文蛤與微藻協同作用對水體氮、磷消除效果研究
2022-03-20 12:20:20張志東陳愛華吳楊平陳素華李秋潔
水產科學 2022年2期
張志東,陳愛華,吳楊平,張 雨,曹 奕,陳素華,田 鎮,李秋潔
( 江蘇省海洋水產研究所,江蘇省文蛤良種場,江蘇省海洋經濟貝類研發中心,江蘇 南通 226007 )
微藻為形體微小且只能在顯微鏡下才能分辨其形態的藻類總稱[1]。其種類繁多,在海洋、湖泊、河流、池塘及水庫等各類水體中均有分布。目前已探知的微藻中,絕大部分營自養生活,其可通過自身的光能轉化作用直接從水體中吸收氮、磷元素,并通過氧化磷酸化作用合成腺苷三磷酸為細胞供能,完成生長繁殖[2-3]。在此過程中產生的氧氣可改善水質,同時自身又可作為水產動物的餌料,提高水產品品質[4]。因此,微藻在水產生態養殖及其水環境調控中具有重要作用,利用微藻降低或消除富營養化水體中的氮、磷等元素含量具有極大潛力。然而,當水體中氮、磷超過一定閾值,水體中微藻將大量繁殖,導致水華,從而破壞水域生態系統平衡。針對微藻過量繁殖的現象,實驗室常用固定化微藻技術控制微藻繁殖,達到水質凈化的目的[3,5]。然而該技術成本高,難以實現推廣,因此在實際生產中仍用濾食性貝類攝食過量微藻,使水體中微藻數量保持在適宜的范圍[6-7]。
貝類擁有強大的濾食能力,對水體中藻相結構產生重要影響。有研究顯示,在魚池內混養三角帆蚌(Hyriopsiscumingii)可控制浮游藻類數量,促進養殖水體中大型藻類的生長,進而提高養殖水體的生物總量[7]。朱小龍等[8]研究發現,河蜆(Corbiculafluminea)能有效抑制富營養化湖泊中藻類暴發現象。方磊等[9]認為,貝類如果過量攝食會產生大量“假糞”,此過程雖無法完全消除藻類,但經過貝類消化道的藻類,被貝類黏液包裹,其生長繁殖受到一定抑制。即便假糞中的藻類被再次攪動激活,也只有其中一部分能被激活。因此混養濾食性貝類對提高池塘養殖產量、池塘養殖模式優化具有積極意義。然而目前有關濾食性貝類去除水體氮、磷的研究均在海洋、湖泊、池塘等大水域環境下進行,忽略了水域中微藻的作用。單獨放養貝類及在有藻和無藻的環境中對水體中氮、磷營養鹽的消除效果研究尚未見報道。因此,筆者以文蛤(Meretrixmeretrix)為研究對象,比較其在有藻和無藻情況下對水體中無機氮及無機磷的消除差異,并通過響應面法優化出文蛤和微藻的最佳的搭配條件,旨在為水質調控及池塘養殖技術優化提供參考。
1 材料與方法
1.1 試驗材料
