富齒巖蟲南移養殖與營養成分分析
2021-12-22 05:57:00李帥鵬黃民巧楊大佐周一兵
南方水產科學 2021年6期
關鍵詞:生長
李帥鵬,徐 斌,王 坤,黃民巧,楊大佐,周一兵
(1. 大連海洋大學/遼寧省海洋生物資源恢復與生境修復重點實驗室,遼寧 大連 116023;2. 廣東海興農集團有限公司,廣東 廣州 511400)
巖蟲 (Marphysa sp.) 屬于環節動物門、游走目、磯沙蠶科,是一類具有重要經濟和生態價值的大型底棲多毛類動物。由于其全球性分布、物種多樣性以及分類爭議等原因,Fauchald根據主要形態差異將巖蟲屬分為4個組,分別為Mossambica、Sanguinea、Aeana和Falcigers[1]。其中Sanguinea組常被稱為巖蟲復合系 (Marphysa. Sanguine complex)。同時,以巖蟲復合系為對象的基礎研究廣泛展開,在系統分類[2-4]、分子鑒定[5]、生活史[6-7]、人工繁育[8]、生態學[9-10]、基因組學[11-13]、能量代謝[14-15]以及海洋環境污染與修復[16-17]等方面均有報道。Liu等[18]對中國近海不同地區采集的巖蟲復合系重新開展了系統分類學研究,將山東乳山等地的原鑒定為巖蟲 (Marphysa sanguinea) 的物種重新分類,并確定為新的物種富齒巖蟲 (M. maxidenticulata)。以該研究為基礎,結合分子鑒定特征,確定山東乳山近海廣泛分布的大型經濟磯沙蠶為富齒巖蟲。
富齒巖蟲主要生活在山東、遼寧與河北等北方地區的潮間帶富含巖礁石的泥沙底質內,由于其攝食沉積質表層的顆粒性有機物,并通過穴居挖洞和生物擾動作用改善沉積質內氧氣流通,因此具有重要的生態學功能。在海水養殖業中,多毛類因豐富的營養組成,特別是含有豐富的二十二碳五烯酸(DPA) 和花生五烯酸 (EPA) 等多不飽和脂肪酸,能夠顯著促進對蝦的性腺發育和成熟,在國內外常作為經濟性對蝦親體促熟培育的優質活體餌料[19-21]。此外,富齒巖蟲在海洋游釣產業用作優質活體釣餌,其售價高達每千克500元,顯示出較高的經濟價值。近年來由于過度采捕,北方地區富齒巖蟲的資源數量顯著下降,亟須通過人工繁育和養殖等方式,促進資源的可持續利用。然而,由于北方的氣候條件和磯沙蠶科的生長特性,存在生長速度慢、生產周期長等問題,影響了資源的利用效率。參考我國廣泛開展的北種南養生產模式,本文開展了富齒巖蟲在廣西北海的工廠化人工養殖試驗,鑒于其營養構成在水產蝦蟹親體促熟中的重要性,比較和分析了南方養殖與北方野生的富齒巖蟲營養組成,以期為富齒巖蟲的人工養殖、資源開發與可持續利用提供參考。
