基于DNA條形碼的南海美濟礁潟湖魚卵種類鑒定與組成初探
2021-12-22 05:56:46盧芷程李紅婷蔣佩文陳曉雷曹漪婷陳作志
南方水產科學 2021年6期
盧芷程 ,李 敏 ,張 俊 ,張 帥 ,李紅婷 ,蔣佩文 ,陳曉雷 ,曹漪婷 ,陳作志
(1. 中國水產科學研究院南海水產研究所/農業農村部外海漁業開發重點實驗室,廣東 廣州 510300;2. 上海海洋大學海洋科學學院,上海 201306; 3. 南方海洋科學與工程廣東省實驗室 (廣州),廣東 廣州 511458; 4. 大連海洋大學水產與生命學院,遼寧 大連 116023)
魚類浮游生物 (Ichthyoplankton) 是魚卵和仔稚魚的統稱。對其進行研究,可獲得魚類產卵場的直接證據,是漁業生物學和資源養護研究的重要前提[1]。以傳統形態學方法鑒定魚類浮游生物并非易事,其依賴于研究者的經驗,且對于形態差異小的近緣物種難以區分[2-3]。目前中國已報道的海洋魚類種類高達3 000多種[4],但是已鑒定的魚卵和仔稚魚的種類不到300種[5]。通常已鑒定的仔稚魚種類數僅占現存魚種的十分之一,而已鑒定的魚卵種類數也約為仔稚魚種類數的十分之一[6]。
DNA條形碼技術是利用生物體DNA中的一段保守片段對物種進行快速準確鑒定的新興技術。對于魚類的鑒定,DNA條形碼通常基于線粒體細胞色素C氧化酶亞單位I (Cytochrome coxidase subunit I,簡稱COI) 基因[7]。隨著DNA數據庫的建立和完善 (如NCBI),以及擴增技術的發展和成本的降低,基于分子標記的DNA條形碼技術已成為魚卵鑒定的重要手段,并已得到廣泛應用[8]。如Ardura等[9]利用分子標記對北大西洋的魚類浮游生物的研究發現,分子鑒定比形態鑒定具有更高的效率;王利華等[10]聯合COI和Cytb基因對6種鲌屬魚類進行分子鑒定和系統發育分析,表明將COI和Cytb作為DNA條形碼能夠區分出近緣種。陳定標等[11]運用不同方法對浙江沿海魚卵仔稚魚樣品進行鑒定,結果顯示魚卵分子鑒定比形態鑒定識別到種的比例更高。目前,國內也開展了不少基于分子標記對魚卵的研究[12-14],但是以往研究主要集中于中國近海、淡水魚類。Chen等[3]利用COI標記對南海東沙群島南部珊瑚礁海域采集的70個仔魚樣品進行鑒定和分析,結果顯示優勢科為天竺鯛科和鰕虎魚科;吳娜等[15]對南沙永暑礁獲得的71個魚卵樣品進行分子鑒定,認為雙斑櫛齒刺尾魚 (Ctenochaetus binotatus) 是該海域的優勢種。南海珊瑚礁魚類浮游生物的研究尚處于起步階段。
