犏牛PPP1R11基因的克隆及在睪丸中的表達規律研究
2022-03-08 06:59:02閔星宇楊麗雪于海玲胡宇磊楊滿珍楊璐瑜熊顯榮
畜牧獸醫學報 2022年2期
閔星宇,楊麗雪,于海玲,胡宇磊,楊滿珍,楊璐瑜,李 鍵,2,熊顯榮,
(1.西南民族大學畜牧獸醫學院,成都 610041; 2.青藏高原動物遺傳育種資源保護與利用國家教育部重點實驗室,成都 610041; 3.動物科學國家民委重點實驗室,成都 610041)
犏牛為母牦牛(Bosgrunniens)與平原牛(Bostaurus)雜交得到的F1代,主要分布于我國青藏高原地區的優良家畜資源。牦牛生產性能低下,已無法滿足青藏高原地區日益增長的奶肉需求,而平原牛又無法適應高原惡劣的高寒低氧環境。犏牛具有顯著的雜種優勢,產奶性能和產肉性能均優于牦牛,并有良好的高原適應性[1-3]。然而,雄性犏牛精子發生阻滯,使得無法通過橫交固定其雜種優勢,這嚴重制約了雜種優勢的利用[4-5]。因此,開展犏牛繁殖生理的研究對提高青藏高原畜牧生產水平、改善牧民生活品質和發展青藏高原特色畜牧產業都具有重要的價值。
絲氨酸/蘇氨酸蛋白磷酸酶1(serine/threonine protein phosphatase,PP1)是一種去磷酸化酶,能夠通過翻譯后調控參與多種生物學過程[6-7]。在哺乳動物中共發現4種亞型,包括PP1α、PP1β、PP1γ1和PP1γ2,其中PP1γ2蛋白由蛋白磷酸酶1催化亞基γ(protein phosphatase 1 catalytic subunit gamma, PPP1CC)基因編碼,主要富集于睪丸組織并參與精子細胞發生和成熟[8-9],敲除PPP1CC基因造成雄性不育[10-11]。蛋白磷酸酶1調節亞基11(protein phosphatase 1 regulatory subunit 11,PPP1R11)最早通過酵母雙雜交試驗鑒定到是PP1的結合蛋白,又被稱作INH3、TCTEX5、HCGV、IPP3,能夠抑制PP1γ2的去磷酸化作用[12-13]。前期研究表明,PPP1R11基因是t-complex的重要組成部分,而t-complex 與精子活力受損和雄性不育有關[14]。Northern印跡雜交和免疫組織熒光染色顯示,PPP1R11 mRNA和蛋白主要分布在小鼠睪丸組織中,其表達水平顯著高于其他組織,在雄性不育小鼠中檢測到PPP1R11基因的下調,推測該基因的下調導致生殖細胞的凋亡[13,15]。可見,PPP1R11基因在調控雄性配子發生和精子成熟中有重要作用。
目前,關于PPP1R11基因的研究主要集中在小鼠上,而在雄性不育犏牛上的研究還尚未報道。本研究以犏牛為研究對象,利用RT-PCR技術克隆獲得犏牛PPP1R11編碼區完整序列,并對該序列進行生物信息學分析,同時采用qRT-PCR和IHC染色檢測該基因和蛋白在犏牛各組織及各發育階段睪丸組織中的表達,以期為研究PPP1R11調控精子發生機制提供參考,為進一步從分子水平解析雄性犏牛不育機制奠定基礎并提供理論依據。