試驗所用文蛤與微藻均來自江蘇省文蛤良種場,其中文蛤為紅殼色選育系子四代,自養殖池塘取出后在室內砂濾海水暫養7 d,日投喂1×105個/mL的球等鞭金藻(Isochrysisgalbana)1次,并換水1次。試驗前2 d停止投喂,挑選健康、規格相近個體進行試驗,平均殼長為(25.91±2.76) mm,平均體質量為(4.30±1.08) g。試驗用水為二次煮沸的砂濾海水添加硝酸鈉、氯化銨及磷酸二氫鉀配制。氮、磷初始指標為:無機氮1.65 mg/L、無機磷0.23 mg/L。底泥采于江蘇呂四灘涂,運回實驗室后,用烘箱烘干,用于試驗。
1.2 試驗設計
1.2.1 定性試驗
試驗在12個68 cm×46 cm×35 cm聚乙烯水槽中進行,設計有貝無藻組(A組)、無貝有藻組(B組)及有貝有藻組(AB組)3個試驗組,以無貝無藻組(O組)為對照組,每組設3個平行。文蛤放養密度為225個/m2。有藻試驗組初始投喂球等鞭金藻約為1.6×106個/mL,待貝類攝食效率與微藻生長效率基本持平后不再投喂,微藻密度基本保持穩定。試驗為靜水試驗,每個水槽中平鋪100 g底泥。在試驗開始后3、6、12、24、48、72、96 h各取300 mL水樣,4%甲醛固定,用于水質指標的測定。
1.2.2 定量試驗
在定性試驗的基礎上發現,有貝有藻組對水體無機氮及無機磷吸收效果較好。因此針對有貝有藻組結果進行定量化分析,按照中心組合設計(CCD)設置13組多因素試驗(表1),通過響應曲面設計方法(RSM)建立回歸模型,并對回歸方程進行方差分析及顯著性檢驗。以水體無機氮、無機磷日消除率(RIN和RIP)為指標,運用中心組合設計模型,選擇雙因素三水平的中心組合方案,以文蛤密度和微藻密度(所用藻類為球等鞭金藻)為考察因素,進行響應面試驗,試驗因素水平及結果見表1,試驗共持續4 d,試驗裝置及方法同定性試驗有貝有藻組。
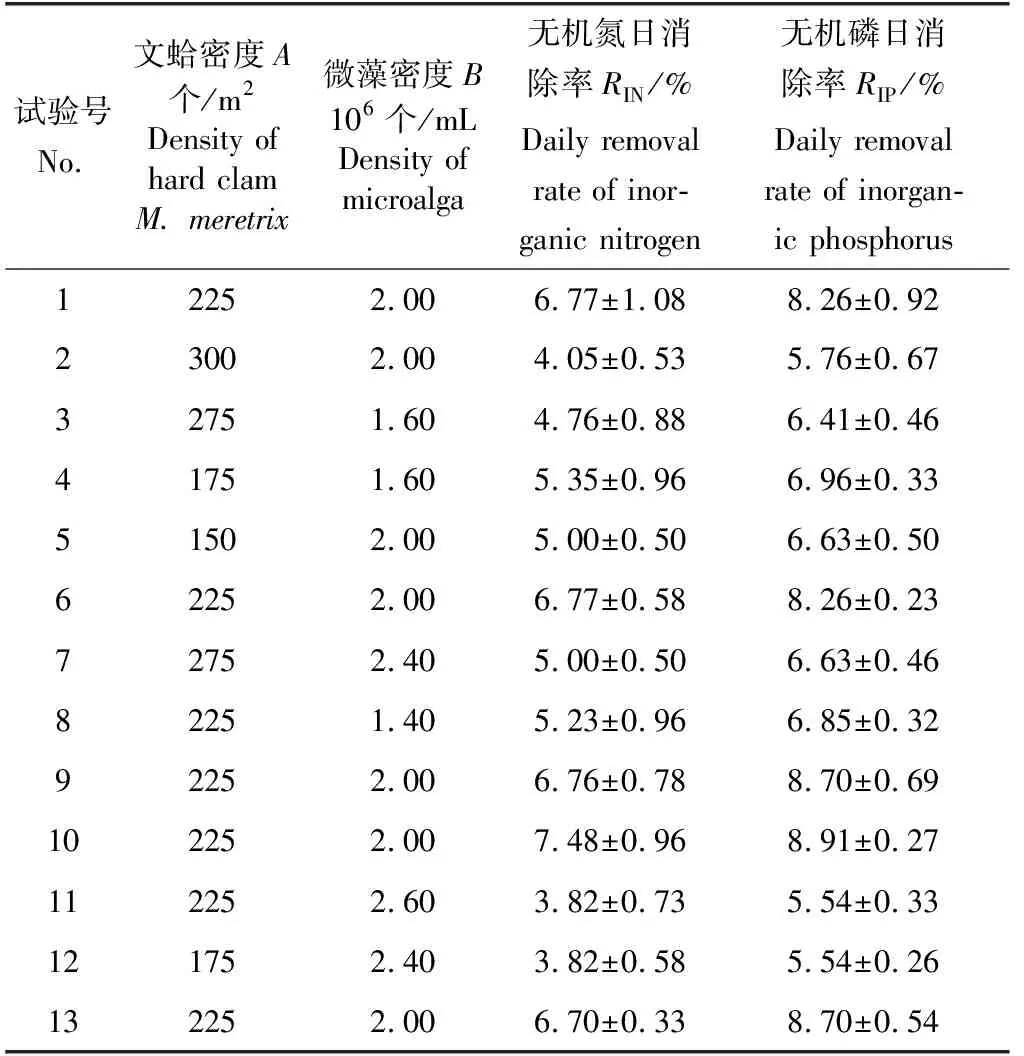
表1 試驗設計與結果Tab.1 Central-Composite design with the observed responses
1.3 水體氮、磷的測定與計算
試驗測定的水質指標主要包括水體中氨氮(NH4+-N)、亞硝態氮(NO2--N)、硝態氮(NO3--N)、無機氮(IN)、無機磷(IP)。氨氮通過次溴酸鹽氧化法測定,亞硝態氮通過磺胺—鹽酸萘乙二胺法測定,硝態氮采用鋅鎘還原法測定,無機磷通過鉬藍分光光度法測定;無機氮質量濃度(ρIN)及無機氮日消除率(RIN)及無機磷日消除率(RIP)計算公式如下:
ρIN=ρ1+ρ2+ρ3
RIN=(ρIN, 0-ρIN, t)/(ρIN, 0×t)×100%
RIP=(ρIP, 0-ρIP, t)/(ρIP, 0×t)×100%
