RFRP-3對豬卵母細胞體外成熟的影響
2022-04-24 02:51:54賈金美馮建豪陽美霞張羽芳張虹亮王水蓮
畜牧獸醫學報 2022年4期
關鍵詞:研究
賈金美,馮建豪,陽美霞,張羽芳,張虹亮,王水蓮
(湖南農業大學動物醫學院,長沙 410128)
卵母細胞的成熟是哺乳動物雌性生殖和胚胎發育的關鍵,需要經歷復雜的減數分裂停滯和恢復,該過程受多種激素和生長因子的調控[1]。緊密圍繞在卵母細胞周圍的卵丘顆粒細胞參與了卵母細胞減數分裂的阻滯和恢復,促進卵母細胞成熟.。卵丘顆粒細胞與卵母細胞相互作用,進行雙向的營養傳遞與信息交換,這種聯系在卵母細胞成熟過程中必不可少[2-3]。體外培養裸卵會導致細胞核和細胞質發育異常,但培養卵丘卵母細胞復合體(cumulus-oocyte complexes,COCs)時,細胞則正常成熟[4-5]。在卵母細胞成熟過程中,卵丘顆粒細胞逐漸擴展,卵丘擴展需要卵丘擴展因子透明質酸合酶2(hyaluronan synthase 2, HAS2)、穿透素3(pentraxin 3, PTX3)和前列腺素內過氧化物合酶2(prostaglandin-endoperoxide synthase 2, PTGS2)的參與,其中HAS2是生成透明質酸的關鍵酶,透明質酸是卵丘擴展基質的基本成分,PTGS2和PTX3具有維持擴展基質的作用[6-7]。卵母細胞自身分泌的生長分化因子9(growth differentiation factor 9, GDF9)和骨形態發生蛋白15(bone morphogenetic protein 15, BMP15)通過誘導卵丘擴展因子(PTGS2、HAS2和PTX3)表達,引起細胞外基質分泌,促進卵丘細胞擴展[8-9]。王兆琛等[10]體外添加GDF9和BMP15培養綿羊卵母細胞發現,GDF9和BMP15有利于卵丘擴展,促進卵母細胞成熟。卵丘顆粒細胞產生的環磷酸腺苷(cyclic adenosine monophosphate, cAMP)被運送到卵母細胞內[11],細胞內不斷累積的cAMP能夠啟動cAMP依賴性蛋白激酶(PKA)途徑,阻止成熟促進因子(maturation promoting factor,MPF)活化,使卵母細胞停滯在M期[12]。