浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚的生長、性成熟和死亡特征研究
2021-12-22 05:56:52裴如德麻秋云田思泉張云飛范青松
南方水產科學 2021年6期
裴如德,麻秋云,田思泉,張云飛,范青松
(1. 上海海洋大學海洋科學學院,上海 201306; 2. 上海科技館自然史研究中心,上海 200041;3. 浙江省海洋水產養殖研究所,浙江溫州 325000)
鱗鰭叫姑魚 (Johnius distinctus) 和皮氏叫姑魚(J. belangerii) 同為石首魚科、叫姑魚屬常見的小型經濟魚類,兩者生活史特征相似,個體大小較為接近 (體長200 mm以下),是生活在泥沙底質的中下層暖水性魚類,以底棲生物為主要食物來源,夏季為其主要產卵繁殖期;捕撈方式以底拖網為主,春、夏季為其主要捕撈季節[1-5]。
目前國內外關于這2種魚類生態學和生活史特征的研究較少,主要集中在群落結構中的重要性指數[6-9]、資源密度[10-12]以及食性分析[13]、耳石形態[14]、環境污染對其生活史的影響[15-16]等方面。近年來對皮氏叫姑魚的基礎生物學研究較多,如馬鞍列島皮氏叫姑魚體長分布、雌雄比例、性腺成熟度隨月份的變化[17],流沙灣皮氏叫姑魚肥滿度隨月份的變化[18],廈門海域皮氏叫姑魚優勢體長組的季節變化、雌雄比例與體長的關系、繁殖特性及生長死亡參數[19]等。然而,對鱗鰭叫姑魚的研究匱乏,僅見部分學者對臺灣省西南海域、閩江口、北部灣鱗鰭叫姑魚體長-體質量關系的研究[20-22]。
皮氏叫姑魚和鱗鰭叫姑魚是我國東海、南海等多個海域生物群落中的重要魚類,也是生態系統中的餌料生物[6-9,23-29],在生態學研究上具有重要意義。近年來,由于捕撈強度加大、環境污染和氣候變化等因素的綜合影響,浙江南部近海等海域傳統經濟性魚類資源出現顯著衰退,皮氏叫姑魚和鱗鰭叫姑魚等小型魚類在生態系統中的重要性上升,在漁業中的占比逐漸增加[17,30]。目前我國大力推進海洋經濟種類的科學管理,研究分析這2種叫姑魚的生物學特征對理解東海生態系統和促進其漁業科學管理有重要意義。本文研究了浙江南部近海皮氏叫姑魚和鱗鰭叫姑魚的生長、年齡、性成熟和死亡等生物學特征,以期為浙江南部近海漁業資源的合理開發和科學管理提供基礎參考資料。
