預后,通過檢測上述指標,發現TFDR能夠有效地提高BMSCs成骨分化關鍵轉錄因子Runx2和Osterix表達,并促進其對成骨分化相關的酶ALP的轉錄、表達,增加末期的鈣鹽沉積量,線粒體活性升高,有效維持BMSCs在低氧環境中成骨分化能力的相對穩定。
綜上,缺氧環境會導致BMSCs成骨分化能力降低,而TFDR能夠維持缺氧環境中BMSCs成骨分化能力穩定。但是,體外模擬的低氧環境無法完全構建出體內的狀況,研究還應進一步通過體內成骨試驗證實TFDR對低氧環境中BMSCs成骨分化潛能的保護作用,但TFDR仍然值得被作為維護缺氧條件下干細胞生物穩定性的天然化合物進行進一步的研究。
4 結 論
骨碎補總黃酮能夠維持低氧濃度下犬BMSCs線粒體活性、促進成骨分化關鍵基因Runx和Osterix基因表達,增加鈣結節生成,進而提高低氧環境中犬BMSCs成骨分化能力,為骨碎補總黃酮防護缺氧環境中犬BMSCs的成骨分化能力降低以及BMSCs在骨折疾病治療中的應用提供試驗依據。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