南沙群島及其附近海域是中國唯一位于珊瑚礁核心區的海域,而位于南沙群島東端的美濟礁(115°32'E, 9°54'N),是南沙群島最大的環礁[16]。美濟礁由外坡、礁坪和潟湖三部分組成,礁體呈環形,中部為潟湖,面積約36 km2,水深20~30 m,正南和西南方向各有一進出水道,屬于典型的低潮高地珊瑚礁[17-18]。熱帶海洋季風氣候,以及潟湖獨特的水文環境,使其擁有較為穩定的高溫條件和鹽度[16]。美濟礁水文環境優良,浮游植物種類豐富[19],是南海珊瑚礁魚類產卵育苗的重要場所,而目前對于美濟礁魚卵資源的研究尚有空缺。因此,本研究選取南海美濟礁為研究對象,基于DNA條形碼技術對其春、秋兩季魚卵的物種組成和季節動態進行了研究,旨在為南海島礁種質資源的保護和利用提供基礎信息。
1 材料與方法
1.1 樣本采集
依托中國水產科學研究院南海水產研究所“南鋒”號漁業調查船,于2019年5月 (春季) 和9月(秋季) 在南海美濟礁潟湖進行魚卵采集。
每個季節設立5個站點 (春季C1—C5,秋季Q1—Q5),采樣范圍為 115°521'E—115°560'E、9°886'N—9°906'N (圖 1)。樣本采集根據《海洋調查規范》(GB 12763.6—2007) 進行,具體為采用浮游動物淺海Ⅰ型網 (網口內直徑50 cm,網目0.505 mm),置于小艇船尾水平拖網采集,拖網時間10 min,船速1.5 kn。樣本用乙醇 (體積分數≥70%)保存,并于-20 ℃貯藏至運回實驗室。
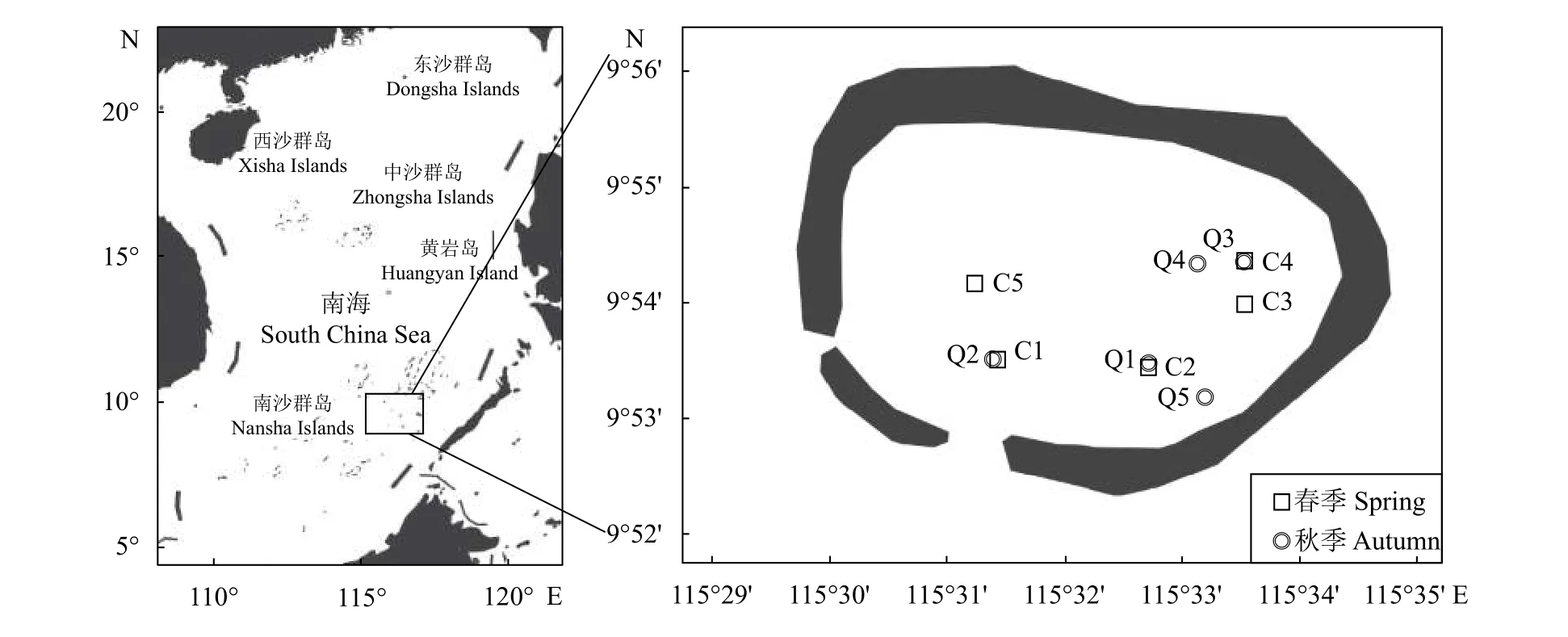
圖1 美濟礁調查站點圖C1—C5為春季采樣點;Q1—Q5為秋季采樣點;該圖基于審圖號為GS (2019) 5218號的標準地圖制作。Figure 1 Sampling stations in Meiji ReefC1-C5. Sampling stations of spring; Q1-Q5. Sampling stations of autumn; this map was made based on the standard map No. GS (2019) 5218.