1 材料與方法
1.1 數據來源
2種叫姑魚的生物學樣品于2015—2018年采自浙江南部溫州、臺州近海水域(圖1),各季度調查中每個站點隨機選取30尾留樣,不足30尾的全取。在實驗室對采集的樣品按照《海洋調查規范》(海洋生物調查GB/T 12763.6—2007) 進行生物學測定,測定內容包括體長、體質量、性別、性腺發育程度等,其中體長精確到1 mm,體質量精確到0.1 g。13個航次調查共采集鱗鰭叫姑魚1 020尾,皮氏叫姑魚874尾 (表1)。
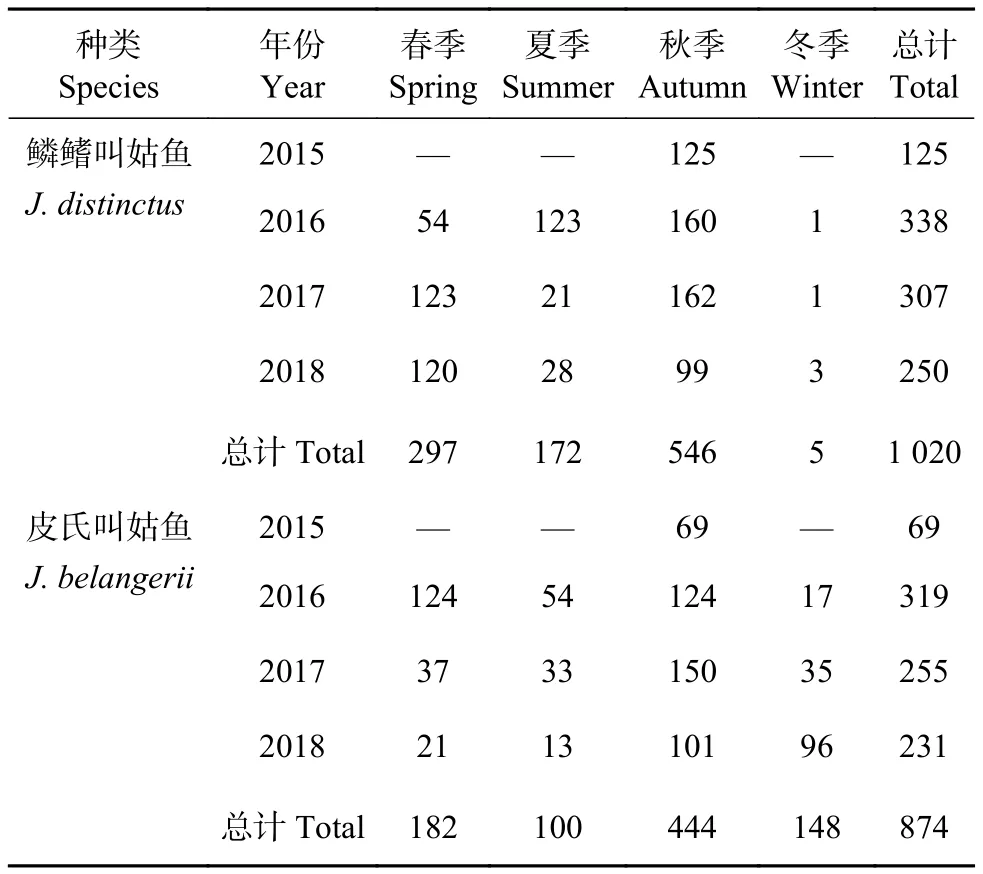
表1 2015—2018年浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚的樣本數量Table 1 Sample size of J. distinctus and J. belangerii in offshore waters of southern Zhejiang Province from 2015 to 2018

圖1 2015—2018年浙江南部近海漁業資源調查站位圖Figure 1 Sampling stations of fishery resources survey in offshore waters of southern Zhejiang Province from 2015 to 2018
1.2 體長和體質量關系
鱗鰭叫姑魚和皮氏叫姑魚的體長和體質量關系用冪指數公式[31]描述:

對數化后為:
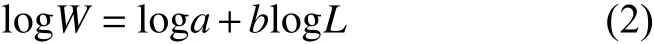
式中:W為體質量;L為體長;a為生長條件因子;b為異速生長因子。為探討體長和體質量關系的年份、季節和性別異質性,本文使用廣義線性模型 (Generalized linear model, GLM) 和21個線性混合效應模型 (Linear mixed effects model, LMEM)分別研究鱗鰭叫姑魚和皮氏叫姑魚的體長-體質量關系。21個LMEM模型在截距loga和斜率b上分別加入季節、年份和性別隨機效應 (表2)。因鱗鰭叫姑魚冬季樣本數量較少 (僅5尾),不具有代表性,因此分析季節異質性時,只有春、夏、秋3季。赤池信息量準則 (Akaike information criterion, AIC )和均方根誤差 (Root mean square error, RMSE) 最小的模型為最優模型。
