lncRNA ACTA2-AS1通過靶向下調miR-586表達抑制子宮內膜癌生物學行為的研究
2021-12-28 09:21:44劉雨欣
醫學研究雜志 2021年12期
關鍵詞:檢測
劉雨欣 秦 黎 王 琳 蔡 晶 余 莉
子宮內膜癌是對女性健康生命危害最大的惡性腫瘤之一,其細胞增殖較快、遷移性較強,因而惡性程度較高[1]。子宮內膜癌位居婦科生殖系統腫瘤第2位,術后復發率高,患者5年生存率較低[2]。針對子宮內膜癌分子機制的研究,對于子宮內膜癌的分子靶向治療具有重要意義。長鏈非編碼RNA(lncRNA)是一種具有生物學功能的非編碼RNA,長度往往超過200個核苷酸[3]。lncRNA廣泛參與細胞的發育、增殖、遷移、衰老等生命活動,其在癌癥的起始和發展環節具有重要調控功能[4, 5]。對lncRNA的深入研究可能為子宮內膜癌的篩查、診療、預后評估提供新的生物學標志物。目前已有報道顯示,ACTA2-AS1在肝癌、肺腺癌、結腸腺癌等多種癌癥組織或細胞株中低表達,發揮抑癌基因的作用[6~8]。ACTA2-AS1在子宮內膜癌組織和細胞株中的表達和作用尚未明確。本研究旨在探究ACTA2-AS1在子宮內膜癌中的表達,分析ACTA2-AS1通過靶向調控miR-586對子宮內膜癌細胞生物學行為的影響,為子宮內膜癌的診療提供新的分子靶點。
材料與方法
1.細胞與試劑:子宮內膜癌細胞HEC-1A、Ishikawa、AN3CA、HEC-1B及人正常子宮內膜上皮細胞ESC購自美國標準培養物保藏中心。qRT-PCR試劑盒購自日本TaKaRa公司。ACTA2-AS1過表達質粒和空白質粒(pGPU6)、miR-586 mimic、miR-NC mimic、熒光素酶報告載體(ACTA2-AS1-wt、ACTA2-AS1-mut)購自上海吉瑪生物制藥有限公司。Lipofectamine 2000購自美國Invitrogen公司。雙熒光素酶報告基因試劑盒購自美國Promega公司。RPMI 1640培養基、胎牛血清、DMEM/F12培養基、購自美國Amresco公司。細胞周期試劑盒購自北京鼎國昌盛生物技術有限公司。一抗PTEN、β-tubulin、p-PI3K、p-AKT、p-mTOR及山羊抗兔二抗均購自美國CST公司。
2.ACTA2-AS1在子宮內膜癌組織中的表達:采用GEPIA數據庫(http:∥gepia.cancer-pku.cn/index.html)分析ACTA2-AS1在子宮內膜癌組織和癌旁組織的表達水平。
3.細胞培養與轉染:子宮內膜癌細胞株Ishikawa、AN3CA、ESC細胞用含10%胎牛血清的DMEM/F12培養基,HEC-1A、HEC-1B細胞用含10%胎牛血清的RPMI 1640培養基,在37℃、5%CO2培養箱中孵育。取生長狀態良好的AN3CA細胞接種于12孔板,待AN3CA細胞生長融合度為30%,根據Lipofectamine 2000轉染試劑說明書轉染空白質粒(NC組)或ACTA2-AS1過表達質粒(ACTA2-AS1組)。
