lncRNA GAS5靶向調控miR-136對高糖誘導的人腎小管上皮細胞中炎性和纖維化因子表達影響及機制研究
2022-03-16 09:22:56宿晶王吉磊劉世賓
河北醫藥 2022年4期
宿晶 王吉磊 劉世賓
腎間質纖維化(RIF)是糖尿病腎病(DN)最常見特征之一,其在糖尿病腎損傷早期出現,是DN等慢性腎病發展為終末期腎病的最終共同途徑。因此,探討RIF進展的分子機制對開發有效的DN治療策略意義重大。長鏈非編碼RNA(lncRNA)定義為一類長度超過200核苷酸蛋白編碼潛能缺失的RNA轉錄本,其可在轉錄和轉錄后水平調節基因表達廣泛參與表觀遺傳調控、細胞周期、細胞分化、凋亡等生物學過程[1]。lncRNAs的異常表達與包括DN在內的多種疾病進展有關[2]。生長抑制特異性轉錄本5(GAS5)是一種lncRNA,研究指出2型糖尿病患者外周血單核細胞中GAS5表達顯著升高[3]。淋巴管內皮細胞中GAS5的表達差異可能與糖尿病相關并發癥發病機制有關[4]。然而GAS5在RIF中的作用和分子機制鮮有報道。miR-136是一種與多種病理過程相關的非編碼RNA,研究指出過表達miR-136顯著抑制腎癌、肺癌細胞的上皮間質轉化進程[5,6]。本研究通過生物信息學分析發現miR-136是GAS5的潛在靶基因,于是推測GAS5可能通過靶向miR-136參與調控RIF進程。因此,本研究以高糖誘導的人腎小管上皮細胞(HK-2)建立RIF體外細胞模型[7],探討GAS5靶向miR-136對高糖誘導的HK-2細胞炎性和纖維化因子表達的影響,以期為改善RIF、延緩DN進展提供有效靶點。
1 材料與方法
1.1 實驗材料 人腎小管上皮細胞HK-2購于中國科學院上海細胞庫;miR-136模擬物(mimics)、抑制物(anti-miR-136)以及相應對照(miR-NC、anti-miR-NC),GAS5小干擾RNA(Si-GAS5)、過表達質粒(pcDNA-GAS5)以及相應對照(Si-NC、pcDNA),熒光素酶報告載體購自上海吉瑪制藥公司;PrimeScript逆轉錄Master Mix購于大連TAKARA公司;Power SYBR Green PCR Master Mix購于美國ABI公司;IL-6、TNF-α酶聯吸附測定(ELISA)試劑盒購于上海康朗生物公司;兔源肌動蛋白α(α-SMA)、纖維連接蛋白(FN1)、磷酸甘油醛脫氫酶(GAPDH)抗體以及山羊抗兔IgG二抗購于北京博奧森生物公司。
1.2 方法
1.2.1 細胞培養與模型構建:HK-2細胞采用含10%胎牛血清的低糖DMEM培養基,在37℃、CO2濃度為5%的細胞培養箱中培養。當細胞密度達到90%時,加入胰酶消化后按照1∶3比例接種新的培養瓶進行傳代。用含30 mmol/L葡萄糖的培養液孵育HK-2細胞24 h建立RIF體外細胞模型[7],記為高糖(HG)組。