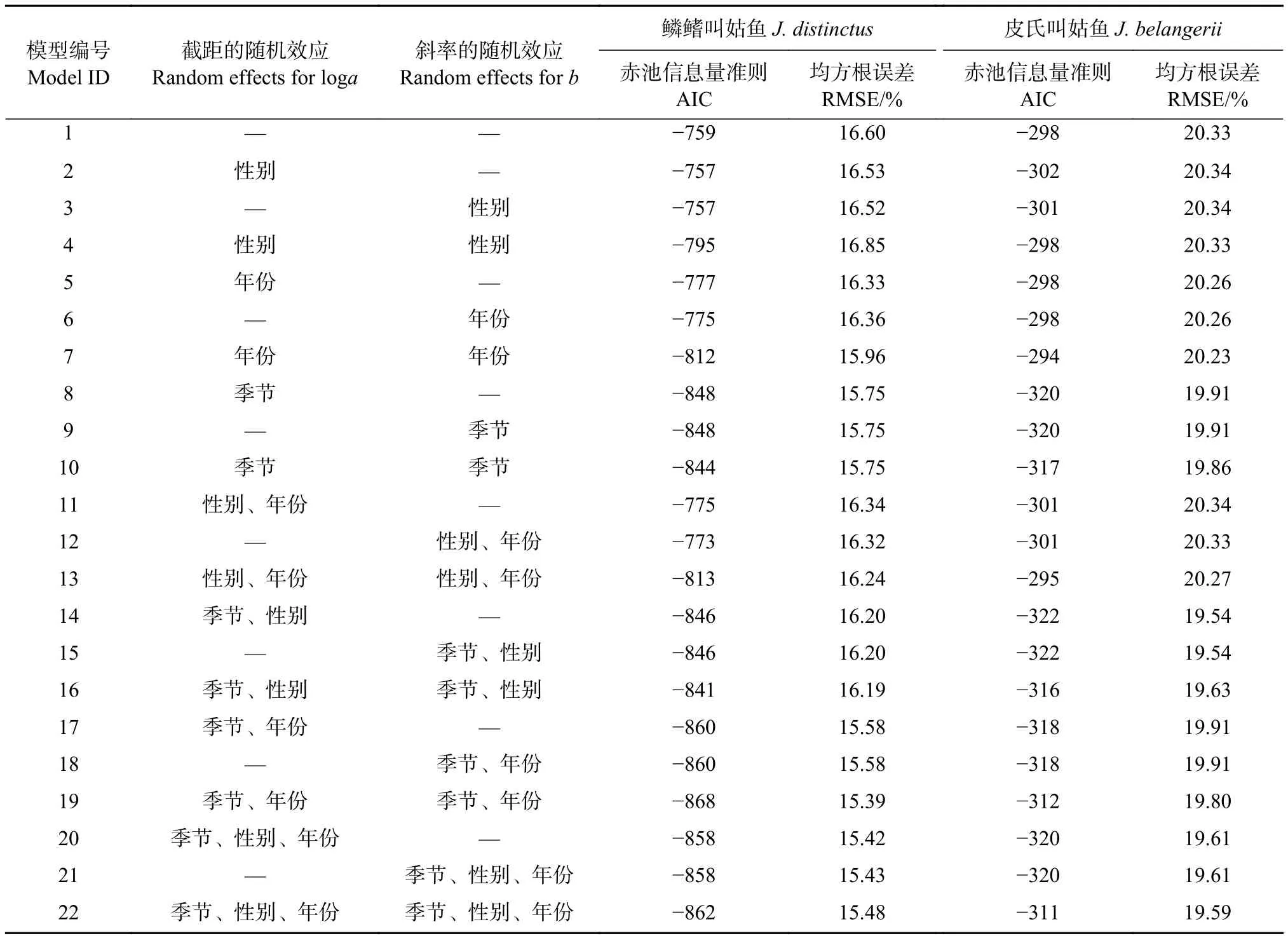
表2 浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚體長體質量關系的擬合模型及其評價指標Table 2 Candidate models to describe length-mass relationship for J. distinctus and J. belangerii in offshore waters of southern Zhejiang Province and their AIC and RMSE values
1.3 年齡和體長關系
使用von Bertalanffy生長方程[32]分別描述鱗鰭叫姑魚與皮氏叫姑魚的年齡-體長關系:
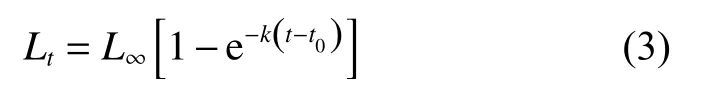
式中:L∞為極限體長;t0為理論生長起始年齡;k為趨近極限體長的相對速率。分別按10 mm體長分組,采用FiSAT II 軟件中的ELEFAN I方法[33]進行體長頻率分析,估算L∞和k。使用Pauly經驗公式[34]估算t0:
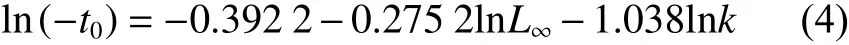
1.4 性成熟特征
分別將鱗鰭叫姑魚和皮氏叫姑魚繁殖期性腺成熟度Ⅲ期及以上個體視為性成熟個體,采用Logistic曲線[35],擬合性成熟概率與體長關系:
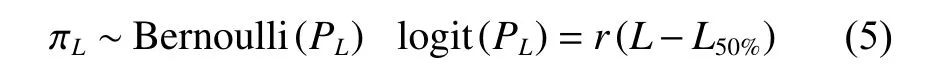
式中:π為個體性成熟與否;Bernoulli為伯努利分布;P為性成熟概率;r為瞬時性成熟速度;L50%為初次性成熟體長 (50%性成熟體長)。
1.5 死亡特征
使用FiSAT II 軟件中的長度變換漁獲曲線法,分別估算鱗鰭叫姑魚和皮氏叫姑魚的總死亡系數(Z)。使用Pauly經驗公式[34]估算自然死亡系數 (M):
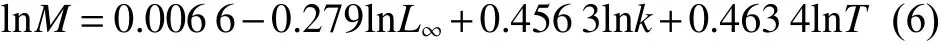
