豬NOX1基因啟動子區轉錄因子的篩選與鑒定
2021-12-23 11:16:26李忠秋何鑫淼王文濤唐曉東張冬杰
中國獸醫學報 2021年11期
汪 亮,李忠秋,何鑫淼,王文濤,唐曉東,劉 娣,張冬杰*
(1.黑龍江省農業科學院,黑龍江 哈爾濱 150086;2.農業農村部種養結合重點實驗室,黑龍江 哈爾濱 150086)
還原型煙酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶(NADPH oxidase,NOX)是一個多基因家族,目前已知有7名成員,包括NOX1~5和Duo x1~2,其中NOX2是最早發現的。NOX是一種由多個蛋白質亞基組成的復合物,其主要亞基就是NOX2(gp91-phox),NOX2特異表達于吞噬細胞,參與機體抗感染免疫防御[1]。后來,人們在非吞噬細胞中發現了gp91-phox的同源分子,即NOX1,它可在多個組織中表達,如大腸、前列腺、子宮和睪丸等。在血管平滑肌細胞中含量較高,許多刺激物如血小板衍生生長因子(platelet derived growth factor,PDGF),前列腺素 F2α(prostaglandin F2α,PG F2α)等均能誘導NOX1 mRNA 的表達增加[2]。將該基因在小鼠成纖維細胞NIH3T3過表達后,細胞內活性氧(reactive oxygen species,ROS)的產出增加,細胞增殖明顯,并在細胞團中形成血管[3]。細胞較高的ROS水平一般與惡性轉化有關,腫瘤細胞的ROS水平通常都會高于非腫瘤細胞。因此,NOX1與肺病、心血管系統疾病和腫瘤等疾病相關,不僅參與組織氧化損傷,還與組織中的炎癥反應有關[4]。
本課題組發現6月齡雌性民豬在遭受-15℃持續14 d的冷處理后,其背部皮下脂肪內的NOX1基因轉錄水平與對照組相比發生了5.9倍的上升,推測其可能受低溫環境誘導,并在脂肪細胞的新陳代謝中發揮作用。為了后續解析NOX1受低溫誘導的分子機理,本研究擬利用雙熒光素酶報告系統篩選調控其轉錄的分子元件,探討其轉錄調控機制。
1 材料與方法
1.1 材料豬基因組DNA和PK15細胞系由本實驗室保存;限制性核酸內切酶KpnⅠ、Hind Ⅲ和pMD18-T 載體、T4連接酶等購自TaKaRa公司;膠回收試劑盒(EG101-02)購自北京全式金生物公司;無內毒素質粒提取試劑盒購自天根生化科技有限公司;pGL3-Basic載體和雙熒光素酶檢測試劑盒購自Promega;脂質體2000購自Invitrogen。
1.2 引物設計與PCR擴增根據NCBI數據庫中已有的豬NOX1基因組序列(NC_010461)設計引物1618S和1618A,擴增-1 618~-1 bp區間(將起始密碼子“A”的位置定義為1)。1618S:5′-GGTACCACAATCTCATGTT CTCACCT-3′,16-18A:5′-AAGCTTTGTCAAGAGGTGTTTTGG-3′。下劃線表示人為添加的酶切位點,分別為KpnⅠ和Hind Ⅲ。PCR反應體系為:DNA(50 mg/L)1 μL;2×PCR Mix 10 μL;上下游引物(10 μmol/L)各0.5 μL;去離子水補齊至20 μL。反應條件:95℃ 5 min;95℃ 30 s,60℃ 30 s,72℃ 2 min,30個循環;72℃ 5 min;16℃ 30 min。PCR反應結束后,使用1%的瓊脂糖凝膠電泳檢測PCR產物,凝膠成像系統觀察、記錄。