1 材料與方法
1.1 試驗動物與樣品采集
將胎牛時期(5~6月齡)、幼年時期(1~2歲)、成年時期(3~4歲)犏牛作為本研究試驗對象,樣品采自成都市周邊屠宰場。健康的犏牛屠宰后迅速采集睪丸、附睪、心、肝、脾、肺、腎、大腸、小腸、胃、肌肉和脂肪。以上所有樣品每組各取3頭為生物學重復,樣品使用高壓滅菌處理的生理鹽水沖洗剪成0.5 cm×0.5 cm×0.5 cm的組織塊,一部分放入凍存管,置于液氮罐中帶回實驗室,于-80 ℃保存;另一部分使用4%多聚甲醛固定,用于IHC染色檢測。
1.2 犏牛和牦牛組織總RNA的提取與反轉錄
冷凍的犏牛和牦牛各組織使用高通量組織研磨器研磨后,按照Trizol法,經離心、沉淀、清洗和溶解等步驟進行總RNA的提取,使用紫外分光光度計(Biospec-nano,日本)檢測總RNA的濃度和純度,選取OD260 nm/OD280 nm介于1.8~2.0的RNA,并經1%瓊脂糖凝膠電泳檢測其完整性,置于-80 ℃保存備用。根據Revert Aid First Strand cDNA Synthesis Kit反轉錄試劑盒(Thermo scientific,美國)說明書合成第一鏈cDNA,置于-20 ℃保存。
1.3 引物的設計與基因克隆
從NCBI數據庫中選取野牦牛(Bosmutus)PPP1R11基因預測序列(登錄號:XM_014483599.1)和β-actin基因序列(登錄號:DQ838049.1),使用Primer 5.0軟件設計引物(表1),并送至南京金斯瑞公司進行合成。以犏牛睪丸組織cDNA為模板,利用RT-PCR技術在PCR儀(Applied Biosystems,美國)擴增犏牛PPP1R11基因CDS區序列,PCR擴增體系為25 μL:12.5 μL 2×Phanta Max Master Mix(Dye Plus)(南京諾唯贊生物科技股份有限公司),1 μL cDNA模板,1 μL上、下游引物,ddH2O補足至25 μL。PCR擴增程序:95 ℃預變性3 min;95 ℃變性15 s,60 ℃退火15 s, 72 ℃延伸45 s,35個循環;72 ℃徹底延伸5 min,4 ℃保存。使用濃度為1.5%瓊脂糖凝膠電泳分離后,在紫外光下切割回收目的產物進行純化,交由上海生工生物工程股份有限公司(成都)測序。
1.4 PPP1R11基因的生物信息學分析
利用ORF Finder查找犏牛PPP1R11克隆序列開放閱讀框并翻譯成氨基酸序列。采用T-COFFEE在線軟件并結合DNAMAN 9.0軟件對不同物種PPP1R11的氨基酸序列同源性進行比較,通過MEME在線工具分析查找具有高度保守性的基序,并用MEGA 7.0軟件構建系統發生樹。利用ExPASy ProtParam、TMHMM、PSORTⅡ和SignalP在線軟件進行犏牛PPP1R11蛋白基本理化性質、跨膜區、亞細胞定位和信號肽分析。糖基化位點和磷酸化位點預測使用NetPhos 3.1、NetOGlyc 4.0和NetNGlyc 1.0軟件在線分析。該蛋白二級結構、三級結構和互作蛋白通過SOPMA、VMD 1.9.3和STRING 11.0預測。