1 材料與方法
1.1 材料
富齒巖蟲苗種來源于山東省乳山市,為人工育苗后經中間育成的大規格苗種。苗種長度為(1.00±0.20) cm,體質量為 (0.03±0.01) g,數量總計達12萬尾。苗種經人工篩網收集后,低溫保存,干法運輸,航空運輸至廣西北海市合浦縣。
1.2 方法
1.2.1 養殖試驗 試驗地點位于廣西壯族自治區北海市廣東海興農集團的合浦縣養殖場。試驗所選室外養殖水泥淺池6個,其規格為長6 m×寬4 m×高0.45 m。經石灰消毒、清洗曝曬后,分別置入北海本地購置的海泥 (記為A1、A2、A3) 和粗砂 (記為B1、B2、B3) 2種底質。其中粗砂粒徑為 2~3 mm,6個養殖池底質厚度均為15 cm。然后用0.15 mg·L-1高錳酸鉀進行消毒,再用 0.2 mg·L-1草酸溶液去除高錳酸鉀殘余,經曝曬及海水多次沖洗后待用。將苗種平均投放各池,養殖密度為833條·m-2,采用帶水充氣法培育,每日模擬潮汐換水1次;每日定時測量水溫和鹽度并進行記錄,養殖過程全程投喂凡納濱對蝦配合飼料 (容川牌6210幼蝦0號料),日投喂量為池內蟲體總體質量的5%~8%。
1.2.2 生長指標測定 采用定期取樣法,每隔60 d從每個養殖池隨機挖50條蟲體,經過濾海水沖洗干凈、麻醉后用紗布和濾紙吸取體表水分,利用直尺測定其體長、體寬,用天平測定其體質量。根據測定結果,計算其特定生長率 (SGR)
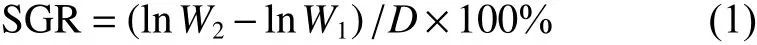
式中:W1為初始體質量 (g);W2為終末體質量 (g);D為養殖時間 (d)。
1.2.3 基本營養成分測定 隨機取規格相同的北海養殖與乳山野生的成品富齒巖蟲各30條,用清水沖洗干凈后,濾紙吸干表面水分,用真空冷凍機進行冷凍干燥,最后粉碎機粉碎至粉末,分裝保存備用。參照GB/T 5009.3—2016 (恒溫干燥法)、GB/T 5009.5—2016 (凱氏定氮法)、GB/T 5009.6—2016(索氏提取法)、GB/T 5009.4—2016 (550 ℃ 灼燒法),分別測定蟲體中水分、粗蛋白、粗脂肪、灰分的含量。