1.2.2 實時定量PCR(RT-qPCR)檢測GAS5和miR-136表達水平:首先以TRIzol試劑提取HK-2細胞中總RNA,利用PrimeScript逆轉錄Master Mix合成互補DNA,再用Power SYBR Green PCR Master Mix進行RT-qPCR檢測。2-ΔΔCt法分析GAS5和miR-136表達水平。GAS5上游引物5’-CCTGTGAGGTATGGTGCTGG-3’,下游引物5’-CTGTGTGCC AATGGCTTGAG-3’;內參GAPDH上游引物5’-TGTGGGCATCAATGGATTTGG-3’,下游引物5’-ACACCATGTATTCCGGGTCAAT-3’;miR-136上游引物5’-ACUCCAUUUGUUUUGAU GAUGGA-3’,下游引物5’-UCCAUCAUCAAAACAAAUGGAGU-3’;內參U6上游引物5’-AAAGCAAATCATCGGACGACC-3’,下游引物5’-GTACAACACATTGTTTCCTCG GA-3’。
1.2.3 細胞轉染和實驗分組:將對數期HK-2細胞鋪96孔板,利用脂質體試劑分別將Si-NC、Si-GAS5、miR-NC、miR-136 mimics、Si-GAS5+anti-miR-NC、Si-GAS5+anti-miR-136轉染50%融合的細胞,收獲轉染48 h細胞檢測轉染效果。用含5.5 mmol/L葡萄糖的培養液孵育HK-2細胞24 h記為正常對照(NC)組;HG組參照建模方法進行細胞處理;用含30 mmol/L葡萄糖的培養液孵育分別轉染Si-NC、Si-GAS5、miR-NC、miR-136 mimics、Si-GAS5+anti-miR-NC、Si-GAS5+anti-miR-136的HK-2細胞24 h,依次記為Si-NC組、Si-GAS5組、miR-NC組、miR-136組、Si-GAS5+anti-miR-NC組、Si-GAS5+anti-miR-136組。
1.2.4 ELISA實驗檢測IL-6和TNF-α表達:胰酶消化各組細胞,3 000 r/min離心10 min收集細胞;用冷磷酸鹽緩沖液(PBS)洗滌細胞3次,用一定功率的超聲波處理細胞懸液;再將帶破碎細胞在-20℃下冰凍,室溫溶解,反復3次,使細胞溶脹破碎。將標本于4℃離心機1 500 r/min離心10 min,收集上清按照試劑盒說明書檢測IL-6和TNF-α表達量。
1.2.5 免疫印跡法檢測α-SMA和FN1蛋白表達:RIPA裂解法處理HK-2細胞獲得總蛋白,蛋白定量后利用SDS-PAGE電泳分離將蛋白樣品分離開。隨后用濕轉移法將凝膠上的蛋白條帶轉移到硝酸纖維素膜上。室溫下用5%脫脂牛奶孵育2 h后加入一抗溶液(α-SMA為1∶300,FN1為1∶500)4℃孵育膜過夜。然后加入山羊抗兔IgG二抗(1∶1 000)室溫下孵育膜2 h。最后加入顯影劑顯色后,Image J軟件分析灰度值。目的蛋白表達量為目的條帶灰度值和內參GAPDH灰度值比值。
1.2.6 雙熒光素酶報告實驗:收集HK-2細胞接種于24孔板,當細胞密度達到50%時進行轉染。用Lipofectamine 2000試劑將miR-NC、miR-136 mimics分別與WT-GAS5或MUT-GAS5共轉染。孵育48 h后收獲轉染細胞,磷酸鹽緩沖液洗滌后,然后使用雙熒光素酶報告基因檢測系統測定熒光素酶活性。同時將pcDNA、pcDNA-GAS5、Si-NC、Si-GAS5分別轉染HK-2細胞,按照上述RT-qPCR步驟檢測轉染48 h細胞中miR-136表達量。