1.5 犏牛PPP1R11基因的表達分析
以β-actin為內參基因,使用Bio-Rad實時熒光定量儀(美國)檢測PPP1R11 mRNA在犏牛各組織(睪丸、附睪、心、肝、脾、肺、腎、大腸、小腸、胃、肌肉、脂肪)中的表達量并進行表達譜分析,及其在不同年齡組犏牛睪丸組織中的表達量。qRT-PCR反應體系如下:2×NovaStart SYBR qPCR Super Mix plus 10 μL,cDNA 1 μL,上、下游引物各0.5 μL,補充ddH2O至20 μL。PCR擴增條件如下:95 ℃預變性3 min;95 ℃變性10 s,60 ℃退火30 s,40個循環;熔解曲線為65~95 ℃每10 s增加0.5 ℃。
1.6 免疫組織化學染色
將不同時期睪丸組織從固定液中取出,制作石蠟切片。切片經二苯甲中脫蠟,酒精洗滌,EDTA抗原修復,PBS洗滌后,放入3%雙氧水阻斷內源性過氧化物酶;5%胎牛血清室溫封閉30 min,滴加一抗(多克隆兔抗PPP1R11,1∶200稀釋,博奧森)后4 ℃孵育過夜,陰性對照不加一抗;滴加二抗(多聚化山羊抗兔IgG,辣根過氧化物酶標記),室溫孵育50 min;滴加DAB顯色液顯色,自來水沖洗切片終止顯色;蘇木精復染3 min左右,復染細胞核;中性樹膠封片。使用激光共聚焦顯微鏡(Zeiss,德國)觀察并拍照。
1.7 數據統計分析
應用2-ΔΔCt法分析qRT-PCR檢測結果,結果使用“平均值±標準誤(Mean ± SEM)”表示。使用SPSS 19.0軟件的T-test檢驗進行數據分析,P<0.05代表差異顯著,P<0.01代表差異極顯著。
2 結 果
2.1 犏牛PPP1R11基因克隆及生物信息學分析
2.1.1 犏牛PPP1R11克隆 以犏牛睪丸組織cDNA為模板,F1、R1為上、下游引物,PCR擴增犏牛PPP1R11基因編碼區序列,1.5%瓊脂糖凝膠電泳檢測顯示,目標條帶與預期結果相符(圖1A)。經測序得到893 bp的核苷酸序列,其中犏牛PPP1R11基因CDS為324 bp,共編碼107個氨基酸(圖1B)。
2.1.2 犏牛PPP1R11同源性比較與系統進化樹 對不同哺乳動物PPP1R11的氨基酸序列同源性分析發現,PPP1R11在不同哺乳動物中高度保守,其中犏牛與牦牛和黃牛同源性最高(100%),其次為瘤牛(99.07%)、水牛(99.07%)、綿羊(99.07%)、山羊(99.07%)、羊駝(98.06%)、豬(97.09%)、馬(97.09%)、大狐蝠(96.12%)、藍鯨(95.28%)、港海豹(95.24%)、智人(95.15%),與鴨嘴獸的同源性最低(84.88%)(圖2A)。通過MEME軟件在線分析獲得PPP1R11蛋白最保守的2個基序(圖2B),分別位于犏牛PPP1R11氨基酸序列N端第6~55和58~107位點。基于15種哺乳動物PPP1R11不同氨基酸序列,采用鄰接算法構建系統進化樹(圖3),發現犏牛和黃牛聚為一支,在進化上與牦牛、瘤牛、水牛、綿羊和山羊親緣關系較近,與非反芻類動物親緣關系較遠,這與同源性比較結果一致。