1.2.4 氨基酸和脂肪酸測定 參照1.2.3中方法獲取樣品粉末,每個地區樣品均設置3個平行組,每組樣品質量保證大于1 g,實驗時統一取1 g樣品,參照GB 5009.124—2016和 GB 5009.168—2016方法,應用日立L-8800型氨基酸分析儀和島津GC-2010型氣相色譜儀,分別測定北海養殖和乳山野生的富齒巖蟲體內18種氨基酸和脂肪酸含量。
1.2.5 營養評價 參照FAO/WHO氨基酸評分標準模式[22]和全雞蛋蛋白模式[23],對富齒巖蟲進行氨基酸評分 (AAS)、化學評分 (CS)、必需氨基酸指數 (EAAI) 計算[24-26],評價富齒巖蟲的營養價值。計算公式為:

式中:n為必需氨基酸數量;a1、a2、···、an為待測蛋白質中的必需氨基酸含量 (mg·g-1);A2、···、An為全雞蛋蛋白質中相應的必需氨基酸含量 (mg·g-1)。
1.3 數據處理
利用SPSS 23.0和Excel 2013軟件進行統計學分析,數據結果均以“平均值±標準差表示,用獨立樣本t檢驗分析北海養殖與乳山野生的富齒巖蟲的差異顯著性,顯著性水平設定為0.05。
2 結果
2.1 北海人工養殖富齒巖蟲的環境條件與生長情況
試驗期間,水溫和鹽度變化見圖1。從10月至翌年6月,養殖池水溫度呈開口向上的二次曲線變化趨勢,全程平均水溫為 (22.6±2.8) ℃,最高水溫為 (29.9±1.0) ℃,最低為 (12.6±2.5) ℃;鹽度的波動較緩,介于24.4~31.2,平均為28.2。
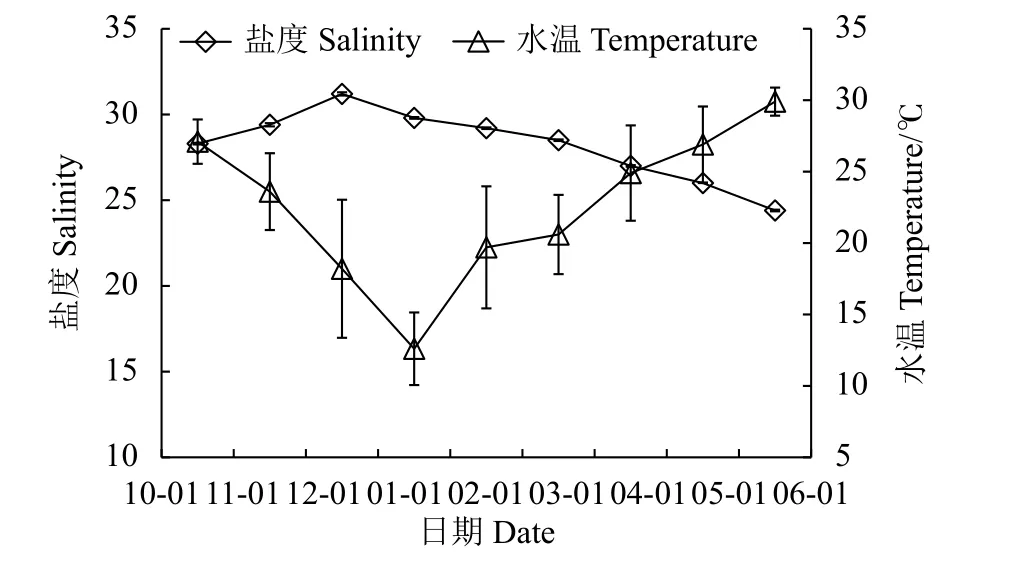
圖1 試驗期內海水鹽度與溫度變化Figure 1 Change in sea water salinity and temperature during experiment
人工養殖的富齒巖蟲平均體長、體寬、體質量隨時間變化見圖2-a—2-c。結果顯示,蟲體平均體長實現快速增長,由初始平均體長 (1.00±0.20) cm生長至 (12.66±1.51) cm,體長增長達12倍 (圖2-a)。平均體寬由 (0.10±0.02) cm 增至 (0.68±0.08) cm (圖 2-b)。至試驗結束時,蟲體由初始平均體質量 (0.03±0.01) g增至 (2.30±0.87) g (圖 2-c)。
富齒巖蟲在海泥和粗砂2種不同底質內表現出較為顯著的生長差異 (圖2-a—2-c)。結果顯示,0~4個月內,海泥底質組 (A) 的平均體長、體寬和體質量增長值顯著高于粗砂底質組 (B)。4~8個月養殖周期內,粗砂底質組的巖蟲平均體長、體寬和體質量增長均高于海泥組。統計學分析顯示,底質對富齒巖蟲體長、體寬和體質量增長的影響差異極顯著 (P<0.01)。