1.3 克隆測序嚴格按照膠回收試劑盒的說明書操作,將回收后的PCR產物連入pMD18-T載體,轉入DH5α感受態細胞后,經挑菌、搖菌和提質粒后獲得pMD18-NOX1質粒。使用KpnⅠ和Hind Ⅲ雙酶切鑒定。雙酶切體系為:KpnⅠ和Hind Ⅲ各0.5 μL,10×Buffer 1 μL,質粒4 μL;去離子水4 μL。37℃酶切15 min。酶切結束后使用1%瓊脂糖凝膠電泳檢測,選取含有目的條帶的菌落送至北京擎科生物科技有限公司測序。使用DNAMAN 7.0軟件對測序結果進行分析。
1.4 熒光素酶報告基因質粒構建將正確連入NOX1基因的pMD18-NOX1質粒與pGL3-Basic載體分別同時使用KpnⅠ和Hind Ⅲ雙酶切,回收目的片段,使用T4連接酶將NOX1片段與pGL3-Basic載體相連接,轉化至DH5α感受態細胞中,在含有氨芐的LB固體培養基中篩選重組質粒,經挑菌、搖菌、菌液PCR、雙酶切、測序驗證后,提取無內毒素的pGL3-Basic-1618質粒,-20℃保存備用。
1.5 PK15細胞的體外培養在含有10%胎牛血清、90% DMEM培養基、鏈霉素(100 U/mL)和青霉素(100 U/mL)中培養PK15細胞并置于5% CO2、37℃的飽和濕度培養箱中,待細胞生長匯合至85%~90%且呈伸展狀態時,用0.25%胰蛋白酶充分消化10~15 min,以胎牛血清終止消化,傳代于24孔板中,過夜培養,第2天進行轉染試驗。

1.7 NOX1基因啟動子核心區鑒定以獲得的-1 618~-1 bp片段為基序列,下游引物不變,單獨設計上游引物,每條引物之間相差300 bp左右,5′端均引入KpnⅠ酶切位點(表1),共組成12對引物。以pMD18-T-NOX1為模板,分別進行PCR擴增,將大小正確的擴增片段回收,分別連入pMD18-T載體,測序驗證正確后,將其從載體中雙酶切下來,連入pGL3-Basic載體,轉染入PK15細胞,進行熒光素酶活性檢測。

表1 擴增截短型啟動子所用引物信息
1.8 NOX1基因啟動子區調控元件鑒定為保證預測的準確性,分別在JASPAR(http://jaspar.genereg.net)、ALGGEN(http://alggen.lsi.upc.es/)和Tfsitescan(http://www.ifti.org/Tfsitescan/)網站對NOX1基因的啟動子區進行調控元件的預測。同時結合不同片段熒光素酶活性的測定結果,預測調控因子的結合位點。使用重疊延伸PCR技術將這些預測的結合位點進行缺失突變,所用引物序列見表2。以第1組引物M-1149-a2(A)和M-1149-a3(S)為例,第一輪反應以1149S和M-1149-a2(A)、M-1149-a3(S)和1618A引物對擴增,第二輪反應以第一輪反應獲得的2個片段為模板,使用1149S和1618A引物對進行拼接擴增,將缺失突變引入產物內部。將擴增產物采用同上的方法連入pGL3-Basic載體,熒光素酶活性的測定也同上。