1.2 魚卵種類鑒定
挑取單個魚卵,使用ddH2O洗脫乙醇,采用海洋動物組織DNA提取試劑盒 (天根,中國) 提取基因組DNA,-20 ℃保存備用。COI基因序列擴增引物為Fish-F1: 5'-TCAACCAACCACAAAGA CATTGGCAC-3',和Fish-R1: 5'-TAGACTTCTGG GTGGCC AAAGAATCA-3'[20]。使用擎科生物Golden Star T6 Super PCR Mix (1.1×) 試劑盒,擴增體系 (總體積 25 μL)為:Mix 預混液 22 μL,正反向引物各1 μL,DNA模板1 μL。PCR反應采用降落 PCR 程序:98 ℃ 2 min;98 ℃ 10 s,65 ℃ 15 s,72 ℃ 4 s,10 個循環;98 ℃ 10 s,62 ℃ 15 s,72 ℃ 4 s,10 個循環;98 ℃ 10 s,58 ℃ 15 s,72 ℃4 s,10 個循環;98 ℃ 10 s,55 ℃ 15 s,72 ℃4 s,5個循環;最后72 ℃延伸4 min。用1%凝膠電泳檢測PCR擴增產物,選取包含目的片段的產物,于廣州擎科有限公司進行進行PAGE純化及測序。
獲得目的片段后,使用軟件MEGA 5.01[21]排列比對,并進行剪切對齊。使用軟件DNAsp[22]提取單倍型。對單倍型進行物種信息比對的數據庫為美國國家生物技術信息中心 (NCBI, https://www.ncbi.nlm.nih.gov/) 數據庫和生命條形碼數據系統(BOLDSYSTEMS, http://www.boldsystems.org/index.php) 數據庫。比對結果采用以下標準判別:相似度≥99%為同種;相似度92%~99%,則定為同屬[23]。