2 結果
2.1 GAS5和miR-136在高糖誘導的HK-2細胞中的表達 與NC組比較,HG組HK-2細胞中GAS5表達量顯著升高,差異有統計學意義(P<0.05),而miR-204-5p表達量顯著降低,差異有統計學意義(P<0.05)。見表1。
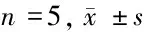
表1 GAS5和miR-136在高糖誘導的HK-2細胞中的表達
2.2 沉默GAS5對高糖誘導的HK-2細胞炎性與纖維化因子表達的影響 與NC組比較,HG組HK-2細胞GAS5表達量顯著升高(P<0.05),IL-6和TNF-α表達顯著升高(P<0.05),α-SMA和FN1蛋白表達量顯著升高(P<0.05);與HG+Si-NC組比較,HG+Si-GAS5組HK-2細胞GAS5表達量顯著降低(P<0.05),IL-6和TNF-α表達顯著降低(P<0.05),α-SMA和FN1蛋白表達量顯著降低,差異有統計學意義(P<0.05)。見表2。
表2 沉默GAS5對高糖誘導的HK-2細胞炎性與纖維化因子表達的影響
2.3 GAS5靶向調控miR-136的表達 Starbase在線分析顯示,GAS5序列中含有miR-136特異性結合位點。雙熒光素酶報告實驗顯示,與轉染miR-NC比較,轉染miR-136顯著降低HK-2細胞WT-GAS5的相對熒光素酶活性,差異有統計學意義(P<0.05),而對MUT-GAS5的相對熒光素酶活性無顯著影響。pcDNA-GAS5組HK-2細胞miR-136表達量顯著低于pcDNA組(P<0.05);Si-GAS5組HK-2細胞miR-136表達量顯著高于Si-NC,差異有統計學意義(P<0.05)。見圖1,表3、4。
圖1 GAS5中含有miR-136結合位點
表3 雙熒光素酶報告實驗
表4 lncRNA GAS5調控miR-136表達
2.4 過表達miR-136對高糖誘導的HK-2細胞炎性與纖維化因子表達的影響 與NC組比較,HG組HK-2細胞miR-136表達量顯著降低(P<0.05),IL-6和TNF-α表達顯著升高(P<0.05),α-SMA和FN1蛋白表達量顯著升高(P<0.05);與HG+miR-NC組比較,HG+miR-136組HK-2細胞miR-136表達量顯著升高(P<0.05),IL-6和TNF-α表達顯著降低(P<0.05),α-SMA和FN1蛋白表達量顯著降低(P<0.05)。見表5,圖2。

圖2 過表達miR-136對高糖誘導的纖維化因子的影響
表5 過表達miR-136對高糖誘導的HK-2細胞炎性與纖維化因子表達的影響
2.5 干擾miR-136表達逆轉沉默GAS5對高糖誘導的HK-2細胞炎性與纖維化因子表達的影響 與HG+Si-GAS5+anti-miR-NC組比較,HG+Si-GAS5+anti-miR-136組HK-2細胞miR-136表達量顯著降低,IL-6和TNF-α表達顯著升高(P<0.05),α-SMA和FN1蛋白表達量顯著升高(P<0.05)。見圖3,表6。
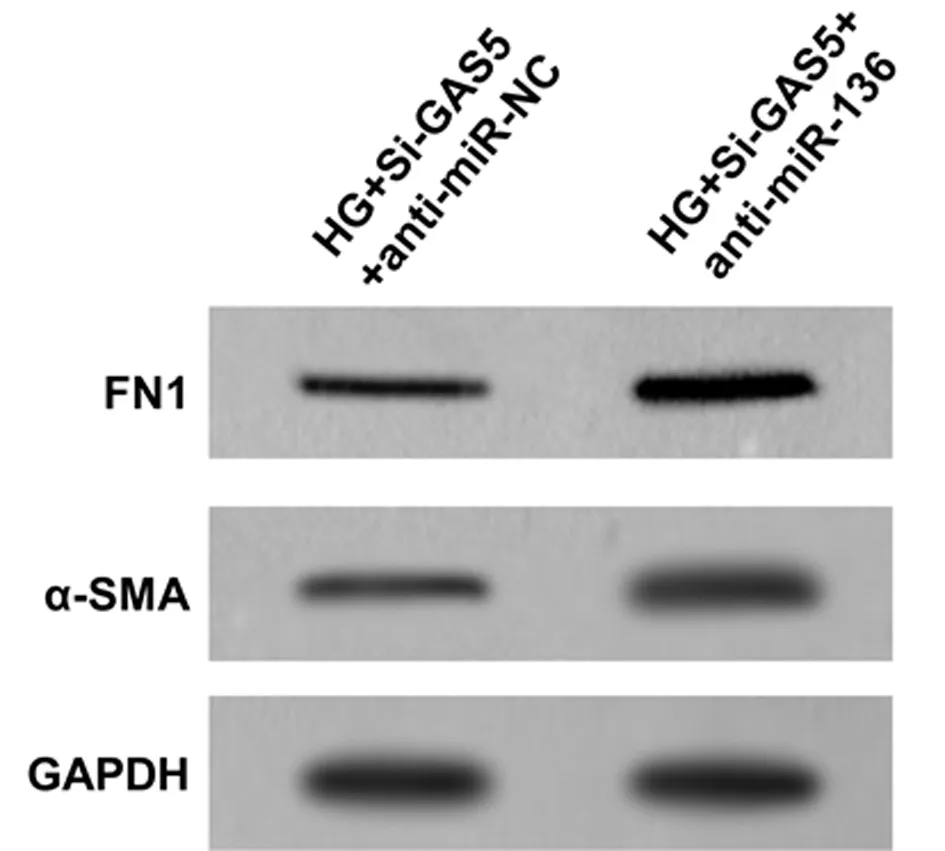