表2 突變調控元件所用引物信息
2 結果
2.1 NOX1基因啟動子區的擴增與鑒定利用引物1618S和1618A進行PCR擴增,瓊脂糖凝膠電泳檢測后在1 618 bp左右發現目的條帶,經測序驗證后表明所擴增的片段為豬的NOX 1基因啟動子區(圖1A)。
2.2 pGL3-Basic-1618熒光素酶報告基因質粒的構建利用Kpn Ⅰ和 Hind Ⅲ對pMD18-T-NOX1質粒及pGL3-Basic空載體進行雙酶切處理,將純化后的目的片段連入線性化載體得到重組質粒pGL3-Basic-1618。對pGL3-Basic-1618質粒雙酶切(圖1B)、測序鑒定后表明該片段正確的連入了pGL3-Basic載體。

A.NOX1基因的PCR擴增結果;B.NOX1重組質粒的雙酶切結果;M.DL2000 DNA Marker;M1.DL4000 DNA Marker;1.重組質粒圖1 NOX1基因的PCR擴增及雙酶切鑒定結果
2.3 pGL3-Basic-1618質粒的熒光素酶活性檢測將pGL3-Basic-1618無內毒素質粒轉染入PK15細胞中,24 h后檢測熒光素酶活性,結果顯示1618(圖2)片段與空載體相比,熒光素酶相對活性顯著升高,表明該片段含有轉錄因子結合位點,具有轉錄調控功能。

圖2 pGL3-Basic-1618熒光素酶活性分析
2.4 NOX1基因啟動子區系列截短序列載體的構建以pMD18-T-NOX1片段為模板,利用12條截短序列的上游引物與1618A配對,共擴增出12條長度不同的片段。將這些片段分別與pMD18-T載體相連,經測序驗證后,從T載體上將目的片段切下,分別連入pGL3-Basic載體,構建12個報告基因重組載體,分別為pGL3-Basic-1326、pGL3-Basic-1251、pGL3-Basic-1185、pGL3-Basic-1149、pGL3-Basic-1091、pGL3-Basic-1052、pGL3-Basic-968、pGL3-Basic-834、 pGL3-Basic-719、pGL3-Basic-325、pGL3-Basic-208和pGL3-Basic-115。經雙酶切(圖3)和測序驗證后表明目的片段均正確連入pGL3-Basic載體。
2.5 系列截短序列的雙熒光素酶活性分析第一輪熒光素酶活性分析主要是從1618 bp片段至325 bp 片段,區間間隔大約為300 bp長度,共檢測了5個片段的熒光素酶活性(圖4)。根據熒光素酶的相對活性,發現1 618 bp片段截短至1 326 bp后,啟動子活性無顯著變化,說明-1 618~-1 326 bp區域內沒有重要的轉錄調控位點。將1 326 bp片段截短至1 091 bp 后,啟動子活性顯著降低(P<0.05),1 091 bp片段截短至719 bp后,啟動子活性明顯升高,說明-1 326~-1 091 bp和-1 091~-719 bp 區域內分別存在正調控元件和負調控元件。719 bp片段截短至325 bp后,啟動子活性無顯著變化;但325 bp片段的雙熒光素酶活性依舊顯著,說明-325~-1 bp間存在增強子。