2.1.3 犏牛PPP1R11理化性質 犏牛PPP1R11蛋白的分子量、理論等電點和分子式分別為12.014 ku、8.33和C503H803N163O164S8;氨基酸組成中,脯氨酸(Pro)占比最高,其含量為13.1%;帶負電荷(Asp + Glu)和帶正電荷(Arg + Lys)的氨基酸殘基總數分別16和18個,表明該蛋白可能帶正電。糖基化位點預測表明,犏牛PPP1R11蛋白含1個 N糖基化位點和18個O糖基化位點;磷酸化位點預測顯示,該蛋白有19個磷酸化位點,其中絲氨酸(Ser)磷酸化位點11個,蘇氨酸(Thr)磷酸化位點7個,酪氨酸(Tyr)磷酸化位點1個。該蛋白亞細胞定位主要存在于細胞核(47.8%),其次為線粒體(34.8%)和細胞質(17.4%);不含信號肽且無跨膜結構域。
2.1.4 犏牛PPP1R11結構與互作蛋白預測 采用SOPMA預測PPP1R11蛋白的二級結構,其中18個氨基酸(16.82%)形成α螺旋,16個氨基酸(14.95%)形成延伸鏈,9個氨基酸(8.41%)形成β轉角,64個氨基酸(59.81%)形成無規則卷曲(圖4A);通過VMD 1.9.3預測到犏牛PPP1R11蛋白三級結構(圖4B)。采用STRING數據庫分析犏牛PPP1R11潛在相互作用蛋白顯示,PPP1R11可能與PPP1R2、PPP1R7、PPP1CA、PPP1CB、PPP1CC、PPA2、CIRH1A和UTP20等10個蛋白存在相互作用(圖4C)。PPP1R11蛋白處于互作網絡核心位置,且與PPP1R7、PPP1CB、PPP1R2、UTP20蛋白間的相互作用具有更高可信度。
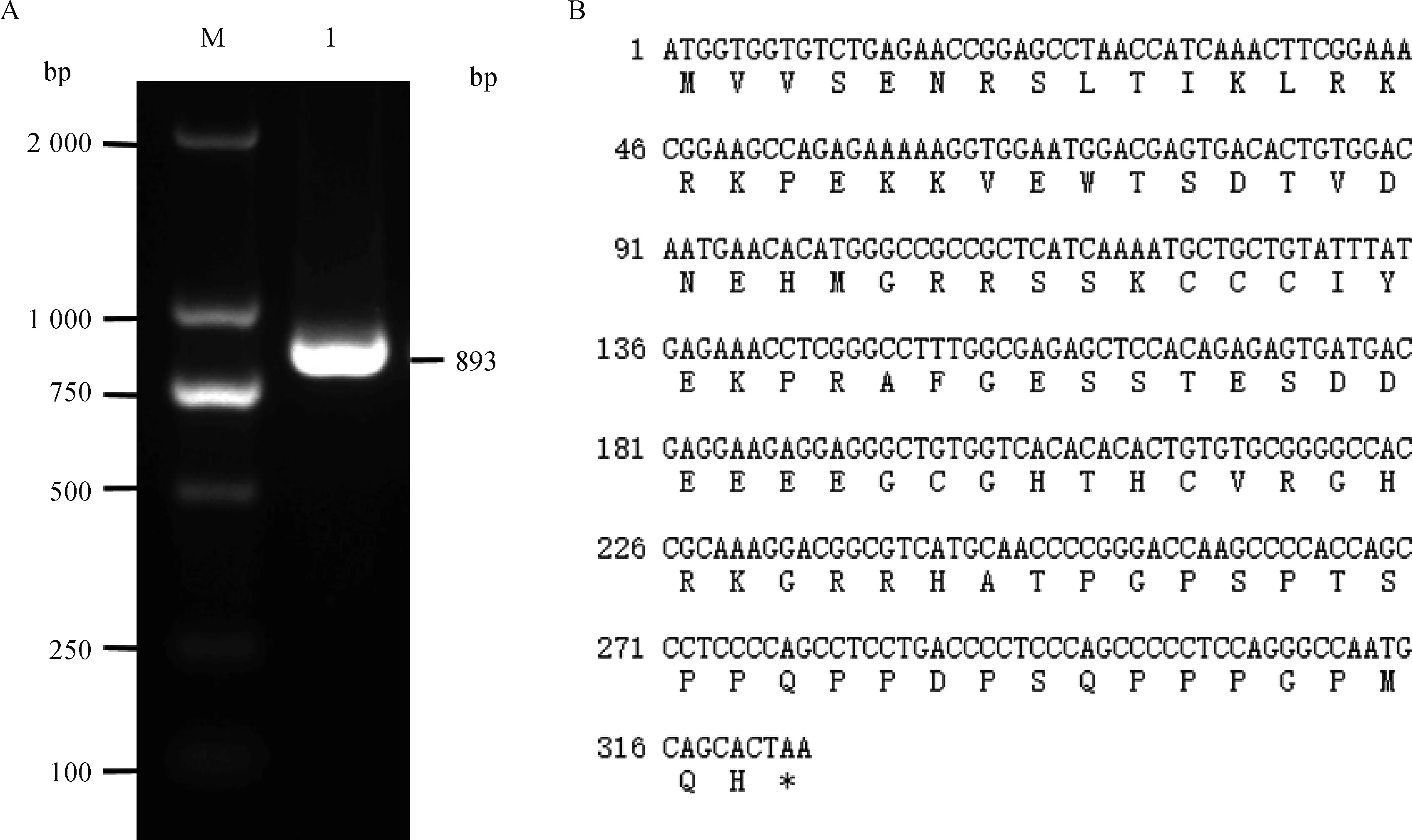
A: M. DNA相對分子質量標準DL2000; 1. 犏牛PPP1R11基因擴增產物;B.犏牛PPP1R11基因核苷酸及其推測的氨基酸序列A: M. DL2000 marker; 1.PCR product of cattle-yak PPP1R11 gene;B. Nucleotide sequence and the deduced amino acid sequence of cattle-yak PPP1R11圖1 犏牛PPP1R11基因的PCR擴增結果Fig.1 PCR amplification results of cattle-yak PPP1R11 gene
2.2 犏牛PPP1R11基因的表達模式