式中,ρIN為水體無機氮質量濃度(mg/L),ρ1、ρ2及ρ3分別為水體氨氮、亞硝態氮及硝態氮質量濃度(mg/L),ρIN, 0及ρIP, 0為無機氮及無機磷初始質量濃度(mg/L),t為試驗時間(d),ρIN, t及ρIP, t為在t時間無機氮及無機磷質量濃度(mg/L)。
1.4 數據統計
定性試驗數據采用SPSS 19.0進行單因素方差分析,用Origin 8.0作圖。定量試驗根據Design-expert 8.0軟件中中心組合設計模型設置試驗組,用該軟件進行二元二次多項式回歸分析和方差分析。
2 結 果
2.1 文蛤及微藻對水體無機氮及無機磷質量濃度的影響
由圖1可見,隨著試驗的進行,有貝無藻組氨氮質量濃度較無貝無藻組無顯著變化(P>0.05);無貝有藻組和有貝有藻組氨氮和無機氮質量濃度顯著低于無貝無藻組(P<0.05)。有貝無藻組和無貝有藻組亞硝態氮質量濃度隨著時間的延長,有明顯升高趨勢(P<0.05),而有貝有藻組顯著低于有貝無藻組和無貝有藻組(P<0.05)。試驗結束時,有貝無藻組和有貝有藻組硝態氮質量濃度顯著低于無貝無藻組和無貝有藻組(P<0.05)。由圖2可見,隨著時間的延長,有貝無藻組無機磷質量濃度無明顯變化,無貝有藻組和有貝有藻組無機磷質量濃度較無貝無藻組有明顯下降趨勢(P<0.05),而無貝有藻組和有貝有藻組之間差異顯著(P<0.05)。
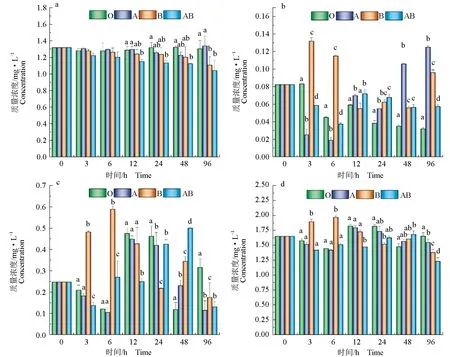
圖1 文蛤及微藻對水體無機氮質量濃度影響Tab.1 Effects of hard clam M. meretrix and microalga on concentration of inorganic nitrogen in watera.氨氮; b.亞硝態氮; c.硝態氮; d.無機氮; O.無貝無藻組; A.有貝無藻組; B.無貝有藻組; AB.有貝有藻組;柱狀圖上方不同的小寫字母表示同一時間不同試驗組之間差異顯著(P<0.05);下同.a.ammonia nitrogen; b.nitrite; c.nitrate; d.inorganic nitrogen; O.no clam without algae group; A.clam without algae group; B.algae without clam group; AB.clam with algae group; different letters above the bar chart indicate significant differences between different groups at the same time (P<0.05); et sequentia.
