可誘導(dǎo)表達(dá)結(jié)核分枝桿菌ESAT-6的THP-1細(xì)胞系的建立
2021-12-23 11:16:14唐亦然戴鑫鈞胡燕萍畢斯琪孟祥苗宋厚輝
中國獸醫(yī)學(xué)報(bào) 2021年11期
關(guān)鍵詞:檢測
唐亦然,戴鑫鈞,胡燕萍,畢斯琪,孟祥苗,楊 楊,宋厚輝
(浙江農(nóng)林大學(xué) 動(dòng)物科技學(xué)院/動(dòng)物醫(yī)學(xué)院 浙江省畜禽綠色生態(tài)健康養(yǎng)殖應(yīng)用技術(shù)研究重點(diǎn)實(shí)驗(yàn)室/動(dòng)物健康互聯(lián)網(wǎng)檢測技術(shù)浙江省工程實(shí)驗(yàn)室,浙江 杭州 311300)
盡管結(jié)核分枝桿菌(Mycobacteriumtuberculosis,MTB)早在100多年前就被發(fā)現(xiàn),但其所造成的結(jié)核病仍持續(xù)流行,成為了全球性公共衛(wèi)生問題,對人類的健康造成嚴(yán)重危害[1]。6 kDa早期分泌抗原靶標(biāo)(6-kDa early secretory antigenic target,ESAT-6) 是MTB獨(dú)有分泌系統(tǒng)ESX-1分泌的小分子蛋白,該基因位于MTB RD-1區(qū)的開放閱讀框的Rv3875處[2-3]。RD-1區(qū)僅存在于致病性分枝桿菌中,包括MTB、牛分枝桿菌、非洲分枝桿菌等,而不存在于所有減毒BCG疫苗株中,其與毒力密切相關(guān)[4]。作為MTB重要的毒力因子,ESAT-6參與MTB的致病過程,包括促進(jìn)MTB在宿主細(xì)胞內(nèi)復(fù)制與細(xì)胞間的擴(kuò)散、誘導(dǎo)細(xì)胞凋亡發(fā)生、成孔和裂解膜、引起Ⅰ型IFN釋放等[5]。ESAT-6可以增加細(xì)胞內(nèi)Ca2+的濃度,促進(jìn)活性氧(ROS)的產(chǎn)生,從而觸發(fā)內(nèi)質(zhì)網(wǎng)的激活,調(diào)控內(nèi)質(zhì)網(wǎng)介導(dǎo)的下游信號通路eIF2a/ATF4/CHOP引起細(xì)胞凋亡的發(fā)生[6-7]。另外,ESAT-6具有成孔功能,可以導(dǎo)致細(xì)胞中吞噬溶酶體的破裂,從而使MTB從吞噬體向細(xì)胞質(zhì)轉(zhuǎn)運(yùn),從而激活一系列的胞內(nèi)反應(yīng)[8-9]。
本試驗(yàn)首次采用Tet-On 3G誘導(dǎo)表達(dá)系統(tǒng)中的反應(yīng)質(zhì)粒pLVX-TRE3G和調(diào)控質(zhì)粒pLVX-Tet3G,將ESAT-6基因和紅色熒光蛋白mCherry融合插入到pLVX-TRE3G中,與調(diào)控質(zhì)粒pLVX-Tet3G分別包裝成慢病毒后感染THP-1細(xì)胞,在Puromycin和G418雙重壓力下篩選可經(jīng)Dox誘導(dǎo)表達(dá)ESAT-6的細(xì)胞系,并分析不同濃度的Dox和不同誘導(dǎo)的時(shí)間對THP-1中ESAT-6表達(dá)的影響以及誘導(dǎo)表達(dá)蛋白ESAT-6的生物活性功能,為深入研究ESAT-6在MTB感染中的作用機(jī)制奠定了基礎(chǔ)。
