我國H9N2亞型禽流感病毒血凝素蛋白145和153位抗原位點變異分析
2022-04-24 02:51:56陳子軒全柯吉陳素娟彭大新劉秀梵
畜牧獸醫學報 2022年4期
陳子軒,張 楠,胡 群,全柯吉,秦 濤,陳素娟*,彭大新*,劉秀梵
(1.揚州大學獸醫學院,揚州 225009;2.江蘇省動物重要疫病與人獸共患病防控協同創新中心,揚州 225009)
H9N2亞型禽流感病毒(avian influenza virus, AIV)于1966年首次在美國威斯康星州被發現。我國于1992年在廣東首次報道發現H9N2亞型AIV,并隨之在南方各省迅速傳播。目前是中國流行最廣泛的低致病性AIV,可和其他病原發生混合感染造成死亡率上升,引起嚴重的經濟損失。此外,它可為新出現的重配病毒如H7N9亞型AIV等提供供體基因[1],同時報道顯示有部分家禽養殖從業者血清呈H9亞型AIV抗體陽性,因此病毒的傳播也存在很大的公共衛生隱患[2]。
為防控我國H9N2亞型禽流感,以A/Chicken/Shanghai/F/1998(H9N2)為代表的全病毒滅活疫苗在家禽養殖中廣泛使用[3]。目前,主要流行的毒株屬于h9.4.2.5譜系,HA基因的遺傳進化分析表明,當前流行株與疫苗株的核苷酸遺傳距離較遠[4-5]。基于交叉血凝抑制和交叉微量中和試驗的抗原圖譜分析研究中,流行株和疫苗株可形成不同的抗原分群,疫苗株的免疫血清難以有效中和抗原差異大的流行株,疫苗株與流行株不匹配,從而導致了免疫失敗[6]。在疫苗的免疫壓力下,AIV更易發生抗原漂移[7]。因此,探究H9N2亞型 AIV抗原變異的分子基礎是解析當前流行株抗原性改變的關鍵。
目前,雖已報道了H9N2 AIV抗原位點鑒定的相關研究,并將HA蛋白頭部的抗原區域分為兩個獨立表位[8]。但前人鑒定的與抗原性相關的位點在自然流行株中很多都保守,不足以解釋我國當前H9N2亞型AIV流行株抗原性改變的原因[9]。因此,本研究利用研制的單抗對抗原位點進行了鑒定,并對本實驗室2017—2019年分離的H9N2亞型AIV進行了抗原譜分析,分析相關抗原位點的變異規律。
1 材料與方法
1.1 病毒和單抗
H9N2亞型AIV A/chicken/Shanghai/F/1998(F98)毒株為本實驗室保存。其相應單抗2E4[10]和2D6[11]為本實驗室制備。
1.2 血凝抑制試驗和中和試驗