式中:T為魚類棲息海域年平均溫度;根據調查海域表層溫度數據,對鱗鰭叫姑魚和皮氏叫姑魚漁獲物加權求平均溫度,分別為21.1和19.7 ℃。根據式 (7) 和 (8) 計算捕撈死亡系數 (F) 和開發率 (E):

2 結果
2.1 體長體質量組成
浙江南部近海鱗鰭叫姑魚樣品體長介于32.9~209.0 mm,平均為 (115.7±27.1) mm,優勢體長組為88.6~142.8 mm,占樣本總數的72.5% (圖2-a)。皮氏叫姑魚樣品體長介于43.0~202.0 mm,平均為(136.3±36.5) mm,優勢體長組為99.9~172.8 mm,占樣本總數的67.0% (圖2-b)。鱗鰭叫姑魚體質量介于0.4~188.2 g,平均為 (37.8±24.7) g,優勢體質量組為13.2~62.6 g,占樣本總數的82.8% (圖2-c);皮氏叫姑魚體質量介于1.5~183.9 g,平均為 (63.8±39.3) g,優勢體質量組為24.5~103.1 g,占樣本總數的 59.4% (圖 2-d)。
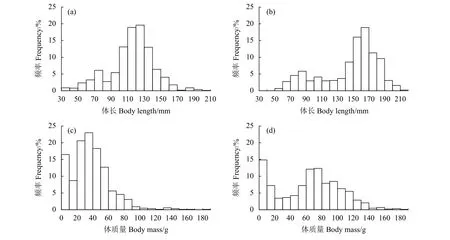
圖2 2015—2018年浙江南部近海鱗鰭叫姑魚 (a、c) 和皮氏叫姑魚 (b、d) 的體長與體質量分布Figure 2 Distribution of body length and body mass of J. distinctus (a, c) and J. belangerii (b, d) in offshore waters of southern Zhejiang Province from 2015 to 2018
2.2 體長與體質量關系
浙江南部近海鱗鰭叫姑魚的體長與體質量關系為W=4.02×10-6L3.34(R2=0.98)。模型中,截距loga和斜率b上同時加入季節和年份隨機效應的LMEM模型AIC和RMSE最小,擬合效果最優(表2,模型19)。該模型中,a和b的固定效應分別為8.57×10-6和3.18,加入隨機效應后a和b分別介于 1.51×10-6~4.09×10-6和 3.06~3.33。該模型結果顯示,在浙江南部近海鱗鰭叫姑魚體長-體質量關系中,季節影響最顯著,年份次之,性別無顯著影響。在體長-體質量關系曲線中,體長大于150.0 mm后,相同體長情況下,鱗鰭叫姑魚的增重率春季最快,秋季次之,夏季最慢;2016年最快,2015年最慢 (圖 3-a1、3-a2)。