1 材料與方法
1.1 材料大腸桿菌DH5α由本實(shí)驗(yàn)室保存。質(zhì)粒pLVX-TRE3G、pLVX-Tet3G、pMD2.0G和psPAX2購自長沙優(yōu)寶生物科技有限公司。真核表達(dá)質(zhì)粒pmCherry-ESAT-6為本試驗(yàn)自行構(gòu)建,在質(zhì)粒pmCheryy-N1的基礎(chǔ)上插入了MTBESAT-6基因片段。限制性內(nèi)切酶SmaⅠ和EcoRⅠ購自NEB公司;DL2000 DNA Marker、DNA連接酶購自TaKaRa公司;DNA聚合酶2×Phanta Max Master Mix購自南京諾維贊生物科技有限公司;轉(zhuǎn)染試劑Lipofectamine 2000購自Invitrogen公司;強(qiáng)力霉素(Doxycycline,Dox)、嘌呤霉素(Puromycin)、G418購自Sigma-Aldrich公司;南美胎牛血清(FBS)和細(xì)胞培養(yǎng)基(DMEM、1640)購自Gibco公司;HRP標(biāo)記的山羊抗鼠 IgG、山羊抗兔 IgG以及Human IL-1β ELISA試劑盒購自上海優(yōu)寧維公司;mCherry單克隆抗體購自Abcam公司;脂多糖LPS和尼日利亞菌素Nigericin;CellTiter-Glo Luminescent Cell Viability Assay試劑盒購自Promega。
1.2 細(xì)胞培養(yǎng)人腎上皮細(xì)胞HEK-293T用含有10%的胎牛血清的DMEM培養(yǎng)基培養(yǎng),人單核細(xì)胞THP-1用含有10%的胎牛血清的1640培養(yǎng)基培養(yǎng),均置于含有5% CO2的37℃培養(yǎng)箱中。
1.3 mCherry-ESAT-6融合基因的擴(kuò)增以實(shí)驗(yàn)室構(gòu)建的質(zhì)粒pmCherry-ESAT-6為模板,設(shè)計(jì)特異性引物(序列如表1所示)用來PCR擴(kuò)增ESAT-6和熒光標(biāo)簽mCherry的融合片段。反應(yīng)體系如下:DNA模板1 μL,2×Phanta Max Master Mix 25 μL,上下游引物各1 μL,ddH2O補(bǔ)足至50 μL。設(shè)置PCR反應(yīng)條件如下:預(yù)熱94℃ 3 min,變性94℃ 30 s,退火60℃ 30 s,延伸68℃ 30 s,循環(huán)35次,延伸68℃ 7 min。經(jīng)1%瓊脂糖核酸電泳驗(yàn)證PCR產(chǎn)物。

表1 引物序列
1.4 重組質(zhì)粒pTRE3G-mCherry-ESAT-6的構(gòu)建使用限制性內(nèi)切酶SmaⅠ和EcoRⅠ酶切質(zhì)粒pLVX-TRE3G以及 ESAT-6基因的PCR產(chǎn)物,37℃孵育4 h。純化回收酶切產(chǎn)物后,用DNA連接酶16℃連接2 h,轉(zhuǎn)到大腸桿菌DH5α感受態(tài)細(xì)胞中,并涂布于含Kana抗性的LB平板上過夜培養(yǎng)。挑取單菌落,經(jīng)菌落PCR篩選陽性克隆,擴(kuò)大培養(yǎng)后提取質(zhì)粒,將重組質(zhì)粒送至浙江有康生物科技有限公司測序。將測序成功的重組質(zhì)粒命名pTRE3G-mCherry-ESAT-6。