MPF是由細胞周期蛋白依賴性激酶1(cyclin-dependent kinase 1,CDK1)和細胞周期相關蛋白B(cyclin B)組成的復合物,主要調控卵母細胞減數分裂的恢復。MPF的活性主要由細胞質中的調節亞基cyclin B在細胞周期中的不斷活化和衰減來調節,隨著卵母細胞的成熟,cyclin B含量逐漸上升,誘導CDK1蛋白發生去磷酸化,形成活化的MPF進而調控卵母細胞的成熟[13]。
Tsutsui等[14]于2000年在日本鵪鶉中發現了一種C端具有RF酰胺結構的神經肽,它以劑量依賴的方式抑制了體外培養的鵪鶉垂體中促性腺激素釋放激素的釋放,因此被命名為促性腺激素抑制激素(GnIH)。隨后的研究表明,哺乳動物體內存在與鳥類GnIH具有相似生理作用的同源物RFRP-3[15]。在哺乳動物中,Li等[16]發現RFRP-3的同源物GnIH除定位在豬的下丘腦等中樞神經系統外,同樣在其它器官中也有表達,尤其在生殖系統細胞中,推測RFRP-3可能在豬的生殖系統中也發揮調控作用。研究表明,使用不同劑量的RFRP-3處理豬下丘腦細胞可以顯著抑制促性腺激素釋放激素分泌及相關基因水平的表達,處理豬顆粒細胞可以顯著降低雌二醇(estradiol, E2)的濃度[17]。汪瑤等[18]也證實RFRP-3可作用于母豬促性腺激素釋放激素的神經元,抑制E2的產生與分泌。此外,RFRP-3處理垂體細胞還能夠抑制發情周期中豬促黃體生成素(luteinizing hormone, LH)的合成和分泌[19]。RFRP-3不僅可以調控豬生殖激素的分泌,還能夠通過降低CDK1和cyclinB1 mRNA的表達水平,抑制cyclinB-CDK1復合物的活化,使部分細胞阻滯在G2/M期,進而影響豬顆粒細胞的增殖[18]。綜上,目前RFRP-3關于豬的研究主要集中在對生殖激素和顆粒細胞的調控,還沒有關于RFRP-3對豬卵母細胞影響的研究,因此,本試驗通過在豬卵母細胞培養基中添加RFRP-3,探究其對體外培養豬卵母細胞成熟的影響,為揭示RFRP-3對哺乳動物繁殖的調控作用提供理論依據。