圖3 干擾miR136表達對高糖誘導的HK-2細胞纖維化因子表達的影響
表6 干擾miR136表達對高糖誘導的HK-2細胞炎性與纖維化因子表達的影響
3 討論
GAS5最初從小鼠NIH3T3細胞分離得到,其在細胞增殖、生長阻滯、凋亡和自噬等多個生物學過程中發揮重要作用。近年研究證實GAS5表達失調與多種纖維化進程有關[8]。心肌纖維化組織和活化的心肌成纖維細胞中GAS5表達降低,恢復GAS5表達水平顯著抑制心肌成纖維細胞增殖,降低MMP-2、a-SMA和Ⅰ型膠原蛋白表達,從而減輕心臟纖維化,改善心臟功能[8,9]。GAS5通過下調miR-23a表達調控信號通路抑制四氯化碳誘導的肝纖維化[10]。本研究發現高糖誘導后HK-2細胞中肌成纖維細胞標記物α-SMA、纖維化標志蛋白FN1表達水平顯著增加,GAS5表達顯著增加,提示GAS5表達改變可能與高糖誘導的RIF相關。轉染Si-GAS5分析GAS5功能發現,沉默GAS5顯著降低α-SMA和FN1蛋白表達水平,提示沉默GAS5可逆轉高糖誘導的RIF。
TGF-β1是一種極強的促纖維化因子,Wang等[11]指出在TGF-β1誘導的HK-2細胞、鏈脲佐菌素誘導的DN小鼠腎組織中GAS5表達升高,沉默GAS5表達抑制轉化生長因β1誘導的HK-2細胞纖維化,這與本研究發現的GAS5促纖維化作用一致。在DN中受損的腎小管上皮細胞、浸潤的淋巴細胞可產生炎性因子加劇腎臟炎性反應,介導腎臟纖維化[12]。本研究發現沉默GAS5表達顯著降低高糖誘導的HK-2細胞IL-6和TNF-α表達水平,說明沉默GAS5可抑制高糖誘導的HK-2細胞炎性反應。
lncRNA通過與miRNA相互作用影響miRNA活性進而調控多種生物學過程[13]。為探討GAS5在高糖誘導的HK-2細胞炎癥和纖維化中的可能機制,本研究通過Starbase數據庫在線分析發現miR-136與GAS5序列存在結合位點,并通過雙熒光素酶報告實驗、RT-qPCR進一步證實GAS5對miR-136的靶向負調控作用。目前對miR-136研究多集中在腫瘤方面,研究顯示口腔鱗狀細胞癌、胃癌、黑色素瘤、乳腺癌中miR-136表達降低,miR-136低表達促進癌細胞EMT進程[14-17]。
本研究發現高糖刺激顯著降低HK-2細胞中miR-136表達水平,提示miR-136低表達可能與高糖誘導的RIF和炎性反應相關。轉染miR-136 mimics進行功能驗證顯示,恢復miR-136表達顯著削弱高糖刺激對HK-2細胞α-SMA、FN1、IL-6和TNF-α表達的影響,說明miR-136可抑制高糖誘導的HK-2纖維化因子表達和炎性反應,這與Liu等[18]報道的miR-136的抗纖維化作用基本吻合。深入研究發現,抑制miR-136表達還可逆轉沉默GAS5對高糖誘導的HK-2細胞炎性與纖維化因子表達的影響,這進一步說明GAS5靶向miR-136在高糖誘導HK-2細胞炎性與纖維化因子表達中的的作用。
綜上所述,本研究表明沉默GAS5可抑制高糖誘導的人腎小管上皮細胞炎性和纖維化因子表達,其機制與上調miR-136表達有關。因此,GAS5/miR-136途徑是改善改善RIF、延緩DN進展的潛在靶點。