圖4 NOX1基因啟動子區第一輪雙熒光素酶活性檢測結果
第二輪熒光素酶活性分析以上述發生顯著變化的區間為基礎,將-1 326~-1 091 bp、-1 091~-719 bp和-325~-1 bp這3個區間進一步分割,結果發現-1 149~-1 091 bp和-208~-115 bp區域內可能存在正調控元件,-1 091~-1 052 bp區域內可能存在負調控元件,這3個區域存在對NOX1基因轉錄影響較大的轉錄因子結合位點(圖5)。
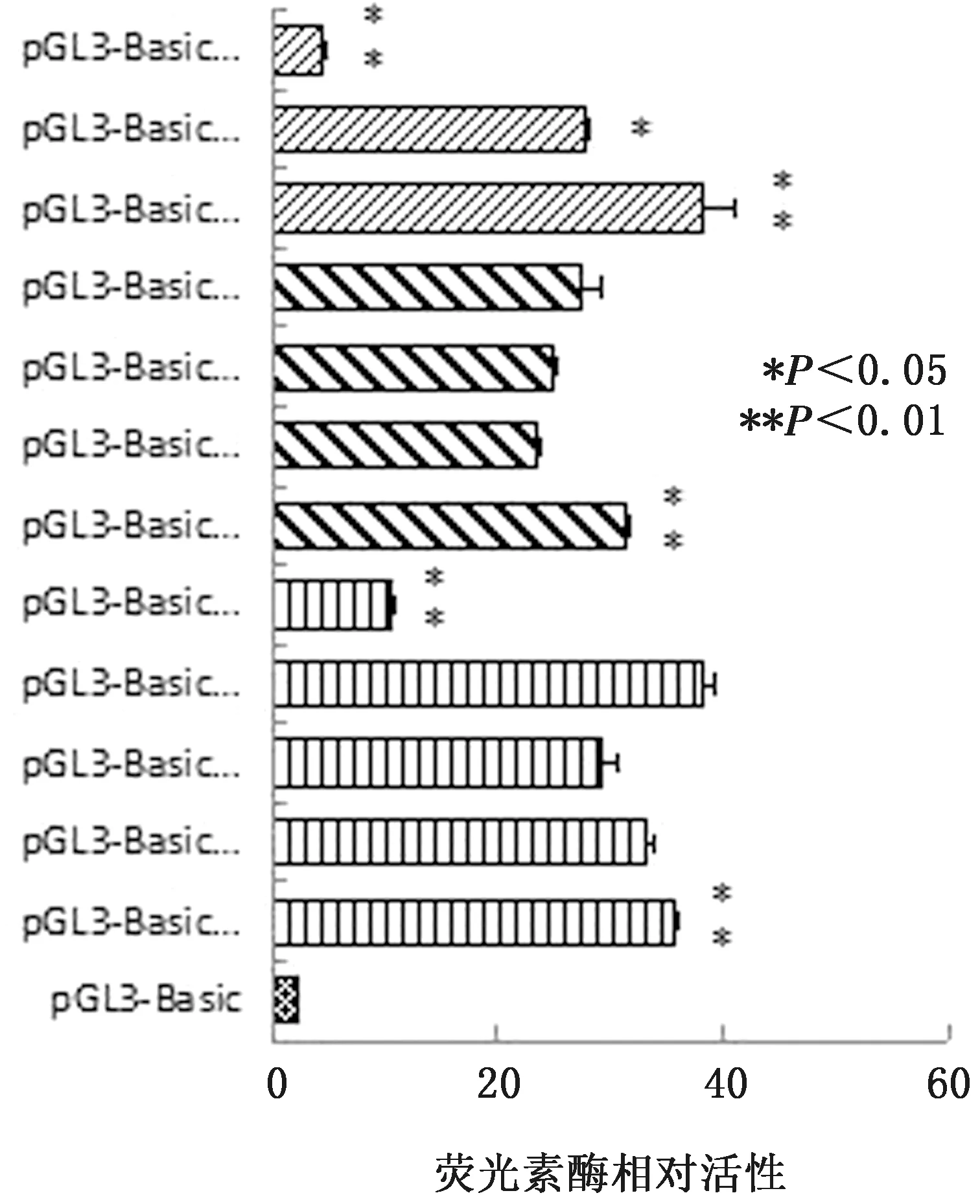
圖5 NOX1基因啟動子區第二輪雙熒光素酶活性檢測結果
2.6 NOX1基因轉錄調控元件的預測與鑒定
2.6.1生物信息學預測結果 綜合3個在線預測軟件的分析結果,同時參考上述熒光素酶相對活性的測定結果,共篩選出11個轉錄因子結合位點(表3),依據這些轉錄因子的結合位點,選擇了適當的突變位置,以判定轉錄因子是否會與這些位點結合。

表3 轉錄因子結合位點預測結果
2.6.2轉錄因子結合位點的驗證 分別以pGL3-Basic-1326和pGL3-Basic-208為模板,利用重疊延伸PCR法對預測的結合位點進行缺失突變,經酶切測序驗證后,獲得了4個報告基因質粒pGL3-Basic-1104、pGL3-Basic-1051、pGL3-Basic-1043和pGL3-Basic-126(數字代表缺失起始位置)。
以pGL3-Basic-1326和pGL3-Basic-208的雙熒光素酶活性為參照,將構建的重組質粒進行雙熒光素酶活性檢測。結果顯示缺失-1 104~-1 092 bp區域,雙熒光素酶活性發生顯著降低,其他2個區間的突變無明顯變化,缺失-126~-116 bp區域雙熒光素酶活性顯著降低,推測ZNF384,ELF-1和TBR1是豬NOX1基因的正調控轉錄因子(圖6)。

圖6 序列缺失對NOX1基因啟動子活性的影響
3 討論
基因轉錄是在細胞核和細胞質內進行的,它是指以DNA的一條鏈為模板,按照堿基互補配對原則,合成RNA的過程。基因轉錄有正調控和負調控之分。轉錄因子是起調控作用的反式作用因子,它是轉錄起始過程中RNA聚合酶所需的輔助因子。真核生物基因在無轉錄因子時處于不表達狀態,RNA聚合酶自身無法啟動基因轉錄,只有當轉錄因子(蛋白質)結合在其識別的DNA序列上后,基因才開始表達。轉錄因子的結合位點(transcription factor binding site,TFBS)是與轉錄因子結合的DNA片段,長度通常在5~20 bp,一個轉錄因子往往同時調控若干個基因,在不同基因上的結合位點具有一定的保守性。較短的DNA片段在規模較大的基因組中重復出現的次數很多,另外,TFBS又允許一定的可變性,這給識別TFBS的工作帶來困難。使得預測TFBS的算法普遍存在假陽性率偏高的問題[5],因此,預測后的結果必須輔以試驗驗證。