皮氏叫姑魚體長與體質量關系為W=5.63×10-6L3.26(R2=0.98)。模型中,截距loga加入季節和性別隨機效應的LMEM模型AIC和RMSE最小,擬合效果最優 (表2,模型14)。該模型中,a和b的固定效應值分別為6.65×10-6和3.22,加入隨機效應后a介于 6.19×10-6~6.91×10-6。該模型結果顯示,在浙江南部近海皮氏叫姑魚體長-體質量關系中,季節影響最顯著,性別影響較小。在體長-體質量關系曲線中,相同體長情況下,皮氏叫姑魚的增重率春季最快,其次為秋季和夏季,冬季最慢,雌性的增重率快于雄性 (圖3-b1、圖3-b2)。總體上,在相同體長下,鱗鰭叫姑魚的增重率快于皮氏叫姑魚 (圖 4)。
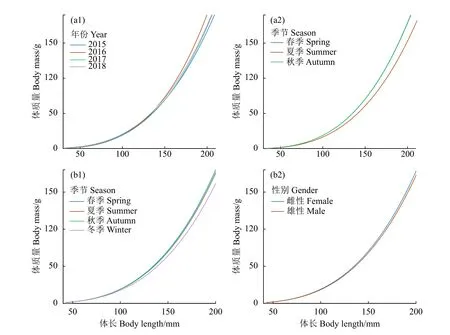
圖3 浙江南部近海鱗鰭叫姑魚 (a) 和皮氏叫姑魚 (b) 的體長-體質量關系異質性Figure 3 Variations of length-mass relationship of J. distinctus (a) and J. belangerii (b) in offshore waters of southern Zhejiang Province
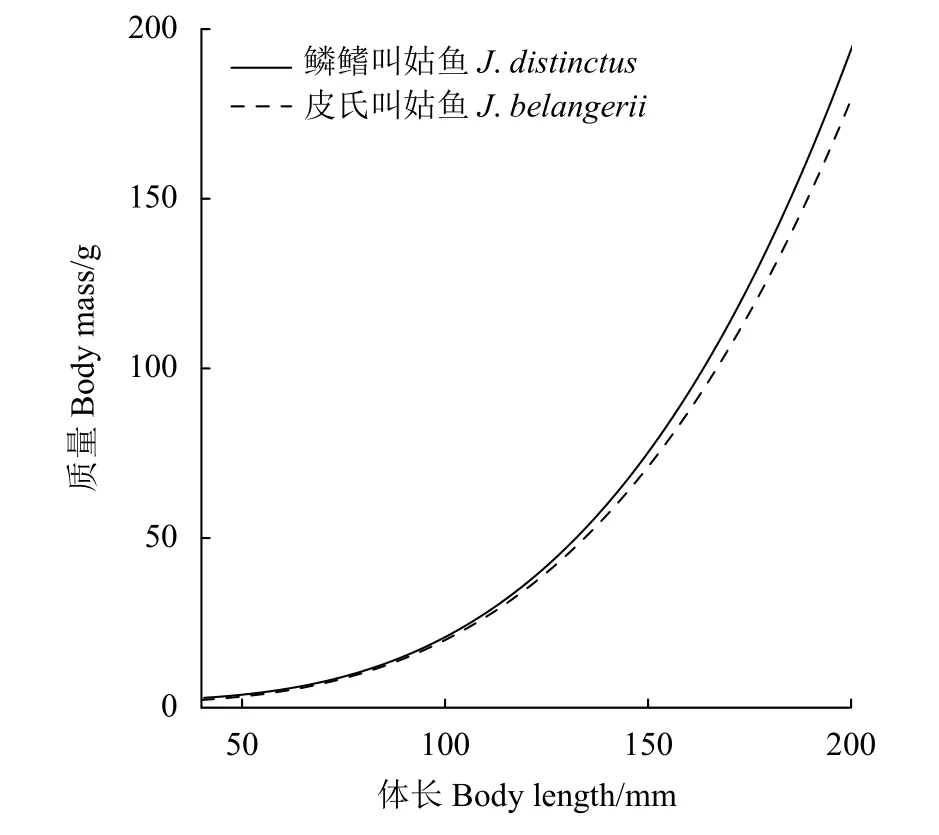
圖4 2015—2018年浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚的體長-體質量關系Figure 4 Length-mass relationship of J. distinctus and J. belangerii in offshore waters of southern Zhejiang Province from 2015 to 2018
2.3 生長參數
估算得出鱗鰭叫姑魚的L∞=216.83 mm,k=0.84;皮氏叫姑魚的L∞=207.38 mm,k=0.63。由Pauly經驗公式估算理論生長起始年齡,鱗鰭叫姑魚t0=-0.18,皮氏叫姑魚t0=-0.25。根據求得的生長參數,繪制了2種叫姑魚的von Bertalanffy生長曲線 (圖 5)。
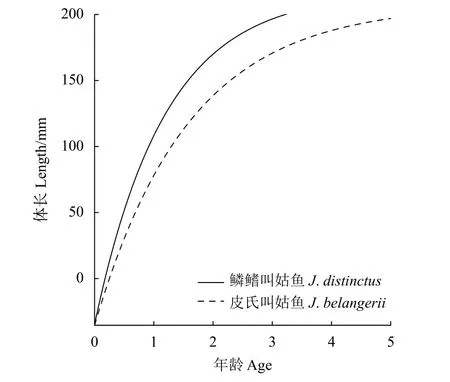