1 材料與方法
1.1 試驗試劑
TCM199培養基購自Gibco公司,人源RFRP-3購自Phoenix Pharmaceuticals公司,青霉素-鏈霉素溶液購自碧云天生物技術有限公司,馬絨毛膜促性腺激素(PMSG)和人絨毛膜促性腺激素(HCG)購自寧波第二激素廠,豬促成熟因子(MPF)和豬環磷酸腺苷(cAMP)檢測試劑盒購自睿信生物科技有限公司,反轉錄試劑盒購自北京全式金生物技術有限公司,細胞生長因子(EGF)、丙酮酸鈉、L-半胱氨酸、牛血清白蛋白(BSA)、D-葡萄糖、胰島素轉鐵蛋白(ITS)和礦物油均購自Sigma公司。
1.2 豬卵母細胞的采集與體外培養
豬卵巢采集于長沙市紅星盛業屠宰場,將卵巢置于含有2%雙抗的37 ℃的生理鹽水中,在2 h內運回實驗室。使用10 mL的注射器抽取卵巢上直徑約為3~6 mm的卵泡,靜置15 min后棄去上清,在顯微鏡下用自制口吸管挑取胞質均勻、且包被3層及以上顆粒細胞的卵丘-卵母細胞復合體(COCs),將COCs培養于含激素的TCM199成熟培養基(Ⅰ液:卵泡液、丙酮酸鈉、L-半胱氨酸、BSA、D-葡萄糖、孕馬血清促性腺激素、人絨毛膜促性腺激素、表皮細胞生長因子和胰島素轉鐵蛋白)中培養,培養條件為飽和濕度,38.5 ℃,5% CO2,培養22~24 h后換液,換成不含激素的TCM199成熟培養基(Ⅱ液:卵泡液、丙酮酸鈉、L-半胱氨酸、BSA、D-葡萄糖)繼續培養20~22 h。在培養基中添加10-6和10-8mol·L-1RFRP-3為RFRP-3處理組,不添加RFRP-3為空白對照組。
1.3 卵丘細胞的擴展與卵母細胞成熟的判定
體外培養豬卵母細胞44 h后,通過體式顯微鏡觀察卵丘擴展情況并計算卵丘擴展指數(cumulus expansion index, CEI),卵丘擴展指數=組內COCs卵丘擴展情況分值總和/組內所有COCs數。卵丘擴展分級:0級表示卵丘無擴展,計0分;1級表示僅最外圍1~2層卵丘細胞發生擴展,計2分;3級表示除放射冠之外所有卵丘細胞均發生擴展,計3分;4級表示所有卵丘細胞均發生擴展,計4分。