2.2.1 犏牛PPP1R11組織表達譜 以β-actin作為內參基因,利用qRT-PCR技術檢測PPP1R11 mRNA在犏牛睪丸、附睪、心、肝、脾、肺、腎、大腸、小腸、胃、肌肉和脂肪組織中的表達量(圖5)。結果顯示,該基因在犏牛12個組織中均有表達,在睪丸、脾和胃組織中表達量較高,其在睪丸中的表達水平極顯著高于其它各個組織(P<0.01)。
2.2.2 犏牛睪丸中PPP1R11的表達規律 采用qRT-PCR檢測PPP1R11 mRNA在犏牛不同發育時期睪丸組織中的相對表達水平(圖6)。結果顯示,該基因在犏牛睪丸組織中的表達隨年齡的增加呈上升趨勢;5~6月齡胎牛睪丸組織中幾乎不表達,表達量低于同齡的牦牛,但差異不顯著(P>0.05);1~2歲和3~4歲犏牛睪丸中的表達水平極顯著低于同時期牦牛(P<0.01)。
2.2.3 犏牛PPP1R11的細胞定位 基于犏牛和牦牛PPP1R11 mRNA在睪丸中的表達差異極顯著,本試驗進一步通過免疫組織化學染色方法檢測PPP1R11蛋白在犏牛和牦牛不同年齡段睪丸組織中的細胞定位和表達情況(圖7)。結果顯示,犏牛和牦牛5~6月齡胎牛睪丸組織生精小管形態無明顯差異;1~2歲和3~4歲犏牛與牦牛睪丸組織中,犏牛生精小管管徑縮小,空腔增大,生精上皮皺縮,細胞組成差異顯著,僅有精原細胞、支持細胞和少量初級精母細胞(PS)松散分布,無次級精母細胞(SS)和精子細胞。IHC染色發現,PPP1R11蛋白在犏牛和牦牛各年齡段睪丸組織中均有表達。其中5~6月齡 犏胎牛睪丸組織中,PPP1R11蛋白主要定位于精原干細胞(SSC),支持細胞(SC)幾乎不表達,與同時期牦牛具有相同的蛋白定位;1~2歲和3~4歲 犏牛睪丸組織中,PPP1R11蛋白主要定位于初級精母細胞和間質細胞(IC),精原細胞和支持細胞中蛋白陽性信號較弱,而牦牛各級生殖細胞、支持細胞和間質細胞中均檢測到PPP1R11陽性信號。

A. 15種不同哺乳動物PPP1R11氨基酸序列的多重比較;B. PPP1R11氨基酸序列中2個最保守基序A. Multiple alignment of PPP1R11 amino acid sequences from 15 different mammalian species; B. Two of most conserved motifs in PPP1R11 amino acid sequence圖2 不同哺乳動物PPP1R11氨基酸序列對比Fig.2 Alignment of amino acid sequence of PPP1R11 among different mammals
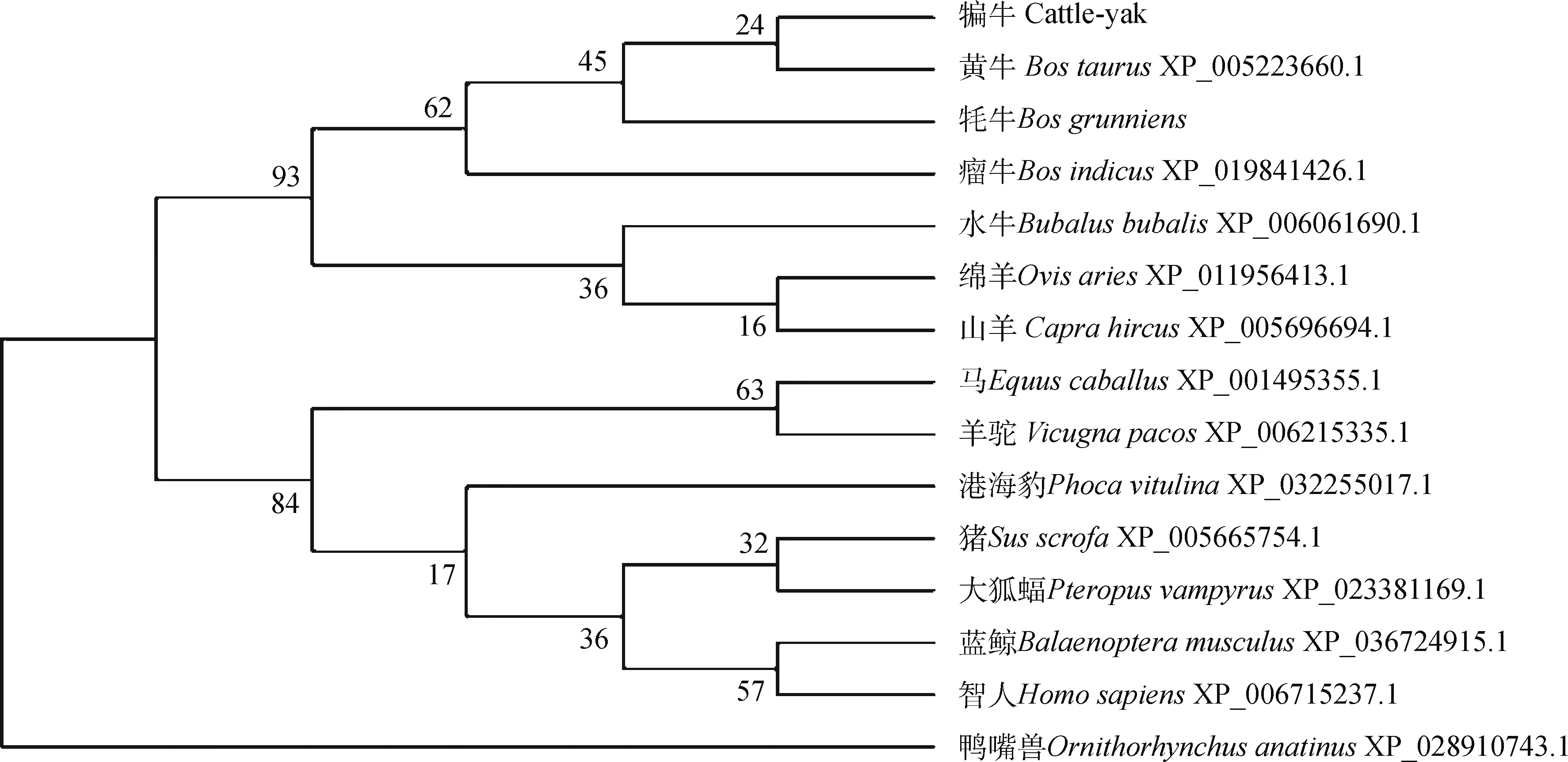
圖3 PPP1R11蛋白氨基酸序列系統進化樹Fig.3 Phylogenetic tree for PPP1R11 amino acid sequences
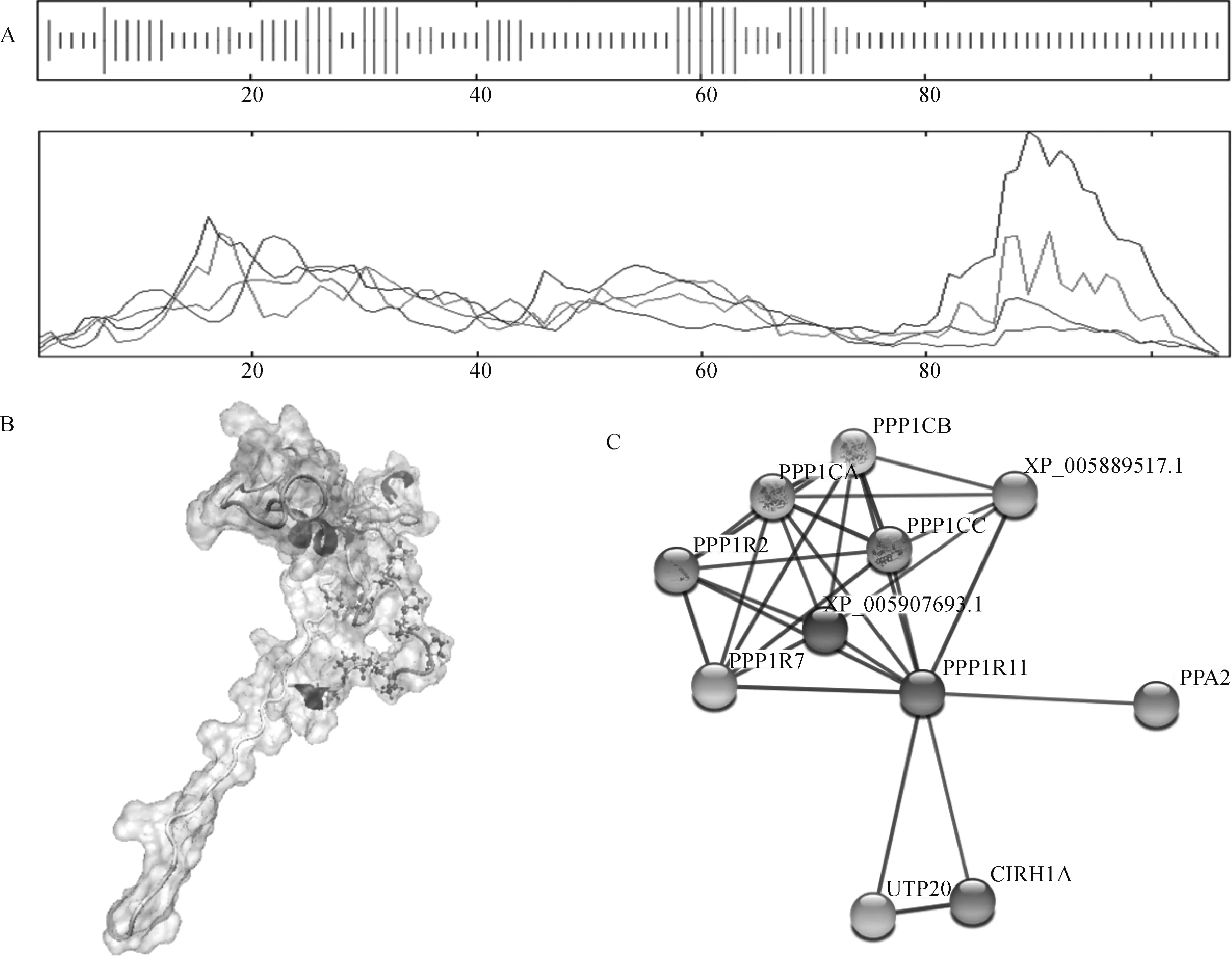