圖5 2015—2018年浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚生長曲線Figure 5 Growth curve of J. distinctus and J. belangerii in offshore waters of southern Zhejiang Province from 2015 to 2018
2.4 性腺與性成熟特征
864尾鱗鰭叫姑魚有性腺數據,雄雌比為1∶0.38,性腺成熟度Ⅱ期的占53.4%,Ⅲ期及以上的僅24.2%。782尾皮氏叫姑魚有性腺數據,雄雌比為1∶0.91,性腺成熟度為Ⅱ期的占47.4%,Ⅲ期及以上的僅占39.1% (圖6)。鱗鰭叫姑魚和皮氏叫姑魚初次性成熟體長分別為133.1和128.9 mm,兩者體長和性成熟概率關系 (圖7) 為:
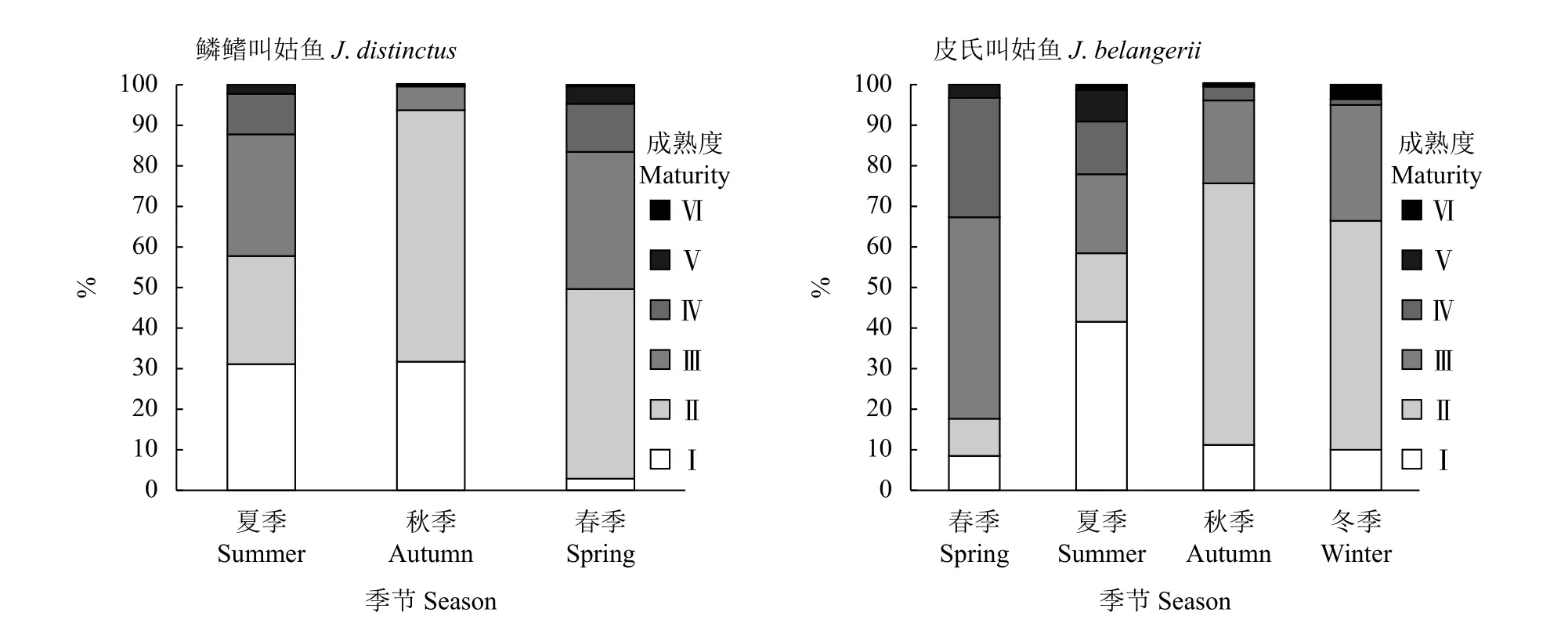
圖6 2015—2018年浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚性腺成熟度的季節組成Figure 6 Seasonal variation of maturity of J. distinctus and J. belangerii in offshore waters of southern Zhejiang Province from 2015 to 2018
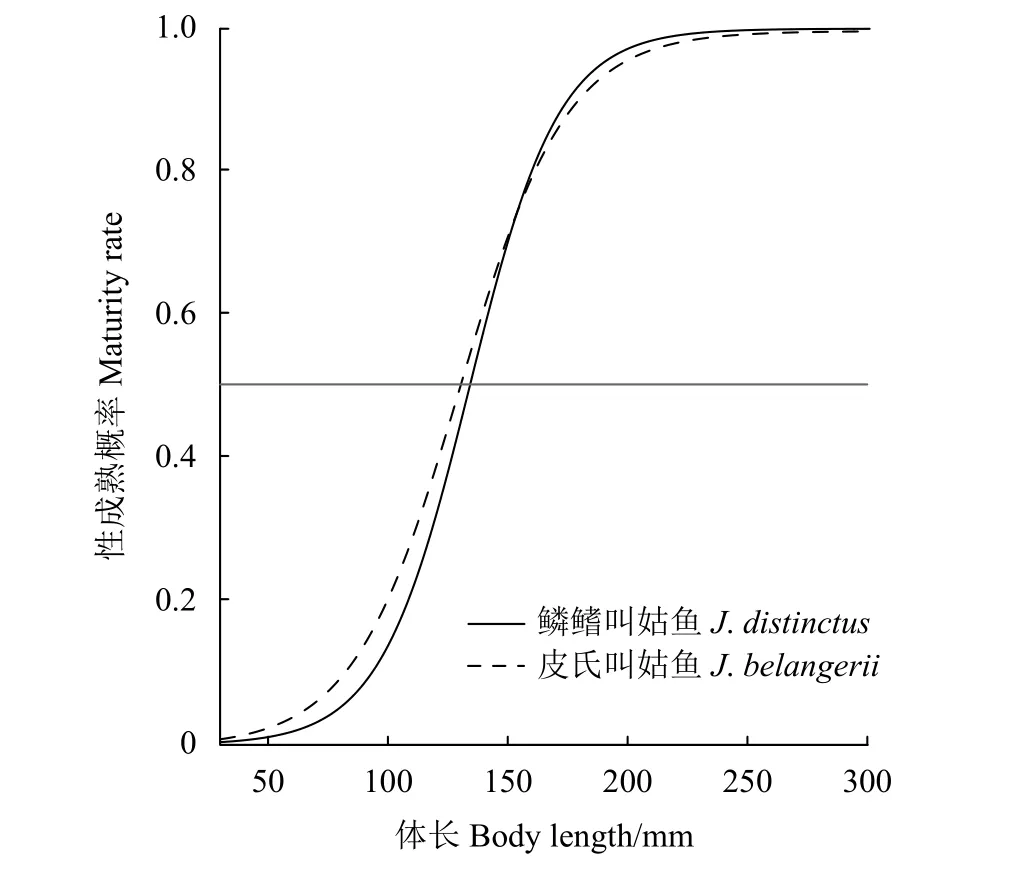
圖7 2015—2018年浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚體長和性成熟概率關系灰色橫線為50%的性成熟概率。Figure 7 Relationship between length and maturity rate of J. distinctus and J. belangerii in offshore waters of southern Zhejiang Province from 2015 to 2018 The grey horizonal line represents the 50% maturity rate.