以體質量變化計算各底質組的特定生長率見圖2-d。試驗期間不同底質的富齒巖蟲特定生長率均表現出明顯的前高后低變化趨勢,養殖前期(0~4個月) 特定生長率顯著高于中后期 (4~8個月)。2種不同底質在不同養殖階段也存在差異,養殖前期,海泥底質組 (A) 特定生長率為2.27%~2.31%,高于粗砂組 (B) (1.83%~2.17%, P<0.05);養殖中期 (4~6個月),粗砂底質組 (B) 特定生長率為1.44%~1.51%,顯著高于海泥底質組 (A) (0.75%~0.81%, P<0.05);養殖后期 (6~8 個月),兩底質組的富齒巖蟲特定生長率差異不顯著 (P>0.05)。
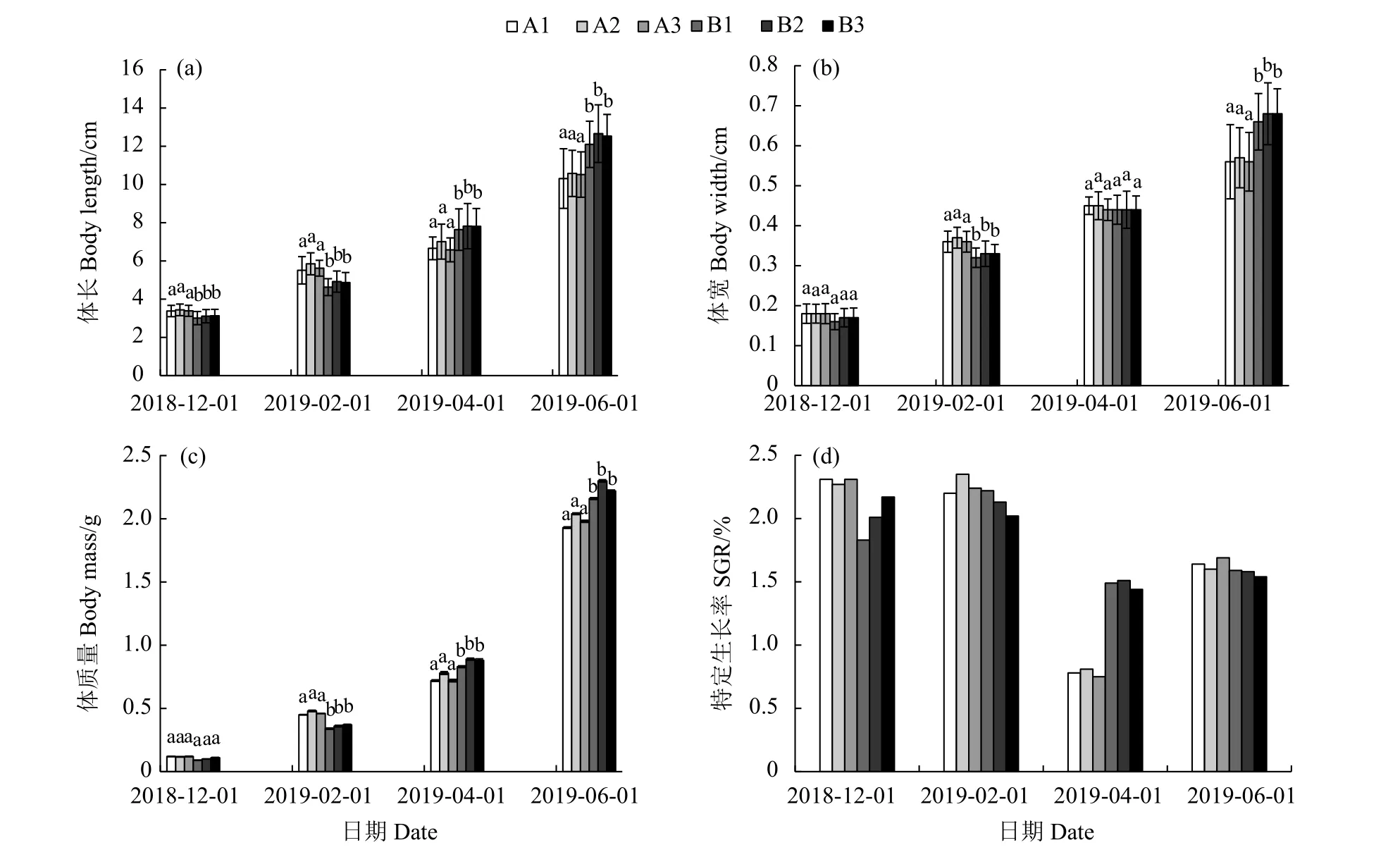
圖2 不同底質組富齒巖蟲體長、體寬、體質量和特定生長率隨時間變化的情況Figure 2 Body length, body width, body mass and special growth rate of M. maxidenticulata with different substrates along with time
2.2 北海養殖與乳山野生富齒巖蟲的基本營養成分分析
北海養殖與乳山野生的富齒巖蟲基本營養組成見表1。北海養殖蟲體的粗蛋白占比為60.96%,低于乳山野生蟲體 (64.24%)。而北海養殖蟲體內的粗脂肪占比達24.13%,顯著高于乳山野生蟲體(19.71%,P<0.05)。灰分與水分含量在2種不同來源的富齒巖蟲中也表現出顯著差異性,其中北海養殖蟲體的含水率高于乳山野生,但其灰分含量低于乳山野生蟲體 (P<0.05)。
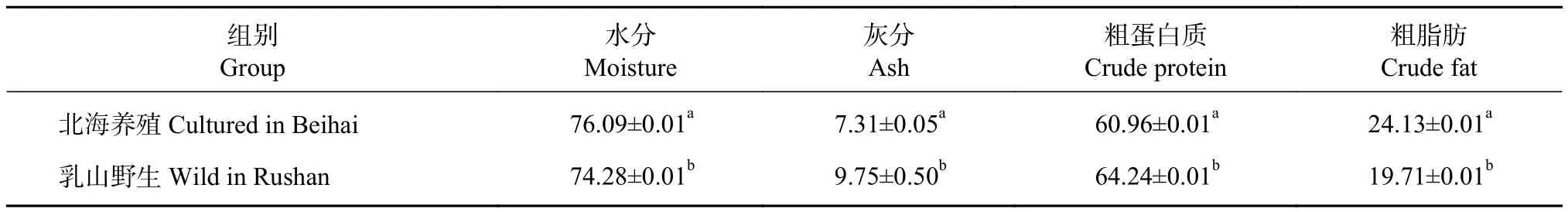
表1 北海養殖與乳山野生的富齒巖蟲基本營養組成 (干質量, N =30)Table 1 Basic nutritional composition of M. maxidenticulata cultured in Beihai and wild M. maxidenticulata in Rushan (dry mass) %
2.3 北海養殖與乳山野生富齒巖蟲的氨基酸組成分析