體外培養豬卵母細胞44 h后,用0.3%的透明質酸酶脫去卵丘顆粒層,以第一極體的排出作為COCs成熟的判定標準,并統計各組的第一極體排出率。
1.4 實時熒光定量PCR
從各組隨機挑取150個卵母細胞,使用微量RNA提取試劑盒(QIAGEN,德國)提取卵母細胞RNA,根據反轉錄試劑盒說明書將RNA反轉錄為cDNA。實時熒光定量反應體系為:5 μL TB Green Premix Ex Taq II, 0.2 μL ROX Reference Dye,正、反向引物各0.2 μL, cDNA 2 μL,DEPC水2.4 μL,每孔3個重復。反應條件:95 ℃ 30 s、95 ℃ 5 s、57 ℃ 30 s、95 ℃ 15 s、61 ℃ 60 s、95 ℃ 15 s、40個循環。各基因mRNA的相對表達量通過2-ΔΔCT法計算[20]。引物序列見表1。

表1 實時熒光定量PCR引物序列
1.5 ELISA檢測豬卵母細胞cAMP和MPF的含量
收集各組培養44 h的COCs,用0.3%透明質酸酶脫去顆粒層,使用蛋白酶溶解透明帶,經過PBS洗凈后,按照ELISA試劑盒說明書進行操作,檢測卵母細胞cAMP和MPF的含量。
1.6 放射免疫檢測雌二醇和孕酮的濃度
分別收集COCs培養22 和44 h的培養基,送北京北方生物技術研究所有限公司,通過放射免疫檢測雌二醇和孕酮的濃度。
1.7 數據分析
本試驗數據均采用SPSS19.0軟件進行分析,試驗結果用“平均數±標準誤(Mean±SEM)”表示,P<0.05表示差異具有統計學意義,P<0.01表示差異極顯著。
2 結 果
2.1 RFRP-3對豬COCs體外成熟率的影響
體外添加不同濃度(0、10-6和10-8mol·L-1)RFRP-3培養豬卵丘卵母細胞復合體44 h,然后脫去卵丘顆粒層,統計第一極體排出率。結果顯示,與對照組((71.8±3.39)%)相比,10-8mol·L-1RFRP-3添加組的成熟率((53±1.80)%)極顯著降低(P<0.01),但添加10-6mol·L-1RFRP-3對豬COCs的成熟率((69.27±0.35)%)無顯著影響(P>0.05)(圖1)。表明10-8mol·L-1RFRP-3可抑制豬COCs的體外成熟。