根據陳真然[24]的研究,將鑒定到種的魚卵按生態分布類型劃分為礁棲性魚類、大洋性中上層魚類和深海魚類3類。
1.3 數據分析
1.3.1 物種豐度和優勢度的計算 物種豐度 (G)和優勢度 (Y) 計算公式為:

式中:G表示單位體積海水中的魚卵個數(粒·m-3);N為全網魚卵個體數 (粒);V為濾水量(m3)[25]。

式中:ni表示第i個種類的物種密度 (粒·m-3);fi表示第i個種類的出現頻率[25]。本調查中,優勢種以Y≥0.001為劃分標準。物種豐度和優勢度的計算均使用Excel 2010 軟件完成。
1.3.2 群落結構的季節變化 為比較不同季節的魚卵種類群落結構的相對豐度和變化程度。使用R語言的pheatmap包來分析不同季節各站點的魚卵種類分布,對樣本進行歸一化處理,選用系統默認聚類方法。本研究中,相對豐度介于0~4,相對豐度≥3為優勢種,以橙色填充作為標記。采用物種更替率 (E) 比較兩季節的魚卵組成差異,E的計算使用Excel 2010 軟件完成。采用分類學多樣性指數平均分類差異指數Δ+和分類差異變異指數Λ+分析群落多樣性[26-27],Δ+是指任意2個隨機選取的不同物種間路徑長度的理論平均值,該值不隨物種數量而改變;Λ+表示Δ+偏離程度的平均值,反映群落中魚卵種間關系分布的均勻程度[26]。相比于傳統多樣性計算方法,基于分類學地位的Δ+和Λ+不受采樣方法、棲息地生態類型自然變化等影響,可解釋群落的種間關系[26,28]。分類學多樣性指數Δ+和Λ+使用軟件Primer 5.2的Taxdtest軟件包進行計算。不同指數的計算公式為:
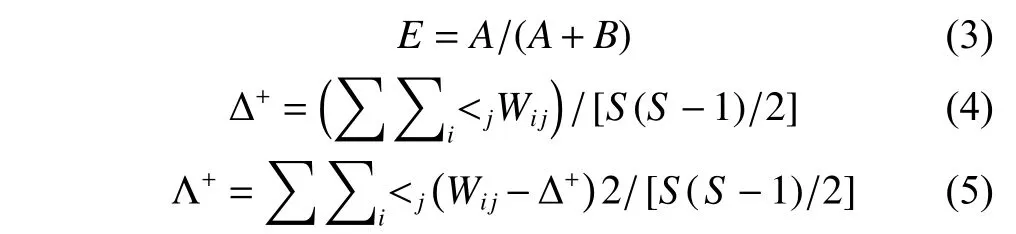