血凝抑制試驗(hemagglutinin inhibition,HI)按照世界動物衛生組織(World Organization for Animal Health,OIE)的標準進行操作。微中和試驗按照文獻[12]進行操作,利用Reed-Muench法計算病毒在CEF細胞上的TCID50,再將2 000 TCID50·mL-1的病毒工作液與等體積的兩倍倍比稀釋的抗體進行孵育,其中每種稀釋度設置3個重復,37 ℃孵育1.5 h后,按照100 μL·孔-1接種于長有CEF細胞的96孔細胞板中,37 ℃吸附1 h后,再換液為含有2 μg·mL-1TPCK胰酶的M199培養基培養,72 h后通過測定上清HA效價判定結果。HA效價為陰性的抗體最高稀釋度為該抗體的中和效價。
1.3 單抗逃逸突變株的篩選和抗原位點的鑒定
根據文獻報道[13-14]并做適當改進開展逃逸突變株篩選,將0.1 mL 的106TCID50的F98病毒與0.5 mL單克隆抗體進行混合,室溫孵育1 h后,將抗原抗體混合物接種于9~11日齡的SPF雞胚(n=3),72 h后收取具有HA效價的尿囊液,所收獲病毒尿囊液通過有限稀釋法進行進一步篩選。將稀釋至10-3的陽性尿囊液0.1 mL與單抗0.5 mL混合,室溫孵育1 h后,將抗原抗體混合物再稀釋至10-1、10-2、10-3、10-4稀釋度,200 μL·胚-1接種雞胚(n=3),72 h后收取最高稀釋度的有HA效價的尿囊液。再按上述有限稀釋法重復篩選1次。
提取純化后病毒的RNA,利用RT-PCR擴增HA基因片段,電泳切膠測序。使用MegAlign軟件比對逃逸株和母本毒株F98的氨基酸序列差異。
1.4 抗原譜分析
使用MEGAX軟件中的MUSCLE將實驗室2017—2019年度分離的H9N2亞型AIVHA基因進行序列比對,繪制Neighbor-joining遺傳進化樹,Bootstrap設置為1 000進行檢驗。使用iTOL對遺傳進化樹進行美化[15]。根據遺傳進化樹選擇代表性H9N2亞型AIV毒株,通過HI效價確定單抗與病毒的反應性。
1.5 HA基因序列比對
在全球流感數據共享平臺(Global Initiative on Sharing Avian Influenza Data,GISAID,https://platform.gisaid.org/epi3/frontend#2a8325)中下載從1996—2021年度(截至6月5日)中國地區禽類中分離的H9N2 AIVHA基因序列,使用Phylosuite軟件中的MATFF進行序列比對,統計145和153位點的氨基酸變異情況。
1.6 位點模擬分析
以蛋白質三維結構數據庫(PDB, https:/www.wwpdb.org/)中的H9N2亞型病毒HA1分子晶體結構(編號:1 JSD.1.A)為模板,在網站SWISS-MODLE(https://www.swissmodel.expasy.org/interactive)將F98 HA1蛋白序列用Alignment Mode方法建模。生成的HA1分子用pymol軟件進一步分析和修飾。