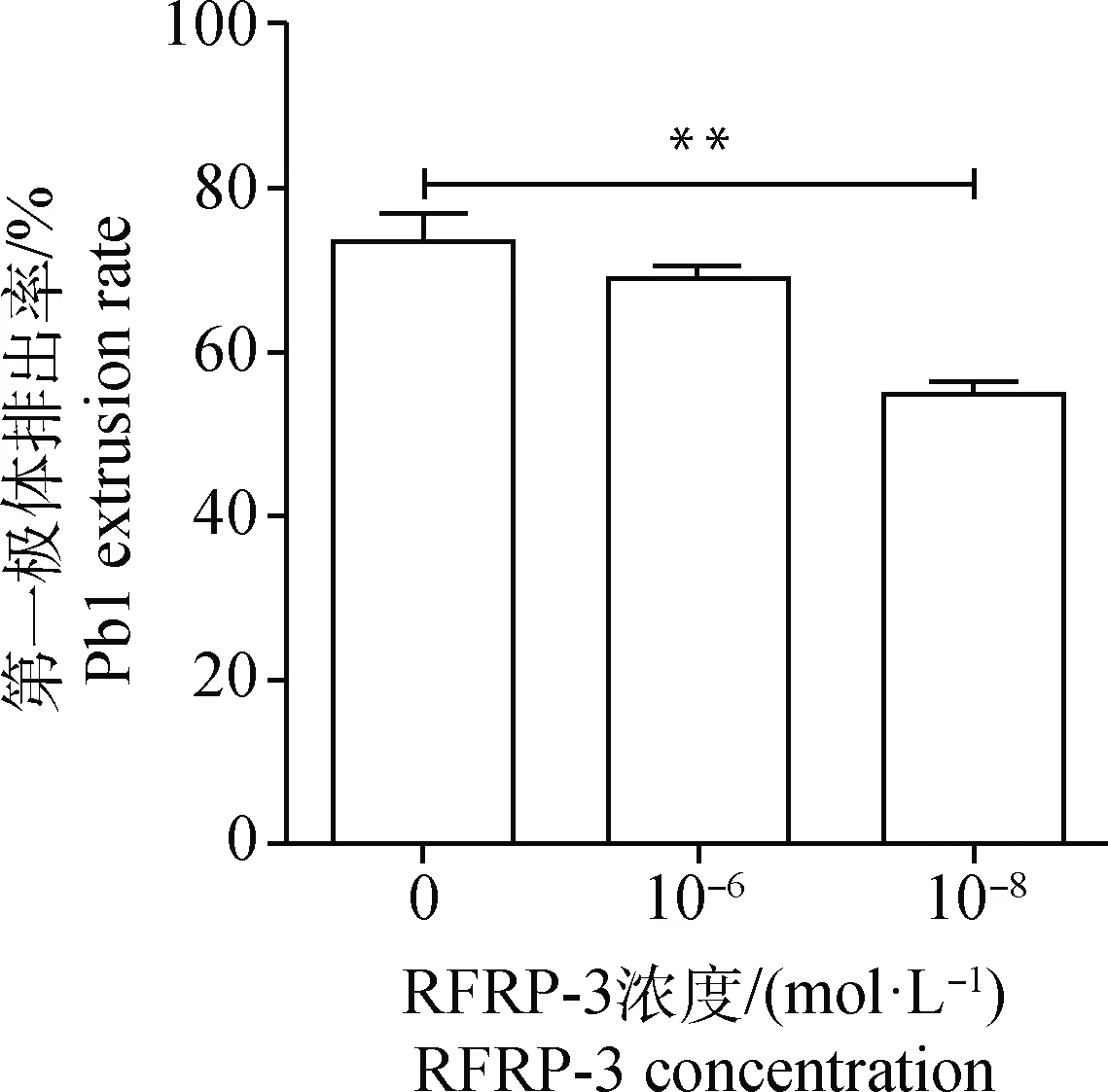
“*”表示與空白對照組相比差異顯著(P<0.05);“**”表示與空白對照組相比差異極顯著(P< 0.01),下同
2.2 RFRP-3對豬COCs卵丘擴展的影響
根據上述試驗結果,本研究后續試驗將選擇10-8mol·L-1RFRP-3作為RFRP-3處理組。添加10-8mol·L-1RFRP-3培養豬COCs 44 h后,通過顯微鏡觀察各組的卵丘擴展情況并計算卵丘擴展指數。結果顯示,RFRP-3添加組的卵丘擴展指數(3.46±0.1)與對照組(3.24±0.08)相比無明顯變化(P>0.05)(圖2A-E)。采用qRT-PCR檢測RFRP-3添加組中卵丘擴展相關因子PTGS2、HAS2和PTX3基因的表達水平。結果如圖2F所示,10-8mol·L-1RFRP-3處理COCs可極顯著下調PTGS2、HAS2和PTX3基因的表達水平(P<0.01)。表明添加RFRP-3對豬COCs卵丘擴展無影響,但可降低相關卵丘擴展因子的表達水平。
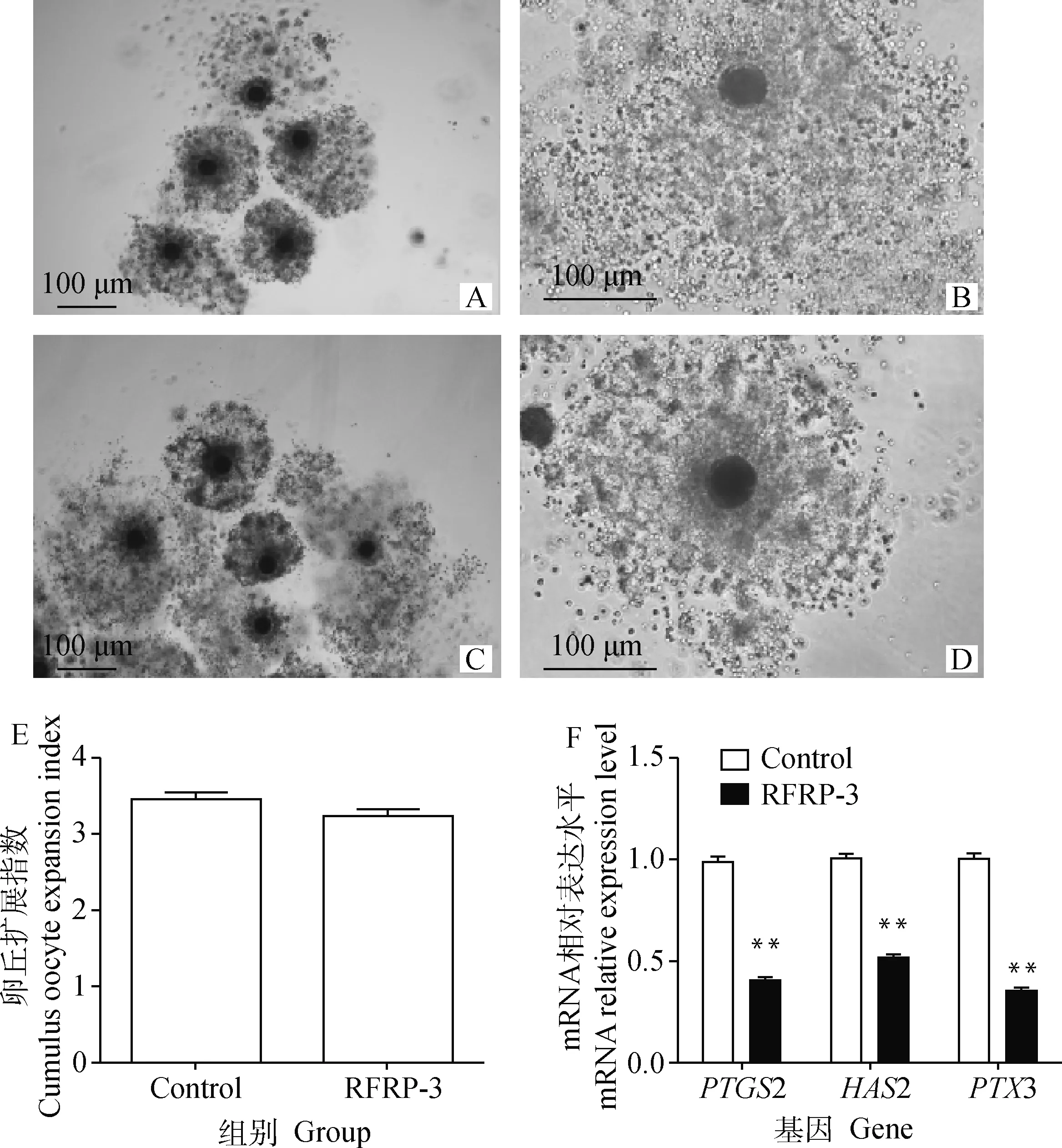
A.Control組卵丘細胞擴展情況(40×);B.Control組卵丘細胞擴展情況(100×);C.RFRP-3組卵丘細胞擴展情況(40×);D.RFRP-3組卵丘細胞擴展情況(100×);E.RFRP-3對卵丘擴展的影響;F.RFRP-3對卵丘擴展因子表達的影響
2.3 RFRP-3對豬卵母細胞成熟相關因子的影響
為進一步探索RFRP-3抑制豬COCs體外成熟的機制,添加RFRP-3培養COCs成熟后,脫去卵丘顆粒層,收集卵母細胞裂解物,采用ELISA檢測卵母細胞胞質中MPF和cAMP的含量。結果表明,RFRP-3添加組與空白對照組相比可極顯著降低細胞質中MPF的含量(P<0.01),但對cAMP的含量無明顯影響(P>0.05)(圖3A,3B)。采用qRT-PCR檢測添加RFRP-3培養COCs后對GDF9和BMP15表達的影響。結果如圖3C所示,與對照組相比,添加RFRP-3可極顯著提高卵母細胞GDF9基因的表達(P<0.01),顯著提高BMP15的表達(P<0.05)。