4.qRT-PCR檢測ACTA2-AS1、miR-586、PTEN mRNA相對表達:用TRIzol溶液提取組織和細胞樣品總RNA,進一步反轉錄為cDNA。ACTA2-AS1、PTEN mRNA以GAPDH為內參,miR-586以U6為內參,根據qRT-PCR試劑盒說明書擴增檢測,以2-ΔΔCt公式計算ACTA2-AS1、miR-586相對表達。ACTA2-AS1上游引物為5′-GTTCTGGAGGCTGGCTTGATATGG-3′,下游引物為5′-TCCTTCA-TCGGTAGGCAACAAACG-3′;U6上游引物為5′-CTCGCTTCGGCAGCACA-3′,下游引物為5′-AACGCTTCACGAATTTGCGT-3′;miR-586上游引物為5′-ACACTCCAGCTGGGTATGCATTGTATTTTTAGGT-3′,下游引物為5′-CTCAACTGGTGTCGTGGA-3′;GAPDH上游引物為5′-CTCAGACACCATGGGGAAGGTGA-3′,下游引物為5′-ATGATCTTGAGGCTGTTGTCATA-3′;PTEN上游引物為5′-AGGGACGAACTGGTGTAATGA-3′,下游引物為5′-CTGGTCCTTACTTCCCCATAGAA-3′。
5.流式細胞法檢測AN3CA細胞周期:NC組和ACTA2-AS1組AN3CA細胞用預冷的磷酸鹽緩沖液洗滌5次,用預冷的乙醇溶液重懸、固定,在4℃下孵育13h。用預冷的磷酸鹽緩沖液洗滌5次,用200μl RNA酶溶液重懸,37℃下孵育20min,加入40μl溴化丙錠溶液,4℃下孵育20min,用流式細胞儀檢測AN3CA細胞周期。
6.細胞劃痕實驗檢測AN3CA細胞遷移能力:NC組和ACTA2-AS1組AN3CA細胞鋪于12孔板上,AN3CA細胞生長融合度為90%,用10μl的無菌移液槍槍頭在12孔底劃線,保證劃痕為單層細胞。用預冷的磷酸鹽緩沖液洗滌5次,至沒有脫落的細胞存在。添加不含胎牛血清的培養基,在100倍倒置顯微鏡下,于0h和24h拍照并測量細胞遷移距離,計算細胞遷移率。
7.雙熒光素酶報告基因檢測驗證ACTA2-AS1與靶基因的結合:運用LncBase v.2數據庫預測ACTA2-AS1的靶基因為miR-586。在AN3CA細胞中,共轉染ACTA2-AS1-wt或ACTA2-AS1-mut以及miR-586、miR-NC。共轉染48h后,采用細胞裂解液收集AN3CA細胞,采用雙熒光素酶報告基因檢測試劑盒分析每組AN3CA細胞中熒光素酶的相對活性。