對比養殖與野生巖蟲氨基酸組成 (表2),2種不同來源的富齒巖蟲體內均含有18種氨基酸,且氨基酸組成基本相同,但北海養殖蟲體中蛋氨酸含量 (1.00%) 顯著高于乳山野生蟲體(0.75%,P<0.05),而其他17種氨基酸兩者無顯著性差異(P>0.05)。養殖與野生的蟲體中均含有4種呈味氨基酸 (DAA)、8種人體必需氨基酸 (EAA)、2種半必需氨基酸 (SEAA)和7種非必需氨基酸 (NEAA)。兩者的DAA分別為20.51%和20.05%,DAA/TAA分別高達41.23%和41.69%,在DAA和所有氨基酸中,谷氨酸含量均為最高 (分別為7.12%和7.09%);而在18種氨基酸中色氨酸含量最低 (均為0.50%)。北海養殖蟲體氨基酸總量 (TAA)、DAA、NEAA、SEAA和EAA,以及SEAA/TAA均略高于乳山野生蟲體,而DAA/TAA、NEAA/TAA略低于乳山野生蟲體,但差異不顯著。
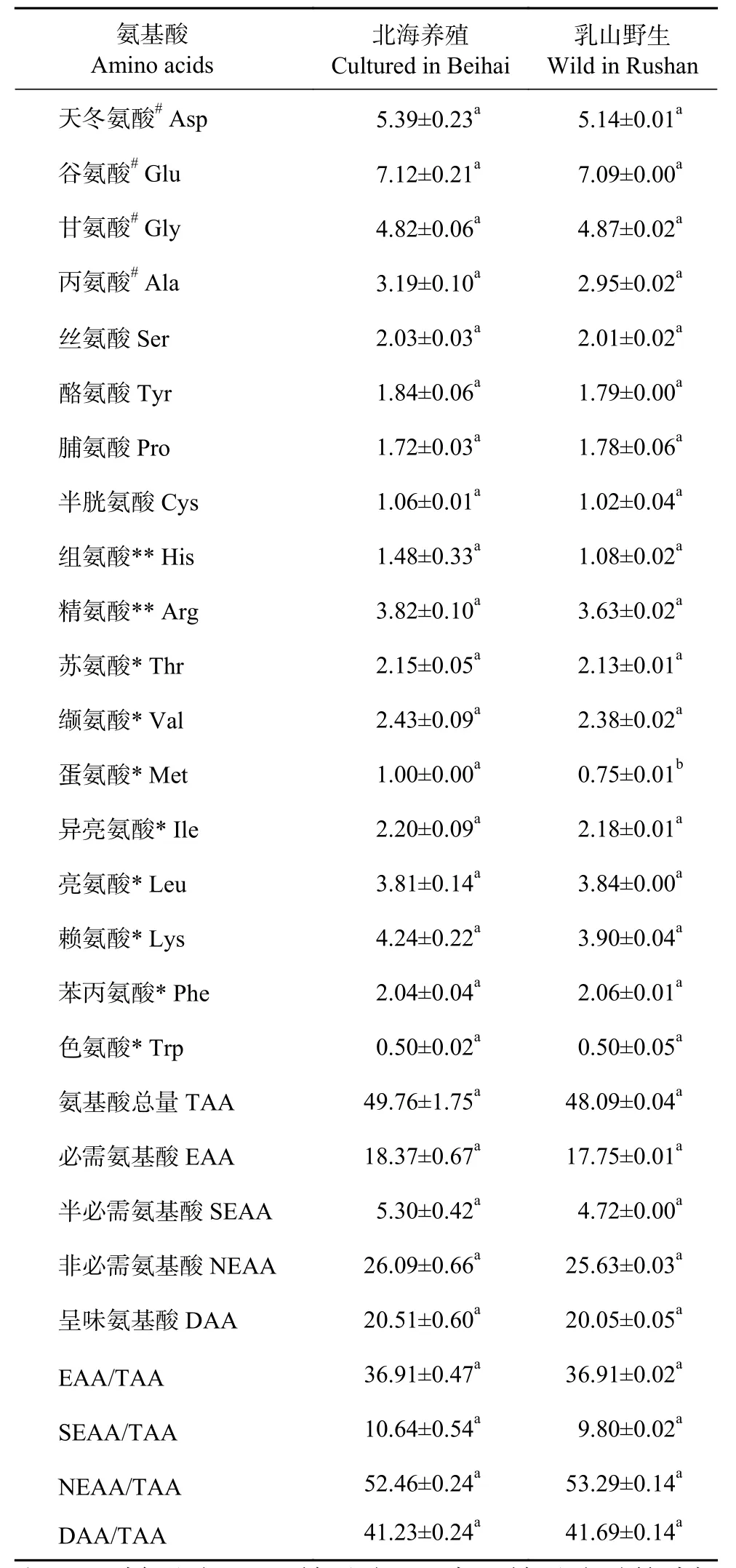
表2 北海養殖與乳山野生的富齒巖蟲氨基酸組成Table 2 Amino acid compositionof M. maxidenticulata cultured in Beihai and wild M. maxidenticulata in Rushan %
2.4 北海養殖與乳山野生富齒巖蟲的氨基酸營養評價
通過AAS和CS法對富齒巖蟲進行了氨基酸營養指標評價 (表3)。從AAS來看,2種不同來源的蟲體,第一限制性氨基酸均為纈氨酸 (Val),第二限制性氨基酸均為色氨酸 (Trp)。從AAS結果來看兩者賴氨酸 (Lys) 均為最高。從CS來看,北海養殖蟲體中只有色氨酸含量低于0.60,其CS為0.52;而乳山野生蟲體中有2種氨基酸含量低于0.60,分別為色氨酸和纈氨酸。此外,在AAS和CS中,北海養殖蟲體的必需氨基酸評分和CS均高于乳山野生蟲體。EAAI作為氨基酸營養評價中的重要指標,北海養殖蟲體為69.91,明顯高于乳山野生蟲體的65.28。
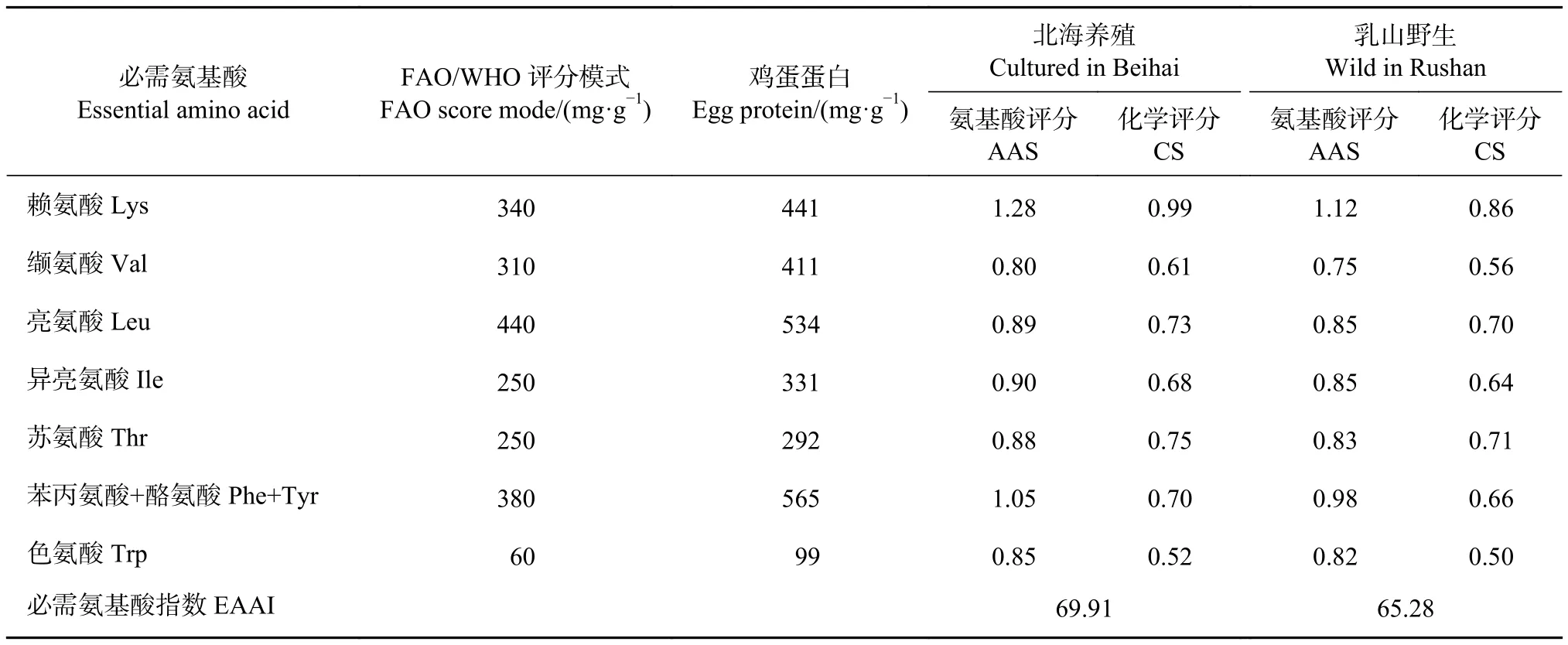
表3 北海養殖與乳山野生的富齒巖蟲氨基酸營養評價Table 3 Amino acids nutritional evaluation of M. maxidenticulata cultured in Beihai and wild M. maxidenticulata in Rushan
2.5 北海養殖與乳山野生富齒巖蟲的脂肪酸組成分析