A.MPF水平;B.環磷酸腺苷水平;C.生長分化因子9和骨形態發生蛋白15基因的表達
2.4 RFRP-3對豬卵母細胞周期相關基因的影響
添加RFRP-3培養COCs成熟后,脫去卵丘顆粒層,收集卵母細胞裂解物,利用qRT-PCR檢測豬卵母細胞中細胞周期相關基因CCNB1和CDK1的表達水平。結果表明,與對照組相比,添加RFRP-3培養豬COCs可以顯著降低卵母細胞中CCNB1和CDK1基因的表達水平(P<0.05,圖4)。
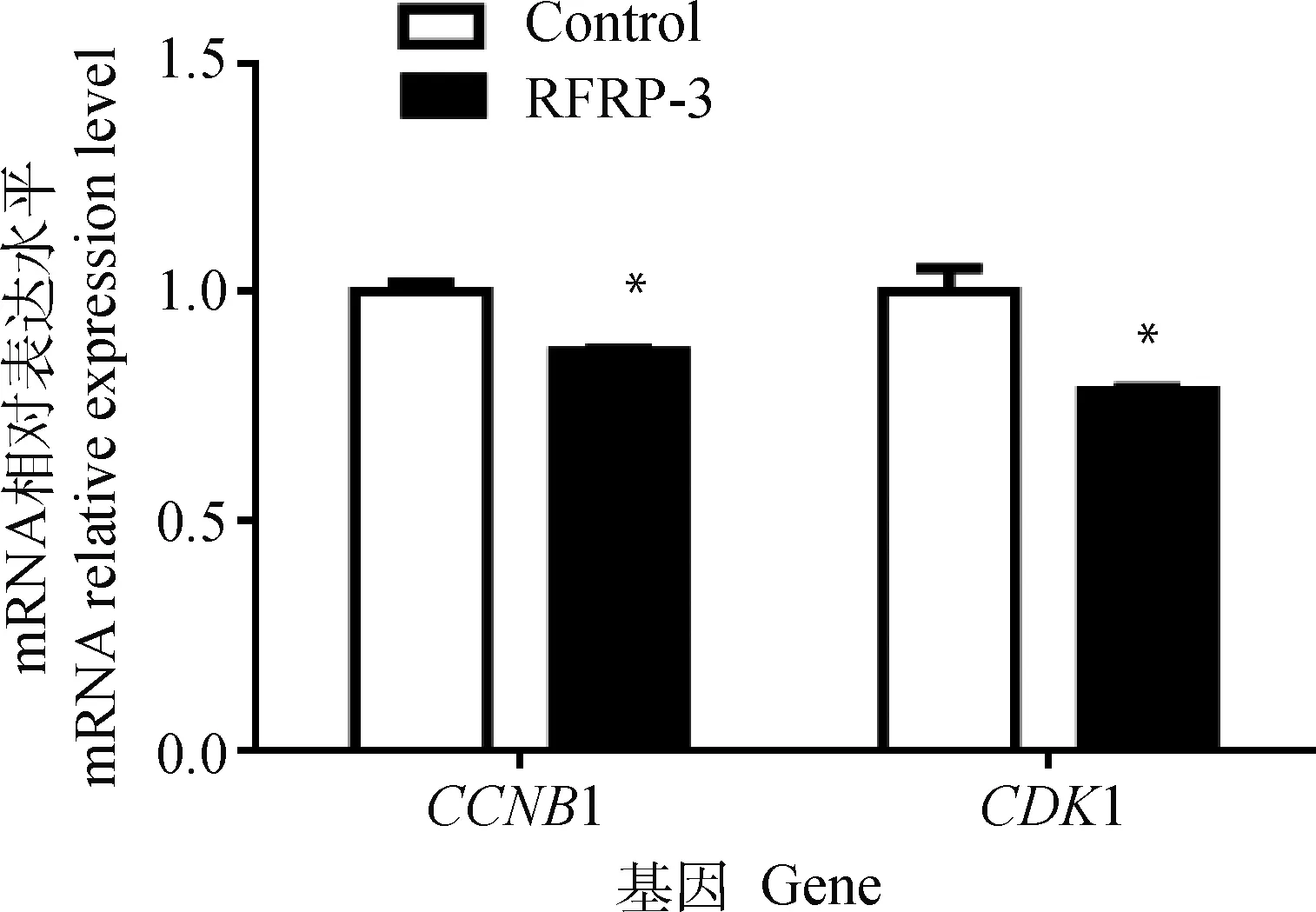
圖4 RFRP-3對豬卵母細胞周期相關基因表達的影響
2.5 RFRP-3對豬COCs激素分泌的影響
添加RFRP-3培養COCs后,分別收集22(Ⅰ液)和44 h(Ⅱ液)的培養基,檢測培養基中雌二醇和孕酮的濃度。結果表明,RFRP-3可極顯著降低培養基中雌二醇(E2)和孕酮(P4)的濃度(P<0.01)(圖5)。

A.雌二醇濃度;B.孕酮濃度
3 討 論
在哺乳動物卵巢中,RFRP-3主要表達在顆粒細胞和黃體細胞中,在卵母細胞中的表達研究較少。Wang等[21]研究表明,添加10-6和10-8mol·L-1RFRP-3可顯著降低豬顆粒細胞cyclin B1和CDK1的表達水平。cyclin B和CDK1可以組成異二聚復合體(即成熟促進因子,MPF),MPF對于卵母細胞的減數分裂恢復是必不可少的,因此本試驗選擇添加0、10-6和10-8mol·L-1RFRP-3體外培養豬COCs,結果顯示,添加10-8mol·L-1RFRP-3可極顯著降低卵母細胞的成熟率,而添加10-6mol·L-1RFRP-3對豬COCs的體外成熟率無顯著影響。Singh等[22]用RFRP-3處理小鼠,也表明RFRP-3可以明顯抑制卵泡發育,使得卵母細胞無法成熟。卵母細胞成熟的實質就是減數分裂的阻滯與恢復,該過程主要由cAMP和MPF調控。