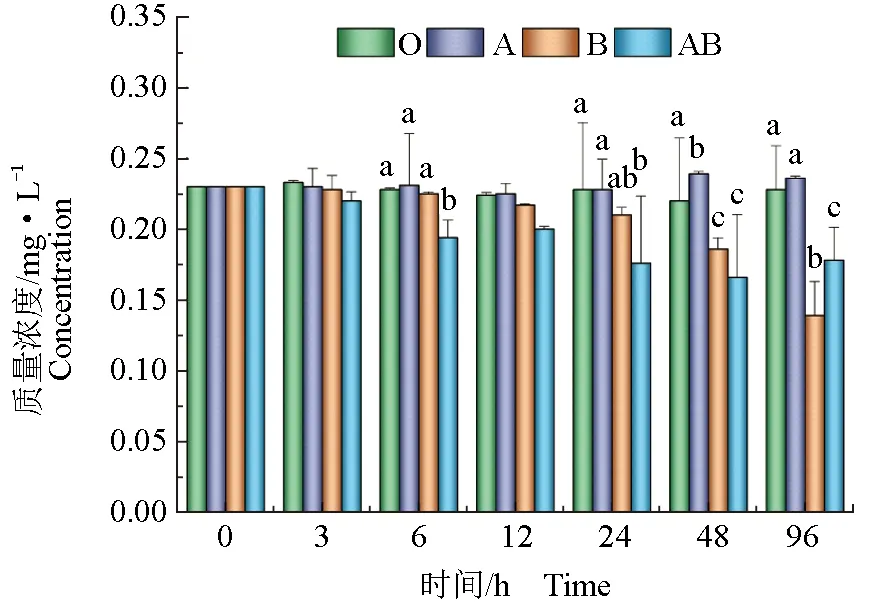
圖2 文蛤及微藻對水體無機磷質量濃度影響Tab.2 Effects of hard clam M. meretrix and microalgae on concentration of inorganic phosphorus in water
2.2 文蛤及微藻對水體無機氮和無機磷日消除率響應面法分析
定量試驗結果顯示(表2、表3),水體無機氮日消除率及無機磷消除率回歸模型的F值分別為26.81和32.30,P<0.01,表明中心組合設計優化法可靠,文蛤密度和微藻密度對無機氮消除率及無機磷消除率的影響顯著,模型具有統計學意義。其次,兩個模型的擬合系數(r2)分別為0.9504和0.9585,說明模型預測值與實測值擬合度高。校正決定系數(Adjr2)分別為0.9149和0.9288,均可解釋數據變異性的90%以上。精密度分別為11.992%和13.087%,說明回歸模型可以準確反映結果。另外,文蛤密度一次效應對水體無機氮及無機磷日消除率影響不顯著(P>0.05);微藻密度一次效應對水體無機氮及無機磷日消除率影響顯著(P<0.05);文蛤密度和微藻密度交互效應對水體無機氮及無機磷日消除率影響顯著(P<0.05);文蛤密度和微藻密度二次效應對水體無機氮及無機磷日消除率影響極顯著(P<0.01)。根據方差分析結果得出水體無機氮及無機磷消除率二次多項回歸方程為:
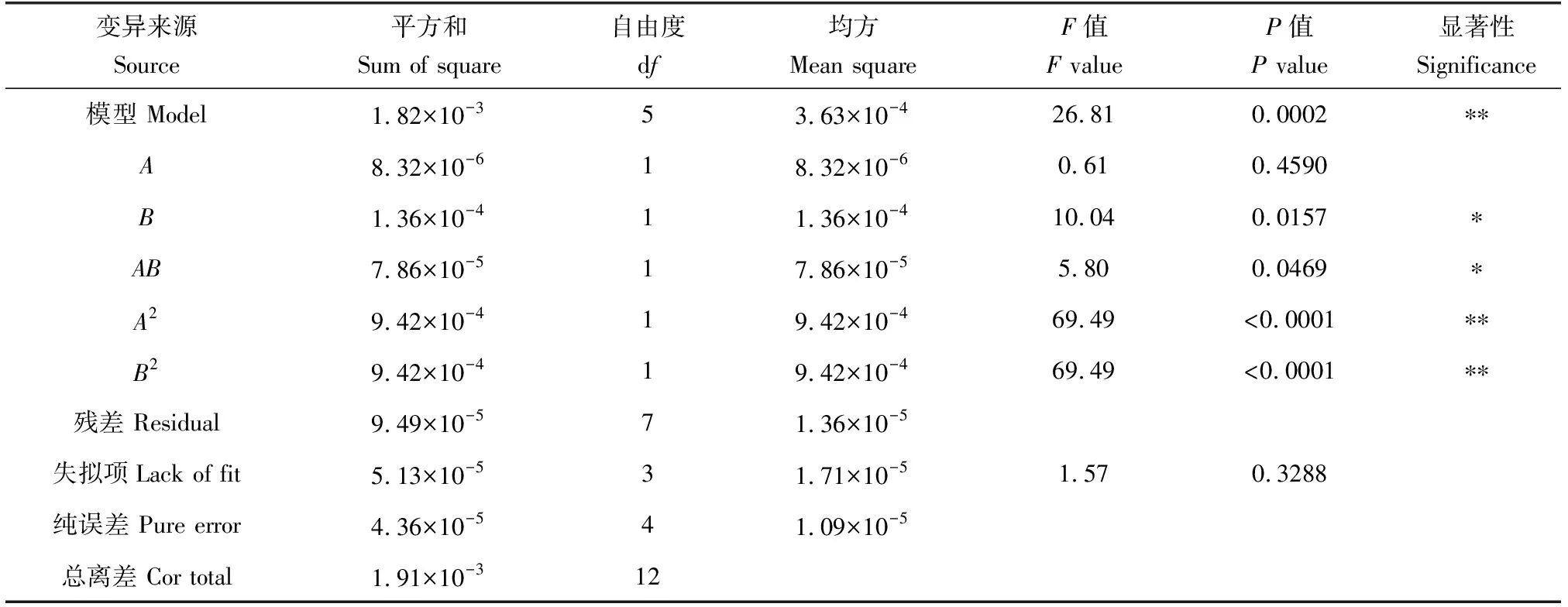
表2 回歸模型方差分析(RIN)Tab.2 ANOVA for the regression models (RIN)
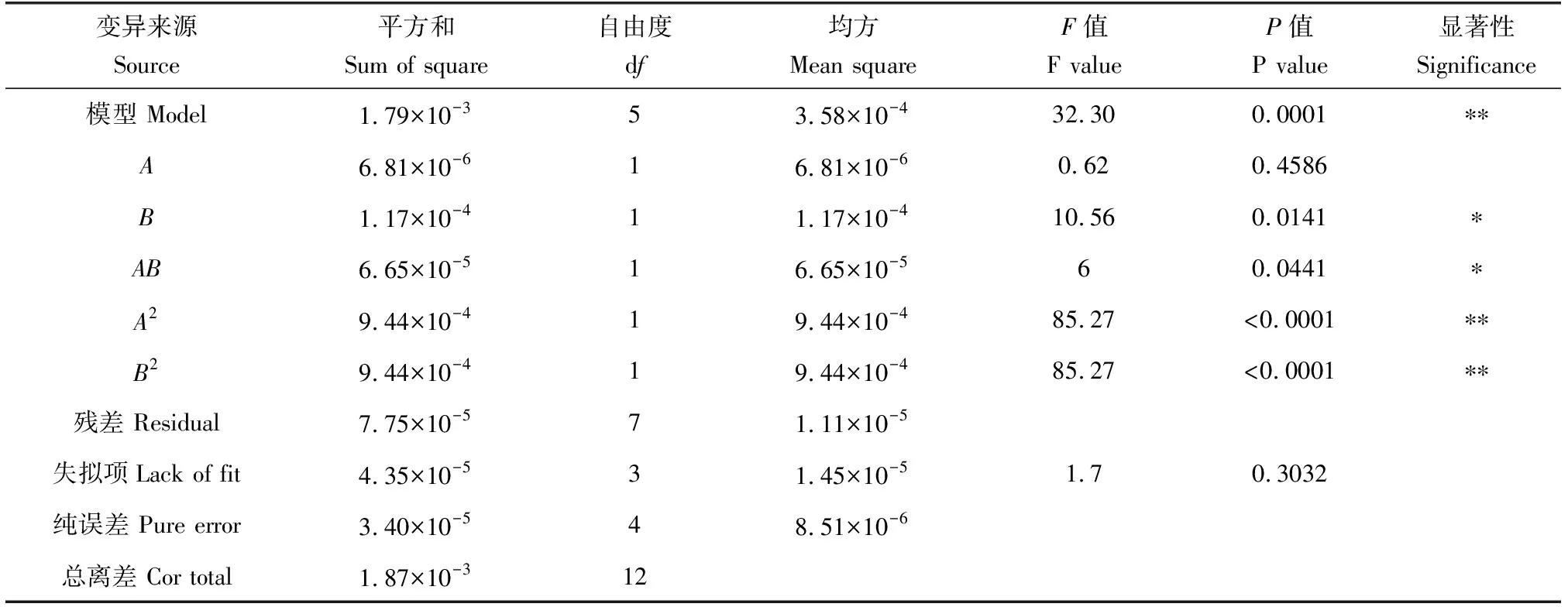
表3 回歸模型方差分析(RIP)Tab.3 ANOVA for the regression models (RIP)
水體無機氮日消除率(RIN)預測值對文蛤密度(A)和微藻密度(B)的二次多項回歸方程為:
RIN=-0.29+1.46×10-3A+2.07×10-3B+2.22×10-6AB-4.26×10-6A2-6.63×10-6B2
水體無機磷日消除率(RIP)預測值對文蛤密度(A)和微藻密度(B)的二次多項回歸方程為:
RIP=-0.28+1.50×10-3A+2.12×10-3B+2.04×10-6AB-4.27×10-6A2-6.67×10-6B2
根據上述公式所建立的數學模型及響應面結果見圖3,水體無機氮及無機磷日消除率的最佳條件為:文蛤密度221個/m3,微藻密度1.92×106個/mL,此條件水體無機氮及無機磷消除率分別為6.93%和8.60%。
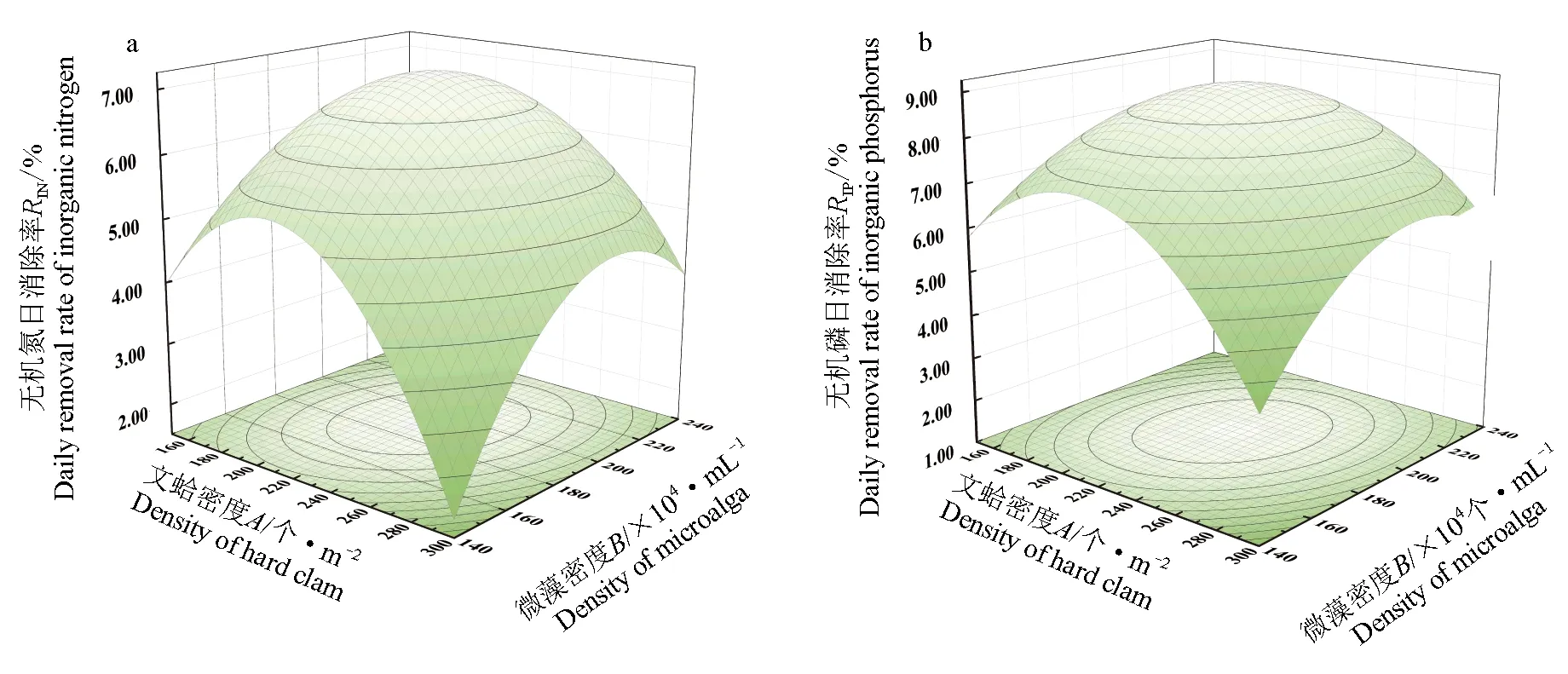