2 結 果
2.1 單抗2E4和2D6的生物學特性
2E4和2D6針對母本毒株F98的HI效價均為213;微量中和試驗結果顯示,2E4和2D6針對F98的中和效價分別為1∶10 240和1∶2 560(表1)。

表1 單抗的生物學特性
2.2 單抗逃逸突變株的篩選和抗原位點的鑒定
在單抗2E4和2D6作用下收獲的陽性尿囊液,再在雞胚中經過兩輪有限稀釋法的純化后獲得了兩株突變株(m2E4-1和m2D6-1),對應篩選抗體的HI效價均≤22(表2)。進一步對突變株m2E4-1和m2D6-1的HA基因與母本毒株F98的HA基因進行比對,發現m2E4-1在434位核苷酸位點發生G→A的突變,導致145位氨基酸位點產生了S→N的替換,由此形成了145—147位NGT模式的潛在糖基化位點。m2D6-1在459位發生A→T的突變,相應的出現了D153E的氨基酸替換(表3)。
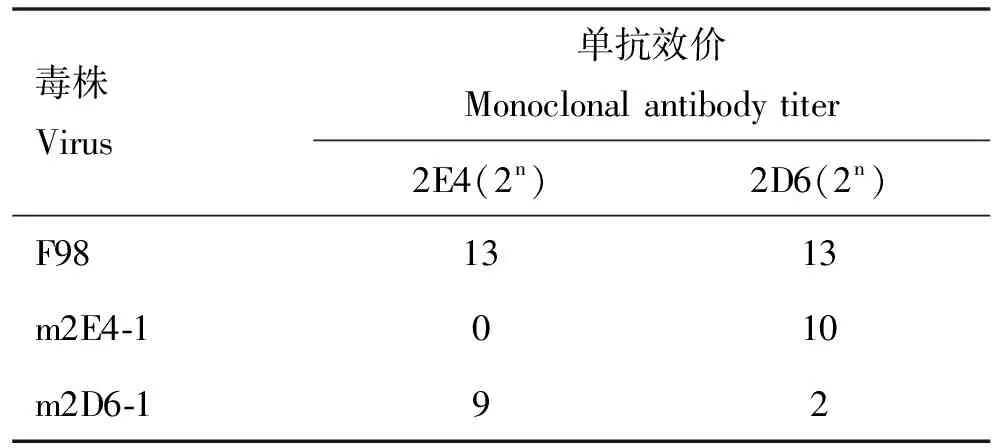
表2 逃逸株與單克隆抗體交叉血凝抑制結果
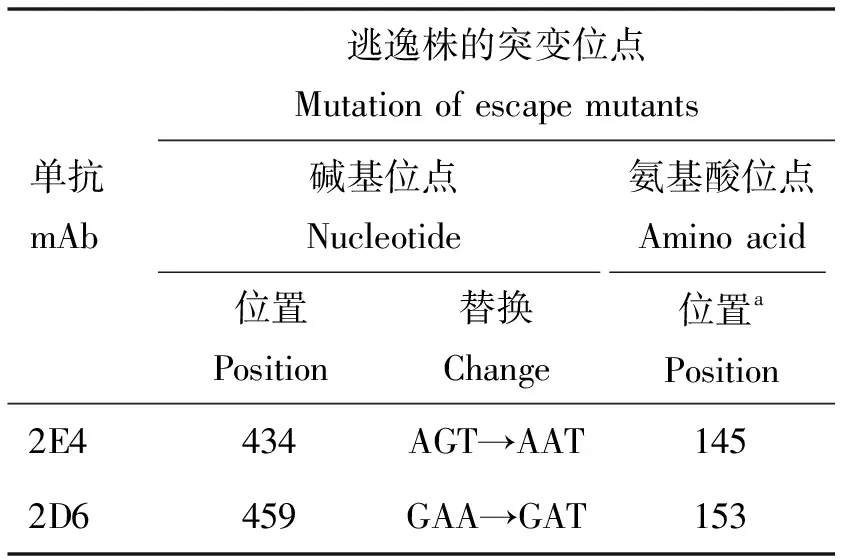
表3 抗原位點鑒定結果
2.3 抗原譜分析
2017—2019年度分離的H9N2亞型AIVHA基因遺傳進化樹顯示分離株均屬于h9.4.2.5譜系(圖1)。

●.參考株;△.抗原譜分析毒株
在遺傳進化樹上,將毒株劃分為兩個大分支Group 1(32/109)和Group 2(77/109),兩個Group 的組間平均遺傳距離為7.22%(圖1)。
從Group 1挑選的5株代表株和Group 2挑選的16株代表株測定與單抗的反應性。Group 1的毒株與單抗2E4的HI效價均≥27,與單抗2D6的HI效價均≥210;Group 2中所有毒株與單抗2E4的HI效價均在20~22,A/chicken/Jiangyin/JY110712/2018和A/chicken/Jiangyin/JY010220/2019毒株與單抗2D6的HI效價分別為25和24以外,其余14株毒株HI效價均為20(表4)。
比對用于抗原譜分析毒株的HA基因的145位和153位氨基酸發現,Group 1毒株的HA蛋白在145位和153位均分別為S(5/5)和D(5/5),Group 2毒株HA蛋白145位均突變為D(16/16),153位突變為G(15/16)。但A/chicken/Jiangyin/JY110712/2018與單抗2D6的HI效價為25,但是其153位卻為D(表4)。
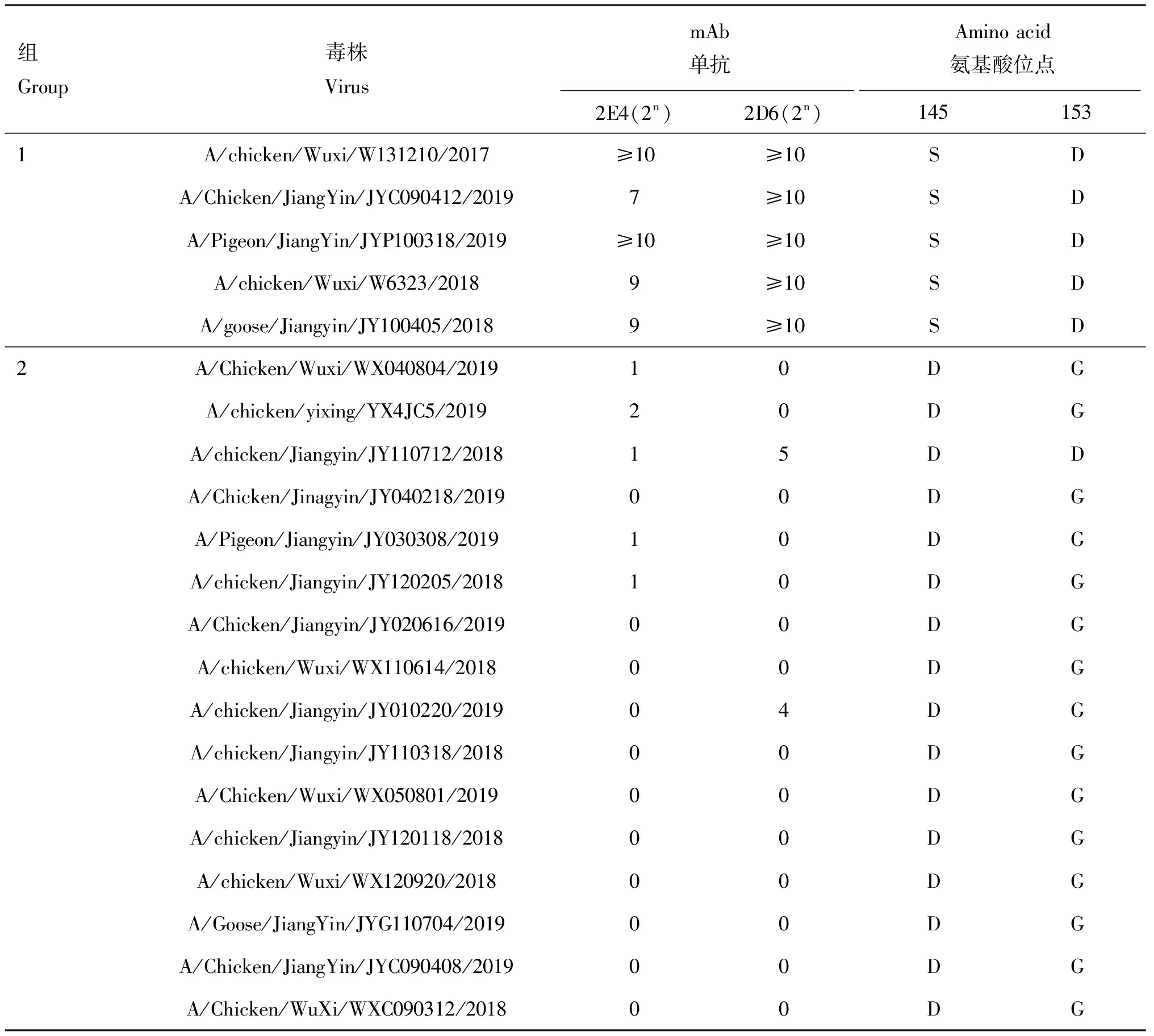