A. 犏牛PPP1R11蛋白二級結構;B. 犏牛PPP1R11蛋白三級結構; C. 犏牛PPP1R11的蛋白互作網絡A. Secondary structure of cattle-yak PPP1R11 protein; B. Tertiary structure of cattle-yak PPP1R11 protein; C. Interaction network for cattle-yak PPP1R11 protein圖4 犏牛PPP1R11蛋白的空間結構及互作蛋白Fig.4 Spatial structure and interaction proteins of cattle-yak PPP1R11
3 討 論
雄性生殖力是種畜的重要評價指標,性狀優良的種公畜往往能夠產生更大的經濟效益,故而提高種公畜生殖力對養殖業的發展具有促進作用。犏牛是牦牛和黃牛的雜交后代,具有雜種優勢顯著和雄性不育的特點[16],其雄性不育犏牛可作為研究雄性生殖調控機理的天然理想模型。據報道,蛋白質磷酸化與去磷酸化參與了30%~70%的細胞信號轉導過程[6],精子發生是一個高度動態變化的復雜過程,需要多種磷酸化酶的參與[17]。其中,去磷酸化酶PP1γ2能夠直接調控哺乳動物的精子發生,而PPP1R11能夠通過抑制PP1γ2的表達來維持精子的正常發生和成熟[15,18]。因此,探索PPP1R11在睪丸中的表達規律對于研究生殖調控和解析犏牛雄性不育機制具有重要的價值。
本研究成功克隆了犏牛PPP1R11基因CDS區全長序列,由324 bp的核苷酸組成,編碼107個氨基酸,這與NCBI中提供的野牦牛預測序列(XM_014483599.1)和已報道黃牛[19]的序列完全一致,該基因CDS區長度與瘤牛、水牛、山羊和綿羊一致。通過氨基酸序列對比發現,犏牛PPP1R11蛋白與其它14種哺乳動物同源性在95%以上(鴨嘴獸除外),并識別到2個最為保守的基序。這些結果表明,犏牛PPP1R11與其它哺乳動物相比具有高度同源性和進化上的高度保守性,PPP1R11可能在不同物種中發揮較為一致的生物學功能。鑒于PPP1R11主要通過與相應蛋白互作發揮功能,本研究進一步分析了犏牛PPP1R11的互作蛋白發現,該蛋白與PPP1R2、PPP1CB等10個蛋白質分子存在相互作用。其中PPP1CA、PPP1CB、PPP1CC和PP2A互作為不同類型蛋白磷酸酶,被廣泛報道能夠參與調控哺乳動物睪丸發育、精子發生和成熟[20-23]。有研究表明,在小鼠原始生殖細胞中特異性敲除PP2A可導致雄性小鼠不育[24],通過轉錄組測序技術檢測到PP2A在雄性不育犏牛睪丸中也表現為下調[25],此外PP2A還可以通過調控減數分裂來調節卵母細胞的成熟[26]。PPP1R2、PPP1R7和PPP1R11均為PP1的調節劑,能夠在小鼠附睪中共同調控精子的成熟[15],PPP1R11能夠通過抑制PPP1R7與PP1結合從而確保有絲分裂過程中動粒與紡錘體的結合[27],而PPP1R11與PPP1R2能否直接互作還未見報道。有研究表明,UTP20能激活RNA聚合酶Ⅰ轉錄并促進細胞增殖[28],推測PPP1R11可能與UTP20互作從而參與調控細胞增殖。綜上,這些潛在與犏牛PPP1R11互作的蛋白分子均在睪丸發育和精子發生中發揮了重要的作用,這為深入理解PPP1R11在哺乳動物睪丸發育及精子發生中的分子機制提供了思路和見解。