2種不同來源的富齒巖蟲體內分別含有27種脂肪酸,但北海養殖蟲體未檢出十七碳一烯酸(C17:1),而乳山野生蟲體未檢出花生三烯酸 (C20:3 n-6) (表4)。棕櫚酸、油酸和亞油酸在2種蟲體內均為含量較高的脂肪酸,為主要脂肪酸。從脂肪酸類型來看,北海養殖蟲體的飽和脂肪酸與多不飽和脂肪酸含量 (30.47%、36.12%) 略低于乳山野生蟲體 (31.70%、37.76%),而單不飽和脂肪酸 (33.41%)含量顯著高于乳山野生蟲體 (30.54%,P<0.05)。在單不飽和脂肪酸中,北海養殖蟲體的油酸 (順) (C18:1 n-9c) 顯著高于乳山野生蟲體 (P<0.05),而神經酸C24:1 n-9顯著低于乳山野生蟲體 (P<0.05)。在多不飽和脂肪酸方面,北海養殖蟲體除亞油酸 (順)C18:2 n-6c和花生二烯酸C20:2顯著高于乳山野生蟲體外,花生四烯酸、EPA和二十二碳六烯酸(DHA) 等重要多不飽和脂肪酸均顯著低于乳山野生蟲體。北海蟲體的∑n-3 PUFA/∑n-6 PUFA為0.20%,顯著低于乳山野生蟲體的0.51% (P<0.05)。

表4 北海和乳山野生的富齒巖蟲脂肪酸含量Table 4 Fatty acid compositionof M. maxidenticulata cultured in Beihai and wild M. maxidenticulata in Rushan %
3 討論
3.1 富齒巖蟲的南移養殖
“北物南養”是我國水產從業者根據南北方氣候條件差異與生物生長特點,在長期生產實踐下創造出特色鮮明的水產養殖方式。該方式充分利用南方冬季溫度適宜、生物生長快的特點,將北方地區的養殖生物南移,加速其生長、縮短養殖周期、提高生物產量。重要經濟物種如皺紋盤鮑 (Haliotis discus hannai) 等均通過季節性南移養殖,產生了較大的生產規模與經濟效益[27]。富齒巖蟲作為北方地區的重要經濟性多毛類動物,其生長周期較長,在北方沿海由幼體至成體性成熟約需2~3年[28],而較長的生長周期與資源可持續利用之間存在一定矛盾,制約了富齒巖蟲在北方地區的人工養殖,進而導致資源數量不足,價格高昂。參考皺紋盤鮑等重要經濟物種季節性南移養殖模式,本研究將山東乳山地區的大規格巖蟲苗種南移至廣西北海,期望能夠利用南北方氣候條件的季節性差異,縮短其生長周期,實現“北蟲南養”,獲得良好的養殖效果。