研究表明,高水平的cAMP可以抑制cyclin B的表達,磷酸化CDK1的蘇氨酸Thr-14和酪氨酸Tyr-15殘基,導致CDK/cyclin B 蛋白復合物失活,MPF的活性被抑制[23],但隨著卵母細胞的發育成熟,cAMP被cAMP磷酸二酯酶降解,cAMP濃度降低,誘導MPF活化,引起卵母細胞減數分裂恢復[24]。郭振偉等[25]體外添加褪黑素培養水牛卵母細胞發現,褪黑素能夠抑制cAMP合成, 進而促進卵母細胞成熟。本研究利用ELISA法檢測了豬卵母細胞cAMP和MPF的濃度,結果發現,RFRP-3可以使卵母細胞中MPF的濃度極顯著降低,但cAMP的濃度無明顯變化,推測正是這一變化導致卵母細胞減數分裂停滯,抑制卵母細胞的成熟。李贊[26]的研究也表明,添加減數分裂的抑制劑AMH,可降低MPF的濃度,維持高水平的cAMP,阻礙卵母細胞成熟。本試驗通過qRT-PCR檢測發現,卵母細胞CCNB1和CDK1的基因表達水平顯著下降,推測RFRP-3可能是通過降低CCNB1和CDK1基因的表達,從而下調了MPF。與本研究結果類似,Wang等[21]研究表明,添加RFRP-3可以降低豬顆粒細胞cyclinB和CDK1的表達水平,抑制cyclin B-CDK1復合物(MPF)的激活從而誘導細胞G2/M期停滯,阻礙豬顆粒細胞增殖。推測RFRP-3可能通過調節MPF和cAMP的水平,進而調控卵母細胞的成熟。
GDF9和BMP15作為卵母細胞分泌因子,可以與卵丘顆粒細胞相互作用,促進卵丘擴展,調控卵母細胞成熟[27]。魏莉娜等[28]研究發現,卵母細胞中GDF9和BMP15參與調控卵母細胞的成熟。在本試驗中,RFRP-3顯著抑制了豬卵母細胞的成熟,但RFRP-3處理組中卵母細胞GDF9和BMP15的基因表達水平卻顯著升高。有研究表明,體外添加高濃度(300 ng·mL-1)的GDF9可以顯著降低卵母細胞的成熟率[29],過表達BMP15可以阻礙卵母細胞的成熟[30],據此推測,體外添加RFRP-3可能通過增加卵母細胞內的GDF9和BMP15表達,進而抑制豬卵母細胞的成熟。