8.Western blot法檢測PTEN蛋白和PI3K-AKT-mTOR信號通路蛋白表達:用RIPA裂解液提取ACTA2-AS1組和NC組AN3CA細胞總蛋白,BCA法分析蛋白濃度,每個泳道60μg蛋白的上樣量行SDS-PAGE凝膠電泳,將分離蛋白電轉至聚偏氟乙烯膜。10%脫脂牛奶封閉0.5h,加入一抗PTEN(1∶1000稀釋)、β-tubulin(1∶3000稀釋)、p-PI3K(1∶1000稀釋)、p-AKT(1∶1000)、p-mTOR(1∶1000),4℃孵育13h。加入羊抗鼠二抗(1∶5000稀釋),在室溫下搖床孵育3h。按比例配制ECL顯影發光液,在暗室內曝光、采集圖像。

結 果
1.ACTA2-AS1在子宮內膜癌組織中的表達:GEPIA數據庫顯示,ACTA2-AS1在子宮內膜癌組織中的表達水平顯著低于癌旁組織(P<0.01,圖1)。

圖1 ACTA2-AS1在子宮內膜癌組織和癌旁組織中的表達水平
2.ACTA2-AS1在子宮內膜癌細胞株中的表達:qRT-PCR檢測顯示,ACTA2-AS1在子宮內膜癌細胞(HEC-1A、Ishikawa、AN3CA、HEC-1B)及正常子宮內膜上皮細胞ESC的表達分別為0.58±0.07、0.41±0.05、0.18±0.06、0.66±0.08和1.05±0.09(圖2)。與ESC細胞比較,ACTA2-AS1在子宮內膜癌細胞株中的表達明顯下降(P<0.05),AN3CA細胞ACTA2-AS1表達下降最明顯(P<0.01),因而采用AN3CA細胞進行研究。

圖2 ACTA2-AS1在子宮內膜癌細胞株和正常子宮內膜上皮細胞中的表達水平與ESC細胞比較,*P<0.05,**P<0.01
3.轉染ACTA2-AS1過表達質粒對AN3CA細胞中ACTA2-AS1表達的影響:轉染ACTA2-AS1過表達質粒后,ACTA2-AS1組和NC組AN3CA細胞中ACTA2-AS1表達分別為12.26±1.12和1.21±0.20,ACTA2-AS1組ACTA2-AS1表達顯著高于NC組(P<0.01)。
4.上調ACTA2-AS1對AN3CA細胞周期的影響:流式細胞法結果顯示,ACTA2-AS1組和NC組AN3CA細胞在G0~G1期的細胞比例分別為61.34%±4.51%和29.73%±3.39%,上調ACTA2-AS1能明顯提升G0~G1期細胞比例,誘導G0~G1期停滯(P<0.05,圖3)。
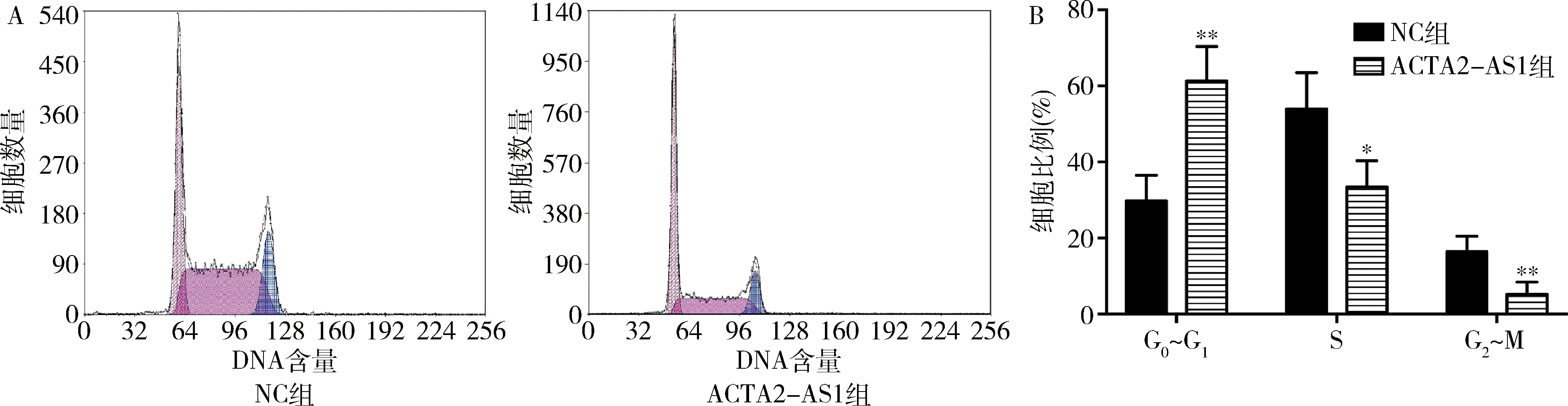
圖3 上調ACTA2-AS1對AN3CA細胞周期的影響A.流式細胞法;B.細胞周期比例分析。與NC組比較,*P<0.05,**P<0.01
5.上調ACTA2-AS1對AN3CA細胞遷移能力的影響:細胞劃痕實驗結果顯示,ACTA2-AS1組和NC組AN3CA細胞的遷移率分別為40.46%±3.76%和69.54%±6.64%,上調ACTA2-AS1可抑制AN3CA細胞的遷移能力(P<0.01,圖4)。
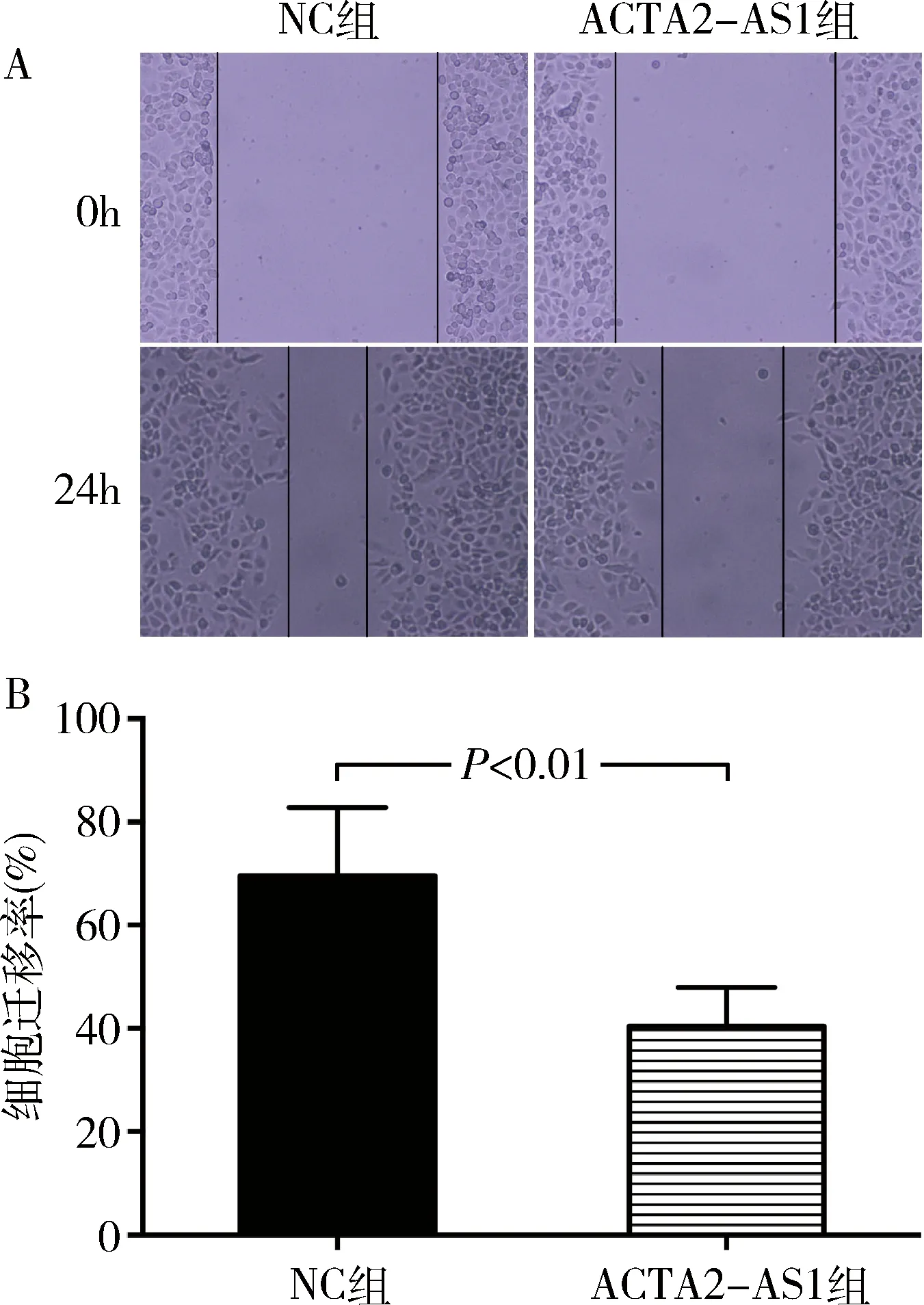