1.5 重組質(zhì)粒誘導(dǎo)表達(dá)目的蛋白的初步驗(yàn)證將HEK293T細(xì)胞以5×105個(gè)/mL的細(xì)胞密度鋪于12孔細(xì)胞培養(yǎng)板中,過夜培養(yǎng)。將質(zhì)粒pTRE3G-mCherry-ESAT-6和pLVX-Tet3G按照質(zhì)量比 4∶1混合,用Lipofectamine 2000轉(zhuǎn)染試劑轉(zhuǎn)染到細(xì)胞中,同時(shí)加入100 μg/L的 Dox。轉(zhuǎn)染24 h后,通過熒光顯微鏡觀察熒光表達(dá)情況。加入200 μL NP40裂解細(xì)胞,收取細(xì)胞總蛋白,用10%的分離膠進(jìn)行SDS-PAGE后,105 V轉(zhuǎn)膜40 min;5%的脫脂奶粉封閉1 h,TBST洗滌3次,5 min/次;用mCherry單克隆抗體(1∶1 000稀釋)4℃孵育過夜后,TBST洗滌5次;用 HRP標(biāo)記的山羊抗兔室溫孵育1 h,TBST洗滌3次;用ECL發(fā)光底物進(jìn)行顯影曝光。
1.6 慢病毒包裝將 HEK293T細(xì)胞以5×105個(gè)/mL的細(xì)胞密度鋪于10 cm培養(yǎng)皿中,過夜培養(yǎng)12~16 h。將pTRE3G-mCherry-ESAT-6與慢病毒包裝質(zhì)粒psPAX2和pMD2.0G以2∶2∶1的比例混合后,用Lipofectamine 2000轉(zhuǎn)染上述混合質(zhì)粒到細(xì)胞中,轉(zhuǎn)染3 d后收集含病毒的上清,并且用0.45 μm濾器過濾除菌,所獲得病毒命名為LV-TRE3G-ESAT-6。用上述相同方法獲取LV-Tet3G慢病毒上清,分裝保存于-80℃。
1.7 可誘導(dǎo)表達(dá)目的蛋白的THP-1細(xì)胞系的篩選與鑒定將THP-1細(xì)胞以每孔5×105個(gè)/mL的細(xì)胞密度鋪于6孔細(xì)胞培養(yǎng)板中,過夜培養(yǎng)。吸去原有培養(yǎng)基,加入含有病毒的培養(yǎng)基(1 mL培養(yǎng)基中含有6 mg/L的polybrene、50 μL LV-TRE3G-ESAT-6和50 μL LVX-Tet3G),300 r/min離心1 h后,再加入1 mL培養(yǎng)基(含有6 mg/L的polybrene)。感染第2天換掉含病毒的培養(yǎng)基,待感染72 h后,換含有1 mg/L Puromycin和800 mg/L G418的新鮮完全培養(yǎng)液,同時(shí)設(shè)置空白對照。每2~3 d更換1次含有抗生素的培養(yǎng)基,直到空白組細(xì)胞全部死亡。陽性細(xì)胞克隆擴(kuò)大培養(yǎng)、傳代、凍存,將所獲得的細(xì)胞株命名為THP-1/ESAT-6。將THP-1/ESAT-6細(xì)胞鋪于24孔細(xì)胞培養(yǎng)板中,添加100 μg/L的Dox誘導(dǎo)劑,培養(yǎng)24 h后,提取細(xì)胞的總蛋白,Western blot檢測 ESAT-6表達(dá)情況。加入不同濃度的Dox培養(yǎng)24 h或者用100 μg/L的Dox培養(yǎng)不同時(shí)間后,Western blot檢測 ESAT-6的表達(dá)與誘導(dǎo)劑濃度和作用時(shí)間的關(guān)系。