表4 單抗2E4、2D6抗原譜分析結果和145位、153位氨基酸位點的突變情況
2.4 禽源H9N2亞型AIV相應位點自然突變率分析
自GISAID中下載的禽源H9N2毒株AIV HA蛋白序列,對145位分析后發現,在1996—2009年和2010—2014年分離株中,含145S的分離株都占57%,在2015—2021年分離株中,含145D的分離株占68%。在1996—2009年和2010—2014年分離株中,含145N的分離株分別占6%和10%,2015—2021年無含145N的分離株。對153位分析后發現,含153D的分離株在1996—2009年和2010—2014年中分別占57%和53%,含153G的分離株在2015—2021年占66%。雞源和鴨源毒株變化趨勢一致(圖2)。
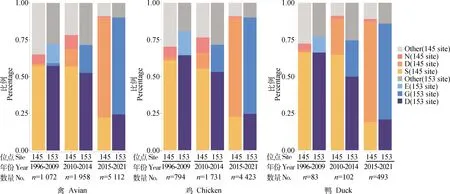
圖2 禽源H9N2 亞型AIV HA蛋白145和153位氨基酸位點的自然突變率
2.5 抗原位點結構模擬分析
從HA1分子的模擬構象可以看出,145和153位均位于分子的頭部的受體結合區域(receptor binding site,RBS)附近(圖3)。與145S相比,145 N和145D的殘基側鏈更長,突出HA1分子表面。與153D相比,153E和153G的殘基側鏈更長,突出HA1分子表面。
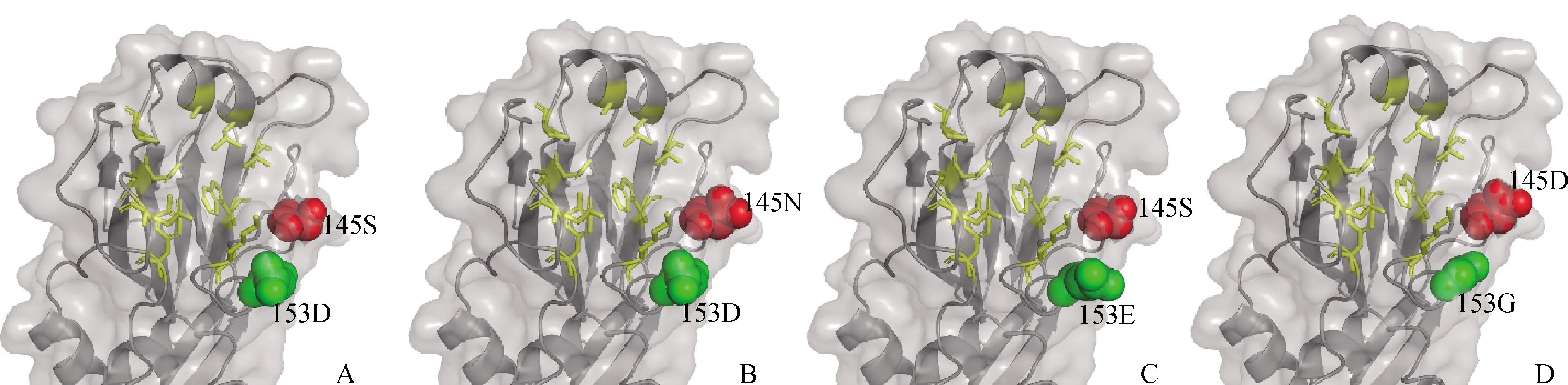
A.F98 HA1(145S 153D)分子模擬構象;B.F98 HA1(145N 153D)分子模擬構象;C.F98 HA1(145S 153E)分子模擬構象;D.F98 HA1(145D 153G)分子模擬構象。黃色:Y109、S148、W161、T163、N191、P194、A198、L202、Q234和G236為RBS區域
3 討 論
H9亞型AIV是我國感染最廣泛的低致病性AIV,目前的H9亞型毒株在各種選擇壓力下已經發生了變異,導致抗原性出現了改變。從本實驗室前期制備的H9N2亞型AIV HA蛋白單抗中選取了兩株HI效價高的單抗,經過鑒定也有很高的微中和效價。將F98毒株與單抗孵育后在SPF雞胚中傳代,篩選到逃逸突變株m2E4-1和m2D6-1,其HA蛋白分別擁有S145 N和D153E的突變。Kaverin等[16]將鑒定的抗原位點劃分成了Site Ⅰ、Site Ⅱ和Overlapping 3個區域,其中145位于Site Ⅰ和Site Ⅱ重疊的Overlapping區域,153位于Site Ⅱ區域。單抗2E4對m2D6-1的HI效價為29,單抗2D6對m2E4-1的HI效價為210,和母本毒株相比略有降低,證明這兩個抗原位點相互影響較小,與Kaverin等[16]的結果相一致。