圖3 交互作用對水體無機氮和無機磷日消除率影響的響應面分析Fig.3 Response surface showing interactive effects of factors affecting daily removal rates of inorganic nitrogen and phosphorus
2.3 模型驗證
通過中心組合設計構建的響應面模型獲得了水體無機氮和無機磷日消除率最大時文蛤和微藻的密度搭配比例。通過實際試驗對模型擬合結果進行驗證。預測結果與實際結果見表4,驗證結果表明該模型分析結果準確。

表4 試驗結果驗證Tab.4 Verification of experimental results
3 討 論
3.1 文蛤對水體無機氮及無機磷含量的影響
貝類的濾食、排泄、管道筑建及穴居等生理活動對養殖水體會產生重要影響[10]。本試驗結果顯示,單放文蛤組(有貝無藻組)對水體無機氮及無機磷含量影響不顯著(P<0.05),這可能與文蛤在清水態(無藻)環境中與外界物質交換量減少、濾水率下降有關。另外,在清水態環境下,文蛤長期處于饑餓脅迫之中,排氨率明顯上升。這可能也是水體無機氮居高不下的原因之一[11]。因此,在文蛤養殖過程中應及時注入“肥水”,使之與外界環境之間有充足的物質交換,從而有利于文蛤對氮、磷的吸收。在投喂微藻后(有貝有藻組),水體中無機氮及無機磷含量顯著降低(P<0.05),且文蛤密度和微藻密度的交互效應顯著(P<0.05),據此推測文蛤與微藻對富營養化水體調控存在一定的正反饋效應。