1.8 可誘導(dǎo)表達(dá)目的蛋白的THP-1細(xì)胞系穩(wěn)定性檢測為了檢測不同代次細(xì)胞系的ESAT-6蛋白的表達(dá)穩(wěn)定性,在將細(xì)胞傳至第2,8,14和20代時(shí)分別鋪于24孔細(xì)胞培養(yǎng)板中,添加100 μg/L的Dox,誘導(dǎo)表達(dá)24 h后進(jìn)行Western blot檢測 ESAT-6的表達(dá)。
1.9 可誘導(dǎo)表達(dá)目的蛋白的生物活性檢測將篩選獲得的細(xì)胞系THP-1/ESAT-6鋪于96孔細(xì)胞培養(yǎng)板中, 加入100 μg/L的Dox分別處理6,9,12,18,24,36和48 h后,通過CellTiter-Glo Luminescent Cell Viability Assay試劑盒測定細(xì)胞的存活率。此外將THP-1/ESAT-6細(xì)胞鋪于24孔細(xì)胞培養(yǎng)板中,添加100 μg/L的Dox誘導(dǎo)劑培養(yǎng)20 h后,加入脂多糖1 mg/L LPS刺激細(xì)胞3 h,10 μmol/L尼日利亞菌素Nigericin作用1 h后,收集細(xì)胞上清,通過ELISA試劑盒檢測細(xì)胞上清中的IL-1β的分泌情況。

2 結(jié)果
2.1 ESAT-6基因的PCR擴(kuò)增以實(shí)驗(yàn)室構(gòu)建的真核表達(dá)質(zhì)粒pmCherry-ESAT-6為模板,PCR擴(kuò)增ESAT-6和mCherry融合片段,預(yù)期片段大小為1 107 bp。PCR擴(kuò)增產(chǎn)物經(jīng)過 1%的瓊脂糖凝膠電泳檢測,與預(yù)期片段大小基本一致(圖1)。
2.2 pTRE3G-mCherry-ESAT-6 重組質(zhì)粒的構(gòu)建雙酶切ESAT-6和mCherry融合片段后,使用DNA連接酶將其插入載體pLVX-TRE3G上,轉(zhuǎn)入大腸桿菌DH5α中。挑選23個(gè)單菌落克隆,經(jīng)菌落PCR驗(yàn)證獲得5個(gè)陽性克隆(圖2)。將陽性克隆擴(kuò)增,提取質(zhì)粒送往公司進(jìn)行測序驗(yàn)證,結(jié)果表明出入的基因片段與已公布的基因序列完全相同,這說明重組質(zhì)粒pTRE3G-mCherry-ESAT-6構(gòu)建成功。

M.DL2000 DNA Marker;1.ESAT-6基因片段圖1 mCherry-ESAT-6 基因的PCR擴(kuò)增