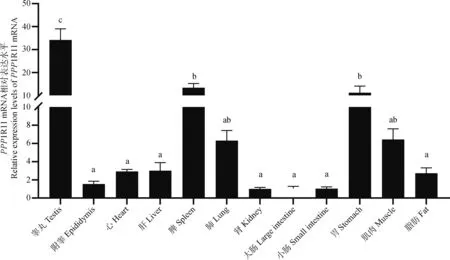
不同的字母表示差異極顯著(P<0.01)。下同Different letters represent the significant differences(P<0.01). The same as below圖5 PPP1R11 mRNA在犏牛中的組織表達譜Fig.5 Tissue expression profile of PPP1R11 mRNA in cattle-yak
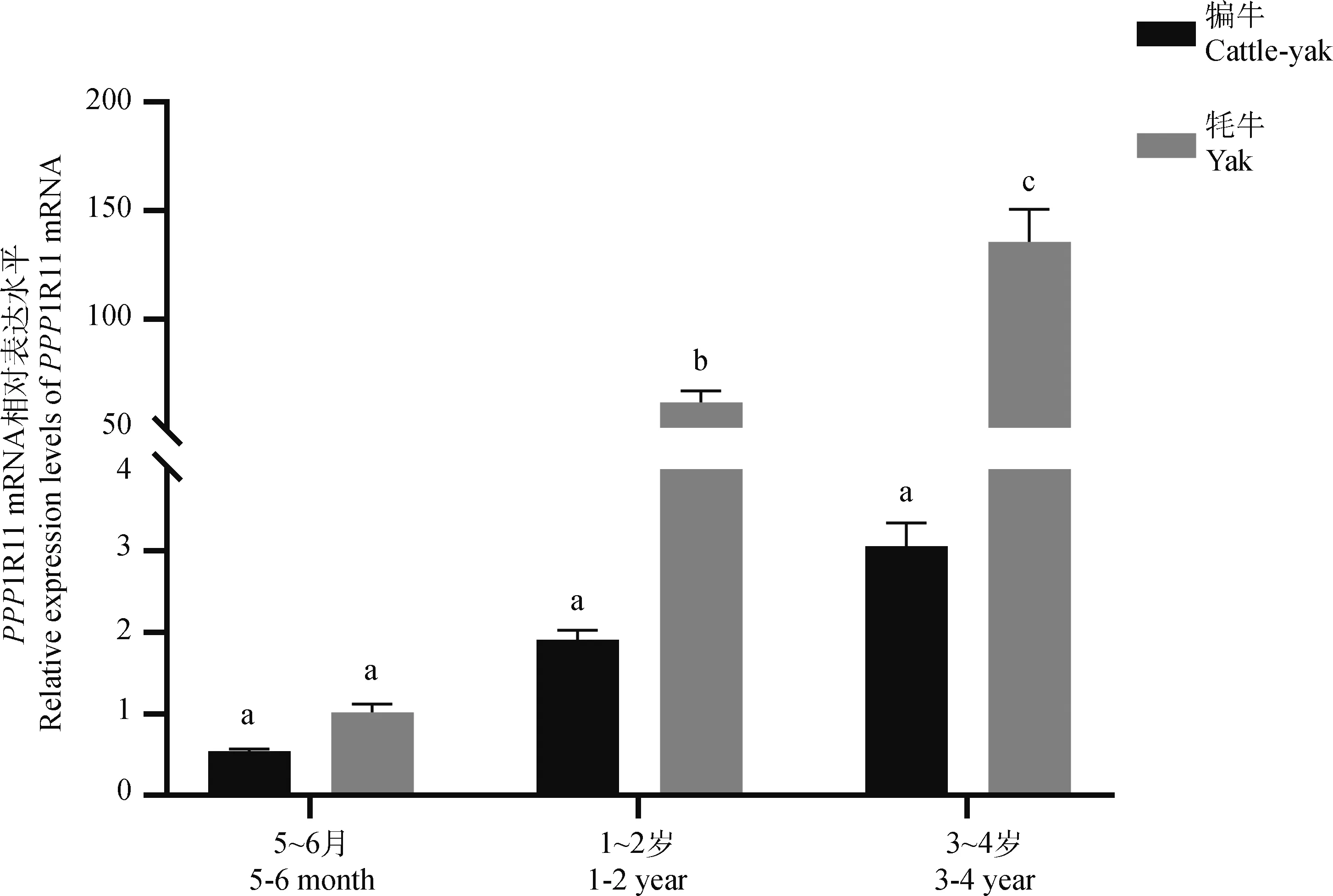
圖6 PPP1R11 mRNA在不同發育階段犏牛睪丸中的表達規律Fig.6 Expression patterns of PPP1R11 mRNA in testes at different developmental stages of cattle-yak
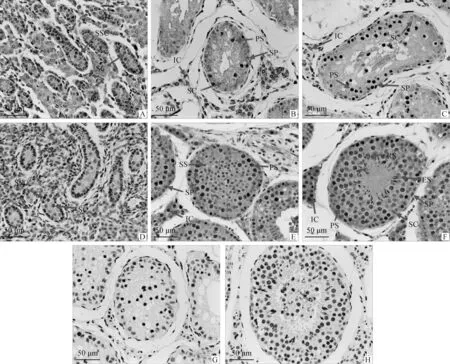
A. 犏胎牛睪丸(5~6月齡);B. 幼年犏牛睪丸(1~2歲);C. 成年犏牛睪丸(3~4歲);D. 牦胎牛睪丸(5~6月齡);E. 幼年牦牛睪丸(1~2歲);F. 成年牦牛睪丸(3~4歲);G. 成年犏牛睪丸(陰性對照);H. 成年牦牛睪丸(陰性對照)。SP. 精原細胞;PS. 初級精母細胞;SS. 次級精母細胞;SC. 支持細胞;IC. 間質細胞;RS. 圓形精子細胞;ES. 長形精子細胞A. Fetal cattle-yak testis (5-6 months); B. Juvenile cattle-yak testis (1-2 years old); C. Adult cattle-yak testis (3-4 years old); D. Fetal yak testis (5-6 months); E. Juvenile yak testis (1-2 years old); F. Adult yak testis (3-4 years old); G. Adult cattle-yak testis (Negative control); H. Adult yak testis (Negative control). SP. Spermatogonium; PS. Primary spermatocyte; SS. Secondary spermatocyte; SC. Sertoli cell; IC. Leydig cell; RS. Round spermatid; ES. Elongating spermatid圖7 不同階段雄性犏牛生殖細胞發育及PPP1R11蛋白定位Fig.7 The growth of male cattle-yak germ cells and location of PPP1R11 protein at different stages