貝類能降低水體氮、磷含量,一方面貝類對水體中微藻和懸浮物高強度濾食,另一方面貝類對底層藻類的生長具有促進作用,抑制了沉積物營養釋放,從而間接降低水體氮、磷含量[10]。但貝類放養密度不宜過高,放養密度過高時,貝類會形成大量假糞,通過微生物作用及自身的生物擾動,加快系統的礦化反應,向水體中釋放大量無機氮及無機磷[12]。本試驗結果顯示,文蛤放養密度約221個/m2時,對水體無機氮及無機磷日消除率達到最大,不足或超過此放養密度時,無機氮及無機磷消除效果均一般,與上述規律一致。
3.2 微藻對水體無機氮及無機磷含量的影響
微藻在水生生態系統中起著重要作用。它能吸收水體中無機氮和無機磷,并通過光能轉化作用將其變為自身的組成物質,完成元素的轉移[13]。本試驗結果顯示,單放微藻組(無貝有藻組)能顯著降低水體中無機氮(主要是氨氮)和無機磷含量(P<0.05),與上述規律一致。而無貝有藻組亞硝態氮含量有明顯上升趨勢(P<0.05),說明藻類對水體中亞硝態氮利用效率很低。McCarthy等[14]研究認為,水體中藻類利用各種形式氮的優先順序為氨氮>硝態氮>亞硝態氮,其原因是同化亞硝態氮的過程需要更多的能量[14-15]。竇勇等[16-17]在研究威氏海鏈藻(Thalassiosiraweissflogii)及小球藻(Chlorellavulgaris)氮、磷吸收情況時,發現相似規律。