卵母細胞的成熟與排卵受多種細胞和信號因子的調控,其中卵丘細胞通過復雜的縫隙連接為卵母細胞提供部分營養物質,維持卵母細胞減數分裂阻滯和恢復,促進卵母細胞成熟[31]。趙子墨等[32]研究證明,培養COCs的存活率與成熟率顯著高于裸卵組,表明卵丘細胞對于卵母細胞的成熟是必不可少的。其中卵丘細胞擴展在卵母細胞成熟過程中發揮著重要作用,卵丘擴展后卵丘細胞才能與卵母細胞進行營養傳遞與信息交換。本試驗結果表明,在豬COCs體外培養體系中添加RFRP-3對卵丘細胞的擴展無明顯影響。Nikoloff等[33]研究也證明,添加不飽和脂肪酸EPA,體外培養COCs雖然可以顯著降低卵母細胞的成熟率,但對卵丘細胞擴展無明顯影響,這一點與本研究結果一致。卵丘擴展需要卵丘基質來維持,卵丘基質的合成依賴于卵丘擴展因子[34]。Pan等[35]研究發現,在卵母細胞培養基中添加E2可以增強卵丘擴展因子(PTGS2、HAS2和PTX3)基因的表達,從而促進卵母細胞成熟。Nagyova等[36]研究證明,添加10 μmol·L-1表皮生長因子受體酪氨酸激酶的抑制劑(lapatinib),可以使卵丘擴展因子TNFAIP6和PTGS2的表達顯著降低,阻礙豬卵母細胞成熟。本試驗結果顯示,RFRP-3可通過抑制卵丘擴展因子(PTGS2、HAS2和PTX3)的表達來降低卵母細胞的成熟。研究表明,卵丘顆粒細胞不僅通過卵丘擴展因子影響卵母細胞的成熟,顆粒細胞分泌的E2和P4也可以維持卵母細胞減數分裂的停滯與恢復,促進卵母細胞的成熟[37-38]。本研究結果顯示,添加RFRP-3體外培養卵母細胞,可極顯著降低培養基中E2和P4的濃度,表明RFRP-3可通過抑制卵丘顆粒細胞分泌E2和P4,進而抑制卵母細胞的成熟。
4 結 論
RFRP-3可以通過降低豬COCs卵丘擴展因子(PTGS2、HAS2和PTX3)、周期相關因子(CCNB1和CDK1)的基因表達,增加卵母細胞分泌因子(GDF9和BMP15)的基因表達,抑制卵母細胞MPF的生成,抑制卵丘顆粒細胞雌二醇和孕酮的分泌,進而抑制豬卵母細胞的成熟。本研究可以為揭示RFRP-3對哺乳動物卵母細胞的調控作用及機制提供理論依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