2.3 重組質(zhì)粒誘導(dǎo)表達(dá)ESAT-6的初步驗(yàn)證將質(zhì)粒pTRE3G-mCherry-ESAT-6和pLVX-TRE3G分別與調(diào)控質(zhì)粒pLVX-Tet3G按質(zhì)量比4∶1共轉(zhuǎn)染HEK293T細(xì)胞,并加入 100 μg/L的Dox誘導(dǎo)24 h后,熒光顯微鏡下觀察熒光表達(dá)情況,并收集細(xì)胞蛋白進(jìn)行 Western blot分析。結(jié)果表明,pTRE3G-mCherry-ESAT-6和 pLVX-Tet3G共轉(zhuǎn)時(shí),Dox可以誘導(dǎo)細(xì)胞發(fā)出紅色熒光,并且Western blot可檢測到ESAT-6的表達(dá)(圖3),這說明重組質(zhì)粒pTRE3G-mCherry-ESAT-6可用于后續(xù)的試驗(yàn)。
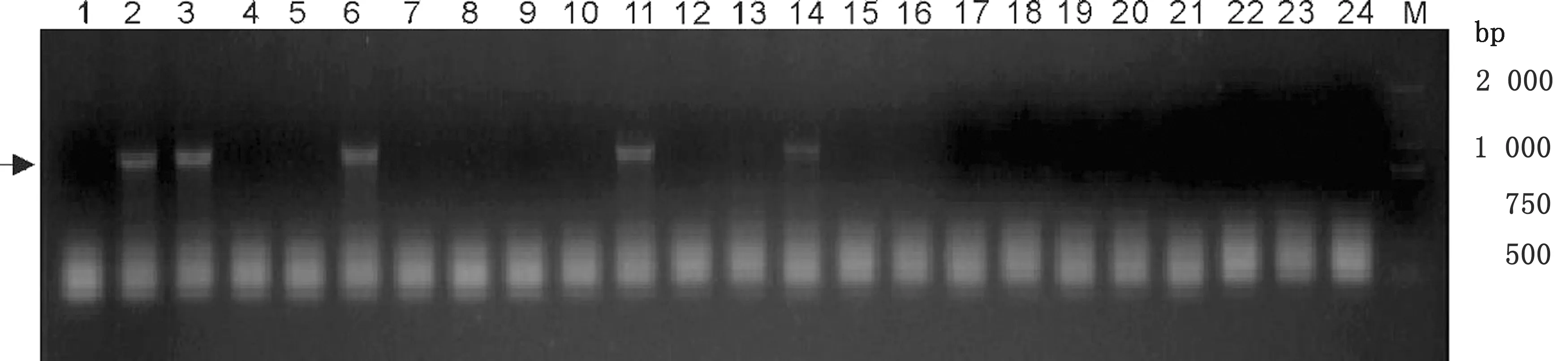
M.DL2000 DNA Marker;1.陰性對照;2~24.待驗(yàn)證的菌落圖2 菌落PCR鑒定

A.熒光顯微鏡觀察熒光表達(dá)情況;B.Western blot檢測ESAT-6蛋白表達(dá)情況圖3 重組質(zhì)粒在HEK293T細(xì)胞中誘導(dǎo)表達(dá)ESAT-6
2.4 可誘導(dǎo)表達(dá)的 THP-1/ESAT-6細(xì)胞系穩(wěn)定性篩選與鑒定用慢病毒LV-TRE3G-ESAT-6和LV-Tet3G按照一定的比例感染THP-1細(xì)胞,并用Puromycin和G418雙重壓力篩選獲得穩(wěn)定細(xì)胞THP-1/ESAT-6,對不同細(xì)胞代次用Dox進(jìn)行誘導(dǎo),提取細(xì)胞的總蛋白進(jìn)行Western blot分析。結(jié)果表明,篩選獲得的THP-1/ESAT-6細(xì)胞在 Dox的誘導(dǎo)下可檢測到ESAT-6的表達(dá),未誘導(dǎo)組未檢測到ESAT-6表達(dá)(圖4),這表明THP-1/ESAT-6細(xì)胞系構(gòu)建成功,且能夠穩(wěn)定表達(dá)ESAT-6。

圖4 不同細(xì)胞代次Dox誘導(dǎo)THP-1/ESAT-6細(xì)胞穩(wěn)定表達(dá)ESAT-6
2.5 ESAT-6表達(dá)與Dox質(zhì)量濃度和誘導(dǎo)時(shí)間的關(guān)系用不同質(zhì)量濃度的 Dox(0,100,200,400,600和800 μg/L)處理THP-1/ESAT-6細(xì)胞,在誘導(dǎo)24 h 后收集細(xì)胞蛋白,采用Western blot方法檢測 ESAT-6的表達(dá)。結(jié)果表明ESAT-6的表達(dá)隨著Dox質(zhì)量濃度的增加而增加,呈正相關(guān)性(圖5)。用100 μg/L的Dox處理細(xì)胞,分別在0,6,9,12,18,24,36,48,60和72 h后檢測 ESAT-6表達(dá)情況。結(jié)果表明Dox誘導(dǎo)9 h后可明顯檢測到ESAT-6,并且ESAT-6的表達(dá)具有時(shí)間依賴性(圖6)。

A.Western blot檢測不同質(zhì)量濃度的Dox誘導(dǎo)ESAT-6表達(dá)情況;B.灰度分析圖5 Dox質(zhì)量濃度對THP-1/ESAT-6細(xì)胞中ESAT-6表達(dá)的影響
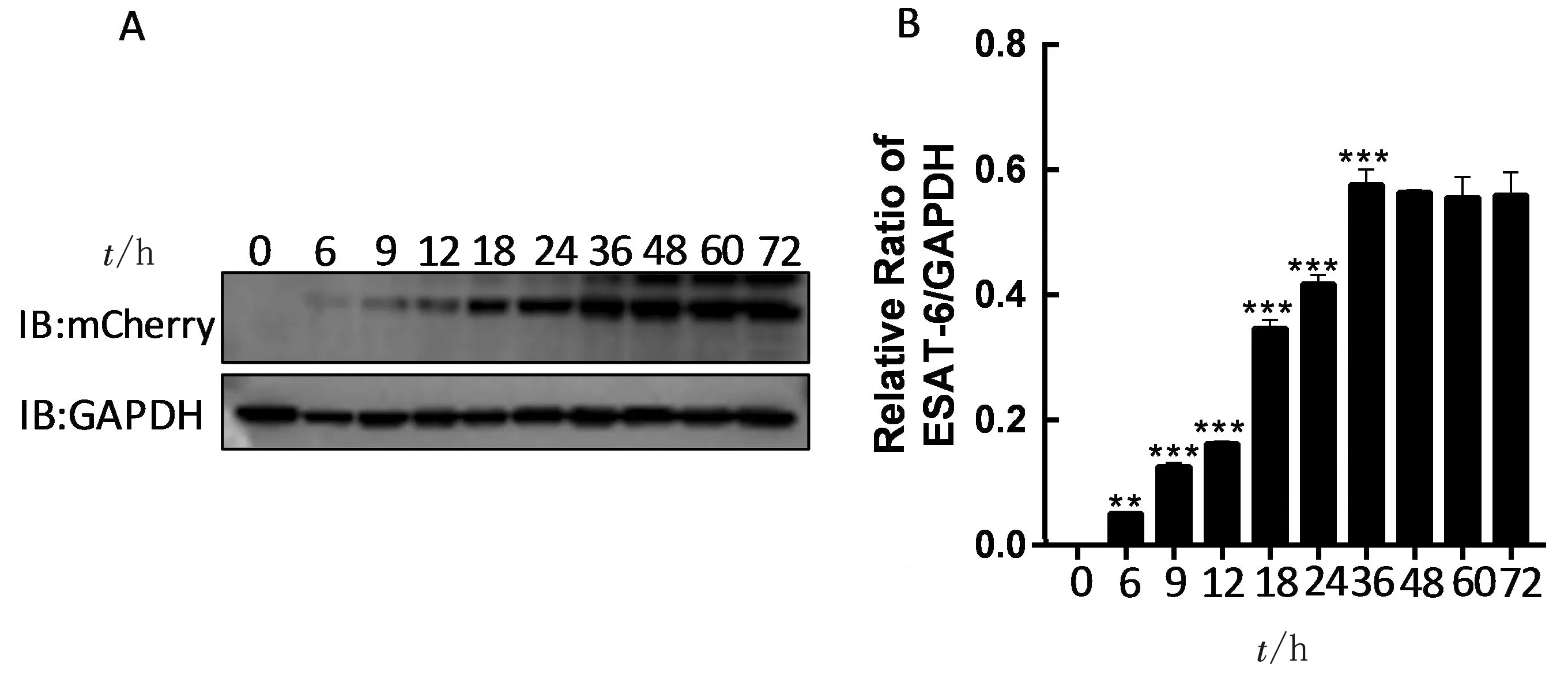