結果表明,秋季北方氣溫下降時,將富齒巖蟲大規格苗種南移至廣西北海,通過人工養殖,取得了較為顯著的生長收益。試驗周期內,北海地區養殖巖蟲在8個月內體質量平均可達2.30 g,最大體長可達15.70 cm,達到了商品規格,成功實現了富齒巖蟲的南移養殖。根據乳山地區人工養殖經驗與報道,北方達到相同規格約需36~48個月[28-29]。由此可見,南移養殖可顯著提高富齒巖蟲的生長速度、縮短養殖周期,是切實可行的多毛類動物生產方式。究其原因,作為潮間帶的重要底棲動物,富齒巖蟲對溫度、鹽度具有較強的耐受能力,而北海地區冬季平均水溫為 (18.2±4.8) ℃,與富齒巖蟲生長適溫較為接近,為其生長提供了良好的生態環境,促進了富齒巖蟲的快速生長。
在南移養殖過程中,通過對比2種不同底質中富齒巖蟲的生長參數,發現其在不同底質內表現出較顯著的生長差異。試驗前期,海泥底質組富齒巖蟲的特定生長率顯著高于粗砂底質組,而養殖中后期,粗砂底質組中富齒巖蟲的特定生長率顯著高于海泥底質組。造成這種現象的原因是由于底質作為底棲動物生長、棲息等生命活動的主要場所,其顆粒大小、穩定程度、表面構造、營養成分以及深度對底棲動物的生長有顯著影響。泥底富含有機質顆粒,但透水、透氣性較差,而粗砂底質正好相反。由于工廠化養殖的生物密度通常較高,幼體階段生物個體較小、占據空間較少,因此,有機質含量豐富的海泥底質更能促進其快速生長。養殖中后期,大型個體需要良好的底內生活環境,粗砂底質由于空隙大、透水、透氣性好等原因,更能促進生物生長。這也為富齒巖蟲工廠化養殖的底質選擇提供了一定參考。
3.2 多毛類動物的營養
多毛類動物由于營養組成豐富,富含多種不飽和脂肪酸,是海洋生態系統和水產養殖中的重要餌料生物。特別是在經濟性蝦、蟹的親體促熟培育中,是不可替代的優質活體餌料[30-31]。富齒巖蟲作為經濟性蝦、蟹親體促熟培育中的優質生物餌料,通過人工養殖后,特別是南移養殖,大幅縮短生長周期的情況下,其營養組成變化是決定其作為活體生物餌料價值的重要因素。本研究結果表明,南移養殖的富齒巖蟲粗營養成分與乳山野生個體有一定差異,表現為水分和粗脂肪含量高于乳山野生富齒巖蟲,粗蛋白含量略低。在氨基酸組成方面,南北2種富齒巖蟲體內均含有18種氨基酸,但南移養殖蟲體內必需氨基酸含量高于乳山野生蟲體。南北富齒巖蟲EAA/TAA均接近40%,說明其體內蛋白質較為優質。相比于其他經濟性多毛類物種,富齒巖蟲的氨基酸含量略低于雙齒圍沙蠶 (Perinereis aibuhitensis)[32-33]。