圖4 上調ACTA2-AS1對AN3CA細胞遷移能力的影響A.細胞劃痕實驗(×100);B.細胞遷移率定量分析
6.ACTA2-AS1對miR-586的靶向位點預測:采用LncBase v.2數據庫預測ACTA2-AS1的潛在靶基因,ACTA2-AS1與miR-586具有潛在的互補結合位點(圖5)。

圖5 LncBase v.2數據庫預測ACTA2-AS1的潛在靶基因
7.雙熒光素酶報告基因檢測驗證ACTA2-AS1對miR-586的靶向調控:雙熒光素酶報告基因檢測結果顯示,在AN3CA細胞中,與共轉染ACTA2-AS1-wt和miR-NC比較,共轉染ACTA2-AS1-wt和miR-586后相對熒光素酶活性明顯下降(P<0.01),ACTA2-AS1能夠靶向調控miR-586(圖6)。

圖6 雙熒光素酶報告基因法驗證ACTA2-AS1對miR-586的靶向調控與miR-NC組比較,*P<0.01
8.上調ACTA2-AS1對miR-586和PTEN mRNA表達的影響:qRT-PCR檢測結果顯示,ACTA2-AS1組和NC組AN3CA細胞中miR-586表達分別為0.21±0.05和1.12±0.09,上調ACTA2-AS1可抑制miR-586表達(P<0.01)。ACTA2-AS1組和NC組AN3CA細胞中PTEN mRNA表達分別為6.11±1.76和1.23±0.53,上調ACTA2-AS1可促進PTEN mRNA表達(P<0.01)。
9.上調ACTA2-AS1對PTEN蛋白和PI3K-AKT-mTOR信號通路蛋白表達的影響:Western blot法檢測結果顯示,上調ACTA2-AS1表達后,PTEN蛋白表達升高,PI3K-AKT-mTOR信號通路蛋白p-PI3K、p-AKT、p-mTOR表達降低(圖7)。
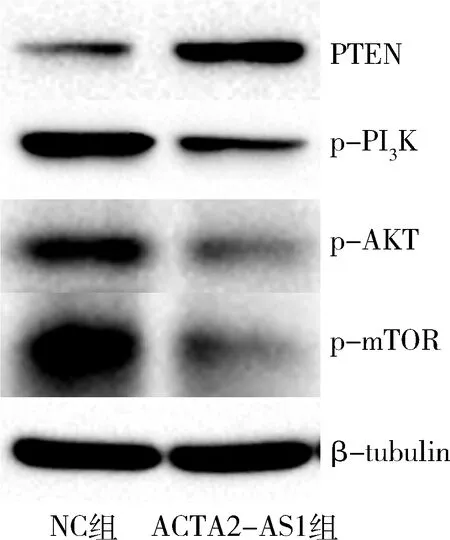
圖7 上調ACTA2-AS1對AN3CA細胞PTEN蛋白和PI3K-AKT-mTOR信號通路蛋白表達的影響
討 論
lncRNA在細胞內發揮廣泛的基因表達調節功能,其異常表達可誘導細胞活動的紊亂,引起腫瘤的發生[9]。不同的lncRNA在同一種腫瘤細胞中的功能不同,可能為抑癌基因,也可能為癌基因[10]。lncRNA如LA16c-313D11.11在子宮內膜癌組織中的表達水平顯著降低,其可充當miR-205-5p的海綿分子,間接增加PTEN基因的表達,抑制子宮內膜癌細胞的侵襲、遷移和活力[11]。lncRNA如RHPN1-AS1、UCA1、DLX6-AS1、CHL1-AS1等在子宮內膜癌組織和細胞株中表達均顯著上調,與子宮內膜癌的分期、組織學分級、淋巴結轉移、不良預后等顯著相關,促進子宮內膜癌細胞的周期進展、遷移以及侵襲[12~15]。近年來,ACTA2-AS1已被證實與多種腫瘤的發生和發展有關。Ying等[8]研究發現,ACTA2-AS1在肺腺癌組織和細胞株中低表達,上調ACTA2-AS1可抑制肺腺癌細胞的上皮間充質轉化并誘導細胞凋亡。其可能成為垂體泌乳素瘤發病機制的新型生物學標志物。Pan等[6]研究發現,ACTA2-AS1在結腸腺癌組織和細胞株中表達降低,其可明顯抑制結腸腺癌細胞的活力和集落形成能力。