A.Western blot檢測Dox不同誘導(dǎo)時(shí)間下ESAT-6表達(dá)情況;B.灰度分析圖6 Dox誘導(dǎo)時(shí)間對THP-1/ESAT-6細(xì)胞中ESAT-6表達(dá)的影響
2.6 可誘導(dǎo)表達(dá)目的蛋白的生物活性檢測有研究發(fā)現(xiàn)MTB ESAT-6能夠引起NLRP3炎癥小體的激活介導(dǎo)IL-1β的分泌從而促進(jìn)細(xì)胞死亡[10]。為了驗(yàn)證誘導(dǎo)表達(dá)ESAT-6蛋白對細(xì)胞存活的影響,本試驗(yàn)用100 μg/L的Dox處理細(xì)胞,分別在0,6,9,12,18,24,36 和48 h后檢測細(xì)胞的存活率。結(jié)果表明Dox誘導(dǎo)18 h后細(xì)胞的存活率顯著下降(圖7A),說明隨著Dox誘導(dǎo)時(shí)間的增加ESAT-6的表達(dá)的增強(qiáng)可以促進(jìn)細(xì)胞的死亡。此外,為了驗(yàn)證誘導(dǎo)表達(dá)ESAT-6蛋白對NLRP3激活劑Nigericin(Nig)所引起的IL-1β分泌的影響,使用100 μg/L的Dox誘導(dǎo)劑培養(yǎng)THP-1/ESAT-6細(xì)胞20 h后,再加入1 mg/L脂多糖LPS刺激細(xì)胞3 h,10 μmol/L尼日利亞菌素Nigericin作用1 h,通過ELISA檢測細(xì)胞上清IL-1β的變化。結(jié)果表明ESAT-6能夠促進(jìn)由尼日利亞菌素Nigericin引起的IL-1β的分泌(圖7B)。這些結(jié)果表明誘導(dǎo)表達(dá)蛋白ESAT-6具有促進(jìn)IL-1β的分泌產(chǎn)生和細(xì)胞死亡的生物活性。
3 討論
ESAT-6是MTB的免疫優(yōu)勢抗原和主要的毒力因子,參與調(diào)控宿主細(xì)胞的多種固有免疫反應(yīng)。有研究表明MTB ESAT-6能夠介導(dǎo)NLRP3炎癥小體的激活,觸發(fā)下游因子Caspase-1的活化,從而促進(jìn)IL-1β的分泌和細(xì)胞焦亡的發(fā)生[10-11]。ESAT-6還能夠觸發(fā)I型干擾素的產(chǎn)生,這主要是通過激活STING-TBK1-IRF3信號轉(zhuǎn)導(dǎo)軸特異性激活配體導(dǎo)致的[5]。目前,對ESAT-6功能的研究主要通過蛋白純化表達(dá)和缺失菌株的構(gòu)建來完成。通過大腸桿菌原核表達(dá)所獲得的ESAT-6蛋白中往往含有大腸桿菌本身的菌體物質(zhì),如脂多糖(LPS),或者含有在純化過程中添加的去污劑。然而,這些物質(zhì)的存在有時(shí)會造成試驗(yàn)結(jié)果出現(xiàn)誤差。多項(xiàng)研究報(bào)道發(fā)現(xiàn)ESAT-6以酸性pH依賴性方式誘導(dǎo)脂質(zhì)泄露,具有膜溶解的功能[12-13]。但是,CONRA等[14]發(fā)現(xiàn)ESAT-6本身不具備溶解細(xì)胞膜的功能,之前的研究發(fā)現(xiàn)ESAT-6之所以能夠溶劑細(xì)胞膜,完全歸因于ESAT-6中殘留的去污劑ASB-14。因此,本試驗(yàn)通過Tet-On 3G系統(tǒng)構(gòu)建可誘導(dǎo)表達(dá)ESAT-6的細(xì)胞系,避免大腸桿菌菌體物質(zhì)和去污劑等造成的影響。