通過qRT-PCR檢測了PPP1R11基因在犏牛不同組織中的表達,組織表達譜分析顯示,該基因在犏牛各組織中均有表達,在睪丸組織中表達最高,這與小鼠[29]、大鼠[30]和綿羊[31]PPP1R11基因組織表達譜相一致,PPP1R11基因在多種哺乳動物睪丸組織中的高表達表明該基因與雄性生殖相關,且在犏牛睪丸中也可能發揮了相似的功能。Goswami等[15]研究表明,PPP1R11基因在小鼠附睪中能夠促進精子的成熟,而犏牛附睪中該基因表達水平較低,推測是因為犏牛精子發生阻滯,附睪中無精子細胞可成熟。此外,本試驗還檢測到該基因在脾和胃中有較高表達,推測該基因除參與睪丸發育、精子生成和成熟外,在其他組織中也可能發揮作用,其具體功能有待進一步研究。
本試驗檢測到PPP1R11在犏牛睪丸組織中的表達水平隨睪丸生長發育呈上升趨勢,與同時期牦牛相比均較低,且在幼年和成年時期差異極顯著。有研究表明,過表達PPP1R11能夠抑制細胞的凋亡[32],在T-單倍型突變雄性不育小鼠模型中,PPP1R11也表現為下調[33],由此推測PPP1R11在睪丸中的表達水平與雄性生殖力成正相關,而雄性犏牛不育也可能與此有關聯。為進一步探討PPP1R11基因在犏牛睪丸中發揮的作用,利用IHC染色檢測PPP1R11蛋白在犏牛3個時期睪丸中的表達和細胞定位。成年期犏牛生精小管內只觀察到精原細胞、支持細胞和少量初級精母細胞,未觀察到次級精母細胞和精子細胞,說明雄性犏牛精子發生阻滯于初級精母細胞,這與前人研究結果一致[34-36]。在幼年期和成年期犏牛睪丸組織中,PPP1R11蛋白主要定位于犏牛初級精母細胞和間質細胞,其蛋白陽性信號強度顯著低于同時期牦牛,這與qRT-PCR的結果一致。正常生殖力小鼠睪丸中,PPP1R11蛋白主要定位于圓形期精子和長形期精子,其他各級生殖細胞和支持細胞亦有表達[13-15],正常生殖力牦牛生精小管中各級生殖細胞均檢測到PPP1R11蛋白陽性信號,本試驗中犏牛精原細胞幾乎不表達PPP1R11蛋白,這與小鼠和牦牛的結果不一致,推測該蛋白在犏牛精原細胞中的下調導致初級精母細胞生成的減少,從而造成精子發生阻滯。綜上,PPP1R11基因可能參與調控哺乳動物精子發生,其mRNA和蛋白在犏牛睪丸中的差異表達可能是造成雄性犏牛減數分裂阻滯的原因之一。
4 結 論
本試驗成功克隆了犏牛PPP1R11基因序列,該基因在犏牛與牦牛睪丸組織中的mRNA表達水平和蛋白定位存在差異,提示PPP1R11可能與維持雄性正常生殖力相關。本研究結果為進一步探究雄性犏牛不育的機制和PPP1R11在雄性生殖系統中的生理功能和調控機制提供了理論依據。