從AAS和CS結果來看,南移養殖與乳山野生的富齒巖蟲賴氨酸均最高,除纈氨酸低于0.80外,各種必需氨基酸含量較為均衡,接近FAO/WHO理想模式。此外,南移養殖的富齒巖蟲EAAI大于乳山野生蟲體,表明南移養殖的富齒巖蟲必需氨基酸總量更加接近標準蛋白質。因此,雖然兩者在蛋白質和氨基酸組成上存在一定差異,但南移養殖富齒巖蟲的部分指標卻優于乳山野生種。
從脂肪酸組成來看,南移養殖的富齒巖蟲體內飽和脂肪酸總量和總多不飽和脂肪酸占比與乳山野生蟲體之間無顯著差異。但EPA、DHA和花生四烯酸等多不飽和脂肪酸占比顯著低于乳山野生種,∑n-3/∑n-6也略低于乳山野生蟲體。EPA、DHA是公認的在對蝦性腺成熟中具有重要促進功能的優質多不飽和脂肪酸,而花生四烯酸是前列腺素合成的前體,在斑節對蝦 (Penaeus monodon) 性腺成熟調控中具有重要作用[19]。此外,飼料中的∑n-3/∑n-6越接近于對蝦生殖腺的組成比例 (大部分蝦中約為1),越有利于其促進蝦蟹的性腺成熟[34]。本試驗結果顯示,南移養殖的富齒巖蟲其體內重要多不飽和脂肪酸占比低于野生蟲體,說明南移養殖雖然可以顯著提高富齒巖蟲的生長速度,縮短養殖周期,但對于其體內的關鍵營養成分,如必需氨基酸、EPA、DHA的合成與累積仍低于野生自然種群。究其原因,低溫和生長周期較長,對于動物的營養累積具有重要作用。已有研究表明,一定范圍內變溫可以提高某些水生動物攝食量、降低水生動物基礎代謝、提高食物轉化率和同化率,促進生長,如中國對蝦等[35];而低溫條件下,水生動物攝食量和基礎代謝降低,生理和生化發生變化,如低溫條件下羅非魚 (Oreochromissp.) 和鋸緣青蟹 (Scylla serrata) 的脂肪酸總量降低,但不飽和脂肪酸含量升高等[36-37]。因此,如何提高南移養殖蟲體體內關鍵不飽和脂肪酸含量,對于促進其在水產經濟蝦、蟹親體培育中的應用,具有重要的理論和實踐意義。
4 結論
在8個月的養殖周期內,富齒巖蟲在廣西北海地區可生長至2.30 g。養殖前期海泥底質幼蟲生長迅速,中后期粗砂底質更適合其生長。相比于乳山野生種群,南移養殖的富齒巖蟲在粗營養成分含量、氨基酸和脂肪酸組成方面差異不大,但EPA、DHA和花生四烯酸等重要多不飽和脂肪酸含量方面,與乳山野生種群仍有差異。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