而微藻對水體中無機氮及無機磷的吸收效果與其密度有關,當水生生態系統中微藻密度很高時,一方面,其自身生長受到抑制,氮、磷吸收效率降低,另一方面,其生物固氮作用加強,從而將氮氣還原為氨氮,使水體中無機氮含量未降反升[18]。而且微藻密度過高時,會影響生態系統的穩定性。因此,本試驗通過響應面法優化出微藻的密度約為192萬個/mL時,對水體的氮、磷消除率達到最大,與上述規律一致。常用濾食性貝類可以控制水體微藻密度過高現象,防止水華的發生。
本試驗通過響應面法擬合出文蛤和藻類搭配的最佳比例為:文蛤密度221 個/m2、微藻密度192萬個/mL,此時對水體無機氮及無機磷日消除率達到最大,分別為6.93%和8.60%。由于實際操作過程中無法做到如此精細,故文蛤密度約225個/m2(15萬個/畝),微藻密度約200萬個/mL即可。模型驗證可靠,可用于實際生產中。另外,在實際生產中,分析文蛤及微藻對無機氮及無機磷消除效果時,還應充分考慮溫度、鹽度及光照度等因素[19]。
4 結 論
在清水態環境(無藻)中,文蛤對水體無機氮及無機磷消除效果有限,需添加微藻與其協同作用,才可降低水體無機氮及無機磷含量。在實際生產中,文蛤密度約225 個/m2(15萬個/畝),微藻密度約為200萬個/mL,可對水體無機氮及無機磷達到最大消除效果,此時無機氮及無機磷日消除率分別為(6.90±0.33)%和(8.57±0.29)%。