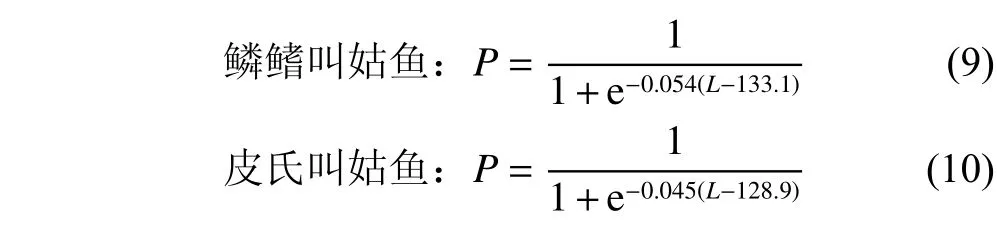
2.5 死亡特征
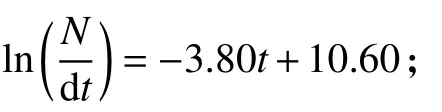
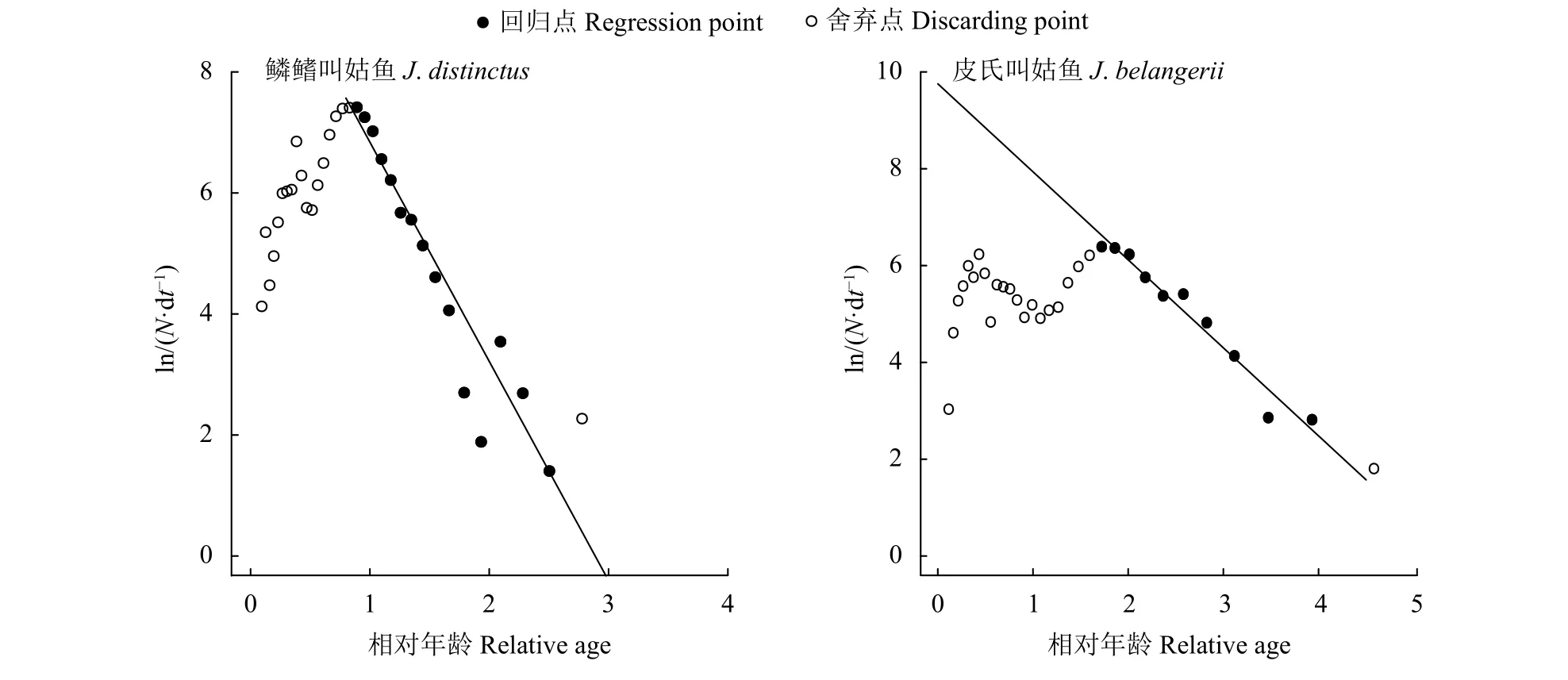
圖8 根據體長變換漁獲曲線估算鱗鰭叫姑魚和皮氏叫姑魚總死亡系數Figure 8 Estimation of total morality based on length-converted catch curve of J. distinctus and J. belangerii
3 討論
3.1 體長-體質量關系分析
本研究通過比較2種叫姑魚的各項生物學特征,發現棲息地環境和繁殖模式相似的鱗鰭叫姑魚與皮氏叫姑魚的體質量差異較大,相同體長條件下,鱗鰭叫姑魚體質量更大。在魚類體長-體質量關系方程中,生長條件因子a和異速生長因子b共同決定了魚類的生長模式。其中a與魚類餌料基礎、棲息地水文條件等環境因素有關[32],a越大,說明該水域生活環境條件越利于魚類生長。
本研究通過線性混合效應模型得到的皮氏叫姑魚 a 介于 6.19×10-6~6.91×10-6,相對其他區域的研究結果較小 (表3),這可能由環境因素影響所致。馬鞍列島、閩江口、廈門海域、珠江口、北部灣等處于島礁區,有大型河流入海、影響較大的河口、灣區域及低緯度區域,水文條件、餌料生物數量等生長棲息環境均優于浙江南部近海[6,36]。b一般介于2.5~3.5,可以判斷魚類是否為等速生長[37]。本研究得到的皮氏叫姑魚b為3.22,說明浙江南部近海皮氏叫姑魚為正異速生長,除了珠江口與臺灣西南部外,與其他研究結果一致,數值和福建沿岸較為接近。種群差異可能是導致不同海域皮式叫姑魚生長模式出現差異的原因[17],浙江南部近海皮氏叫姑魚和福建沿岸皮氏叫姑魚同屬東海南部種群,b較為接近。對鱗鰭叫姑魚生物學、體長-體質量關系的研究相對較少,且各研究結果差異較大,本研究得到的a、b與臺灣西南部[20](W=3.95×10-5L2.88)、閩江口[21](W=1.33×10-5L3.1)、北部灣[22](W=1.75×10-5L3.03)均不同,相關的地域差異還需要更多的研究和分析。

表3 不同海域皮氏叫姑魚生長、死亡參數Table 3 Growth and mortality parameters of J. belangerii in different China's seas
以往對鱗鰭叫姑魚和皮氏叫姑魚體長-體質量關系的研究側重于整體分析,忽略了其生長過程中的時空差異,本研究通過線性混合效應模型發現2種叫姑魚體長-體質量關系有顯著的季節差異,增重率均在春季最高。皮氏叫姑魚產卵期為4—7月[5,23],本研究春季航次為5月,此時性腺成熟度較高,增重率最高;秋季為兩者索餌洄游期,攝食利用率較高;夏季為產卵后期,群體中有大量幼體補充,整體生長速度較快,增重率也較高。而皮氏叫姑魚越冬洄游階段,攝食較少,增重率較低。為全面了解鱗鰭叫姑魚和皮氏叫姑魚生長特征的時空差異,今后應從生長空間差異性、幼魚成魚差異性入手,進一步分析研究這2種叫姑魚體長-體質量關系。
3.2 生長死亡參數分析