NOX1是NADPH氧化酶的一個催化亞基,在血管平滑肌細胞中含量較高,與血管硬化以及缺血再灌注損傷相關,也介導血管平滑肌細胞的增生和肥大[6]。Ang Ⅱ能夠誘導NOX1 mRNA表達的增加,而二聯苯碘(dipheny leneiodo nium,DPI)、線粒體呼吸鏈復合體-Ⅰ的抑制劑等則可抑制NOX1 mRNA的表達[7]。上調NOX1表達和隨后生成的氧化應激產物可促進癌細胞增殖,如果抑制NOX1的活性則可誘導許多癌細胞(包括胰腺癌、間皮瘤和肉瘤)凋亡[8],這也預示著NOX1可能是癌細胞治療的一個重要潛在靶點。在豬上的研究表明,羅伊氏乳桿菌可刺激斷奶仔豬空腸組織內NOX1基因的顯著上調[9]。運輸應激可造成育肥豬空腸組織內NOX1 mRNA的顯著上升[10],進一步研究發現,抑制NOX1表達可降低ROS和自噬水平,緩解氧化應激對細胞造成的損傷[11]。
本研究對豬的NOX1基因啟動子區可能存在的轉錄因子進行了篩選與鑒定,發現該基因啟動子區的-1 149~-1 091 bp和-208~-115 bp區域內存在正調控元件,-1 091~-1 052 bp區域內存在負調控元件。-1 104~-1 092 bp和-126~-116 bp 區域存在正調控該基因轉錄的轉錄因子結合位點,通過在線軟件預測及靶序列定點缺失驗證后認為ZNF384,ELF-1和TBR1可能是該基因的正調控轉錄因子。目前已知鋅指蛋白384(ZNF384)可結合并調節細胞外基質基因MMP1、MMP3、MMP7 及COL1A1的啟動子[12]。ELF-1是ETS轉錄因子家族的一員,是具有啟動子特性的轉錄因子,在多種腫瘤中表達,與腫瘤的增殖、分化、凋亡、遷移等相關[13]。TBR1是來自于T-box腦蛋白家族的1個轉錄因子,它不但參與大腦皮質的早期發育,而且與神經細胞的分化與遷移、軸突尋徑以及片層化的發育都有重要的關系[14],但上述轉錄因子是否會受到冷環境誘導,還未見報道。
徐采云等[15]在對人NOX1基因啟動子區序列分析時發現,-140~-419 bp和-577~-946 bp區域可能存在NOX1啟動子核心區域,與本研究所錨定的區域并不完全相同。此外,人們還發現轉錄因子SP1參與TNF-α介導的肺泡上皮細胞NOX1基因的表達調控[16],沉默NF-κB/p65基因,可有效下調TNF-α誘導A549細胞氧化應激程度及NOX1基因表達水平[17-18]。GLE1、DDX19A、KRT1和KRT10參與TNF-α誘導的A549細胞NOX1活化[19]。由此可知,NOX1基因的轉錄調控非常復雜,在不同類型的細胞內及行使不同生物學功能時其轉錄調控均存在差異。
綜上所述,本研究克隆了豬NOX1基因的啟動子序列,對可能結合到該區間的轉錄因子進行了篩選與分析,初步認定ZNF384、ELF-1和TBR1為該基因轉錄的正調控因子。結合前人的研究結果分析,NOX1基因的轉錄調控機制比較復雜。