S145N的突變使得HA蛋白在145—147位產生了1個NGT氨基酸模式的潛在糖基化位點。研究表明,在毒株的HA分子對唾液酸受體處于高親和力時,HA分子頭部RBS附近的糖基化位點會影響病毒對細胞的入侵,從而產生對疫苗免疫的逃逸[17]。朱寅彪等[13]也利用一株HI中和效價很高(分別為214和104.3)的HA單抗篩選到S145N的單抗逃逸突變體,另外他發現HA基因為145T的毒株也可以發生逃逸,而我們的抗原譜分析中發現擁有145D突變的毒株也會引起單抗逃逸,因此糖基化不是該位點逃逸單抗的唯一機制。并且145和153兩個位點均位于HA1分子的頭部RBS附近,推測單抗的中和效果來源于識別了RBS附近的表位,從而屏蔽RBS與唾液酸受體的結合[18]。并且相對于F98毒株145S和153D的結構,突變后的氨基酸因為相比于原模式氨基酸側鏈更長,更突出于HA1分子的表面,可能會直接影響到與抗體的結合能力[19]。
申松瑋等[20]將2017—2018年度分離的H9N2亞型AIVHA基因劃分為兩個組,其組間遺傳距離為6.71%。筆者劃分的Group 1和Group 2的組間遺傳距離為7.22%,與其基本一致。申松瑋等[20]對兩個組代表株的交叉HI結果進行R值評估后發現,兩個組內抗原性相近,但兩組間的抗原性不同,可分為兩個抗原型。本研究HI抗原譜分析顯示示,Group 1內毒株與單抗的反應性好,而Group 2內毒株與單抗的反應性差,單抗2E4反應譜與基因分型一致,表明單抗2E4可用于當前H9N2亞型AIV的抗原分型。在145位,Group 1代表株均為S,Group 2代表株均為D;在153位,Group 1代表株均為D,Group 2代表株除1株外其余均為G。可見S145D的替換在兩個Group 中的符合性更高,145位更適合作為鑒定病毒抗原變異的分子靶標。
朱寅彪等[13]發現含145N的病毒在雞源和鵪鶉源的自然分離率為10%左右(截至2015年),這與作者分析的2014年之前145N占比相一致。但1996—2021年間禽源H9N2亞型AIV分析顯示近年來含145D和153G的突變株占比分別為68%和66%,說明Group 2抗原型已成為流行的主要抗原型。因此利用單抗2E4對流行株進行抗原分型,加強對145位氨基酸位點的檢測尤為重要。
4 結 論
單抗2E4與2D6分別識別H9N2亞型禽流感病毒HA頭部的145位和153位抗原位點。我國流行株中存在兩種抗原群,單抗2E4可以對兩種抗原群進行區分,抗原位點145位是鑒別Group 1和Group 2抗原群差異的分子靶標之一,為H9N2亞型禽流感的防控提供了疫苗選擇依據。