魚類生長、死亡參數是分析其群體資源動態的重要指標。通過比較發現,浙江南部近海與其他海域皮氏叫姑魚的極限體長、生長速率、理論生長起始年齡、總死亡系數等參數有一定差異 (表3),極限體長與廈門灣接近,總死亡系數介于廈門灣和廈門海域、福建沿岸之間,自然死亡系數大于其他海域,顯示了不同海域棲息地環境和種群結構的差異性。捕撈死亡系數和開發率反映了各海域皮氏叫姑魚捕撈強度的差異,總體來看,廈門灣、福建沿岸和廈門海域開發強度接近,而浙江南部近海開發強度最低。Gullan[39]提出開發率為0.5是漁業資源群體開發的臨界點,本研究中皮氏叫姑魚開發率E低于該臨界點,即資源未過度開發,因此該漁業目前可在實施養護和管理的基礎上適度開發。
3.3 性成熟特征分析
浙江南部近海皮氏叫姑魚雄雌比 (1∶0.91) 與廈門海域[19](1∶0.87) 接近,與馬鞍列島[17](0.59∶1)、膠州灣[40](1∶0.7) 的差異較大,這需要結合營養條件、生長、年齡、季節、捕撈方式等進一步研究。50%性成熟體長值在漁業管理中有著廣泛應用,是確定開捕體長的重要生物學參考點[41-43],本研究中由繁殖期皮氏叫姑魚樣本求得的50%性成熟體長為128.9 mm,接近于馬鞍列島[17](雌性124.8 mm,雄性136.4 mm)。因本研究未考慮性腺成熟度、性比的季節和體長組的差異,今后需從不同海域環境條件、繁殖特征等進行更多相關研究。
對不同海域皮氏叫姑魚與鱗鰭叫姑魚的生活史特征參數進行直接對比,會受樣本數量、采樣時間跨度、站點數量、樣本捕撈獲取方式等因素的影響。如臺灣西南海域鱗鰭叫姑魚樣本數量為151尾,本研究為1 020尾;皮氏叫姑魚流沙灣樣本數為541尾,本研究為874尾;采樣時間跨度馬鞍列島皮氏叫姑魚為1年月度調查,廈門海域和福建沿岸為2年4個季度調查航次,北部灣為2年8個季度調查航次,本研究為4年13個季度調查航次;廈門海域皮氏叫姑魚設7個調查站位,福建沿岸為24個,本研究為42個。因此,相較而言,本研究的時空跨度更長,樣本采集更多,有助于獲得準確性和精確性更高的研究結果。此外,馬鞍列島皮氏叫姑魚樣本用底拖網和刺網獲取,珠江口用尖尾罟網,本研究用底拖網,不同網具的選擇性差異可能導致樣本對種群結構的代表性不同。
4 結論
本文分析了浙江南部近海鱗鰭叫姑魚和皮氏叫姑魚的生長、繁殖和死亡等生物學特征。總體上,2種叫姑魚的生物學特征既有相似性又有差異性。2種魚的體長-體質量關系均有顯著季節差異,春季增重率最高,均為異速生長魚類,性腺成熟度組成均以Ⅱ期為主;相同體長下,鱗鰭叫姑魚體質量更大;相同年齡下,鱗鰭叫姑魚體長更大,生長速度也更快;鱗鰭叫姑魚初次性成熟的體長更大,自然死亡系數更高,開發率高于皮氏叫姑魚。本研究結果為該海域2種魚類的生物學研究提供了基礎資料,并可為其資源評估和漁業可持續開發利用提供參考依據。
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
非公有制企業黨建(2020年9期)2020-09-26 13:22:18
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
文苑(2018年22期)2018-11-19 02:54:14
領導決策信息(2017年12期)2017-05-17 04:49:18
中國衛生(2016年7期)2016-11-13 01:06:44
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