ACTA2-AS1在子宮內膜癌中的作用鮮有報道。本研究通過GEPIA數據庫分析發現,ACTA2-AS1在子宮內膜癌組織中的表達水平明顯低于癌旁組織,且ACTA2-AS1在子宮內膜癌細胞株中的表達水平明顯低于正常子宮內膜上皮細胞,提示ACTA2-AS1表達的異常改變與子宮內膜癌的發生存在相關性。本研究利用ACTA2-AS1表達最低的AN3CA細胞進行了系列細胞實驗,上調ACTA2-AS1表達可以促進G0~G1期細胞比例增加、抑制細胞的遷移能力,ACTA2-AS1在子宮內膜癌中發揮抑癌基因的作用。lncRNA通過與靶微小RNA(miRNA)以不完全或完全的堿基配對方式,降低miRNA的表達,調控細胞周期、凋亡、遷移等生物過程[16, 17]。研究ACTA2-AS1的作用機制,重點在于探究ACTA2-AS1與靶miRNA的相互結合。本研究通過LncBase v.2數據庫發現,ACTA2-AS1與miR-586可能互補結合。Yang等[18]研究發現,miR-586可以促進骨肉瘤細胞的增殖、侵襲、轉移,同時抑制骨肉瘤細胞的凋亡,發揮癌基因作用。
本研究通過雙熒光素酶報告基因實驗證實ACTA2-AS1可互補結合miR-586。qRT-PCR結果也證明了,ACTA2-AS1能負性調控miR-586的表達。這些結果提示ACTA2-AS1能直接靶向作用miR-586。Yu等[19]研究發現,miR-586通過靶向結合PTEN mRNA的非翻譯區,干擾PTEN基因表達,促進癌癥的發生。本研究中qRT-PCR和Western blot法均顯示上調ACTA2-AS1后,PTEN基因表達降低,進一步證明ACTA2-AS1靶向作用miR-586。Zheng等[20]研究發現,PTEN通過影響PI3K-AKT-mTOR信號通路的活化,抑制子宮內膜癌的進展。本研究中Western blot法檢測結果顯示,上調ACTA2-AS1后,PI3K-AKT-mTOR信號通路活化被抑制。
綜上所述,本研究觀察到ACTA2-AS1在子宮內膜癌組織和細胞株中存在異常低表達。對于存在PI3K-AKT-mTOR信號通路的子宮內膜癌類型,上調ACTA2-AS1表達通過靶向調控miR-586,促進PTEN基因表達和抑制PI3K-AKT-mTOR信號通路活化,抑制子宮內膜癌細胞周期和遷移。ACTA2-AS1可能為子宮內膜癌的靶向治療提供了新的靶點和實驗依據。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