A.ELISA檢測Dox對LPS+Nig引起THP-1/ESAT-6細(xì)胞中IL-1β分泌的影響;B.Dox不同誘導(dǎo)時(shí)間對THP-1/ESAT-6細(xì)胞存活的影響圖7 可誘導(dǎo)表達(dá)ESAT-6蛋白的生物活性鑒定
Tet-On 3G系統(tǒng)對Dox具有相當(dāng)高的特異性和敏感度,且Tet毒性低,不影響細(xì)胞內(nèi)的其他基因活性。并且,Tet-On 3G系統(tǒng)誘導(dǎo)基因表達(dá)的效果與Dox的濃度及誘導(dǎo)時(shí)間存在一定的關(guān)系,可以自由控制靶基因表達(dá)的時(shí)間和強(qiáng)度[15]。鑒于Tet-On 3G系統(tǒng)的上述特性,本試驗(yàn)中利用該系統(tǒng)構(gòu)建了可誘導(dǎo)表達(dá)ESAT-6蛋白的THP-1細(xì)胞。當(dāng)未加入Dox的條件下,THP-1細(xì)胞不表達(dá)ESAT-6;當(dāng)加入Dox時(shí),THP-1細(xì)胞可誘導(dǎo)表達(dá)ESAT-6,并且100 μg/L 的Dox誘導(dǎo)6 h即可觀察到ESAT-6的表達(dá),誘導(dǎo)9 h后ESAT-6表達(dá)量明顯上升,在36 h達(dá)到最大值,此后表達(dá)量保持穩(wěn)定。
THP-1分化的巨噬細(xì)胞是研究MTB的主要體外細(xì)胞模型。THP-1細(xì)胞是典型的懸浮細(xì)胞,其轉(zhuǎn)染并瞬時(shí)表達(dá)外源蛋白相對比較困難。Lipofectamine 2000對THP-1毒性較大,轉(zhuǎn)染效率極低,不適合THP-1細(xì)胞轉(zhuǎn)染。腺病毒對THP-1細(xì)胞毒性較低,并且具有極高的轉(zhuǎn)染效率,但腺病毒免疫原性較高,高M(jìn)OI極大影響THP-1細(xì)胞免疫狀態(tài),有影響后續(xù)試驗(yàn)的風(fēng)險(xiǎn)。相比腺病毒來說,慢病毒同樣對THP-1的毒性較低,但其轉(zhuǎn)染效率有所降低[16]。不過,它具有較低的免疫原性,對細(xì)胞免疫狀態(tài)的影響小。考慮3種轉(zhuǎn)染方式的優(yōu)缺點(diǎn),本試驗(yàn)包裝了LV-TRE3G-ESAT-6和LV-Tet3G這2種慢病毒用以感染THP-1細(xì)胞。
綜上,本試驗(yàn)首次采用Tet-On 3G誘導(dǎo)表達(dá)系統(tǒng)成功構(gòu)建了由Dox穩(wěn)定誘導(dǎo)表達(dá)ESAT-6的THP-1細(xì)胞系,分析了不同誘導(dǎo)濃度和誘導(dǎo)時(shí)間對ESAT-6表達(dá)量的影響以及ESAT-6的生物活性,為探究ESAT-6功能提供了新的試驗(yàn)方法,也為深入探究ESAT-6參與MTB調(diào)控細(xì)胞通路的作用機(jī)制奠定了基礎(chǔ)。
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