式中:A為兩季節間減少或者新增加的種類數量;B為兩季節共有的物種數量[29];wij為第i和j個種類在分類系統樹中的路徑長度;S為種類數。將門、綱、目、科、屬、種6個分類等級間的加權路徑長度的權重設置為100.000、83.333、66.667、50.000、33.333、16.667[30]。
2 結果
2.1 魚卵數量統計
采集樣本總數為345粒 (其中春季219,秋季126,表1);春季魚卵密度為0.95粒·m-3,秋季為0.54粒·m-3,春季是秋季的1.76倍。
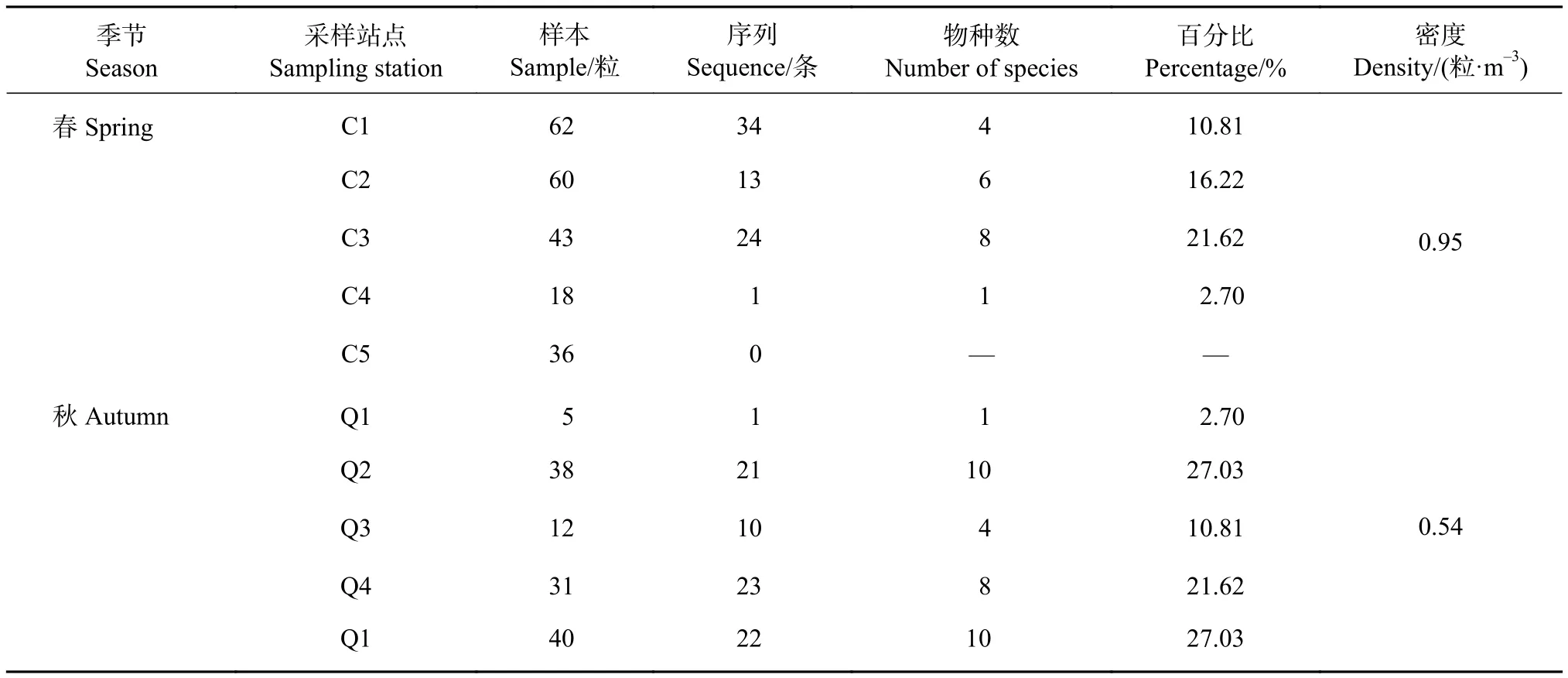
表1 各站點序列數、魚卵種數及其占比Table 1 Number of sequences, kinds of species and proportion in each site
2.2 魚卵種類組成
成功擴增149條序列,實驗成功率為43.19%(春季72條,秋季77條)。與數據庫比對,共鑒定出37種魚卵,隸屬于3目23科31屬,其中4種僅鑒定到屬,分別為鈍鰕虎魚屬未定種1 (Amblygobiussp.1)、眶棘鱸屬未定種1 (Scolopsissp.1)、眶棘鱸屬未定種2 (Scolopsissp.2) 和弱蜥魚屬未定種 1 (Scopelosaurussp.1) (表 2)。
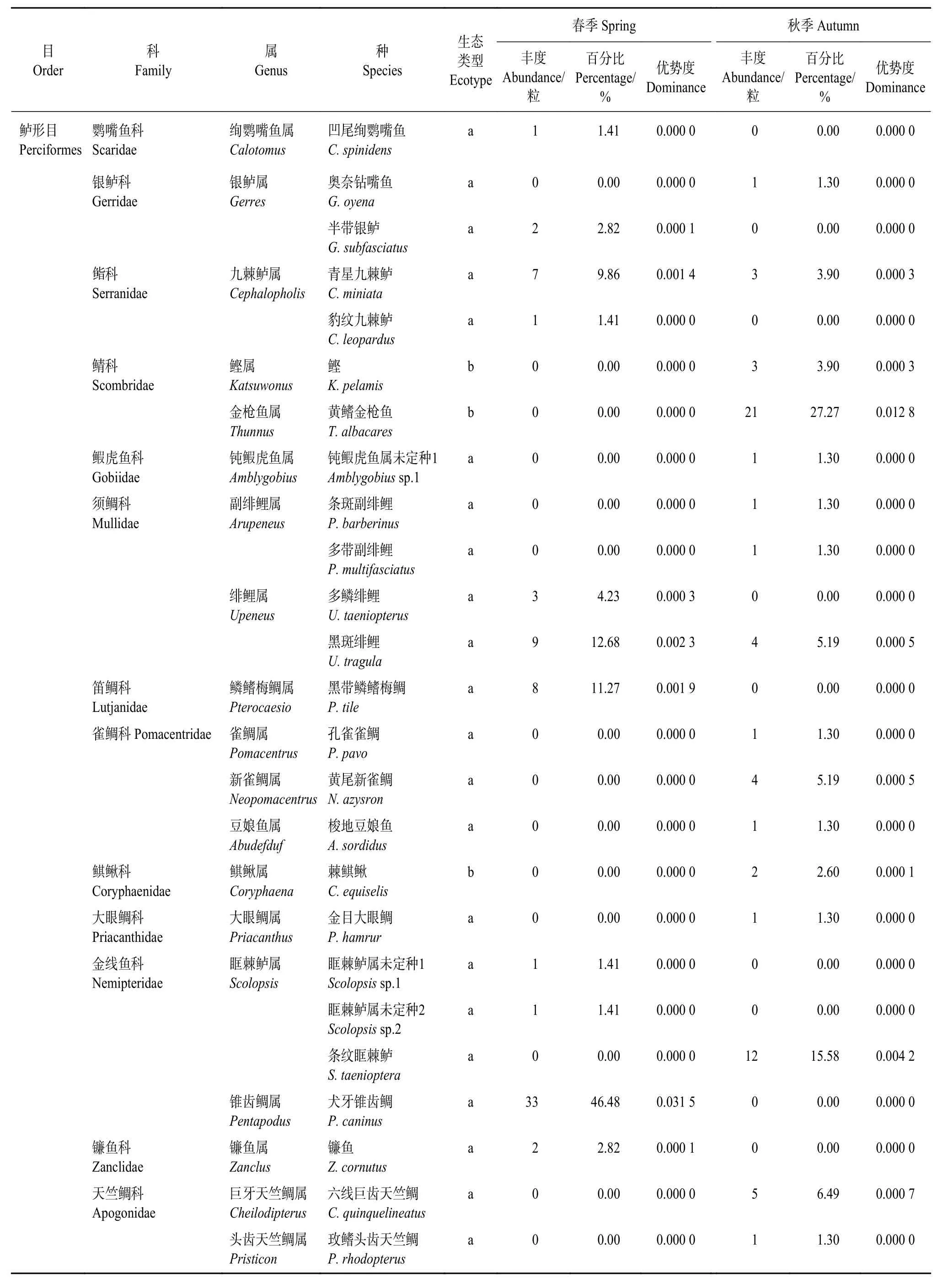
表2 2019年美濟礁春、秋季魚卵種類組成Table 2 Species composition of fish eggs collected from Meiji Reef in spring and autumn, 2019
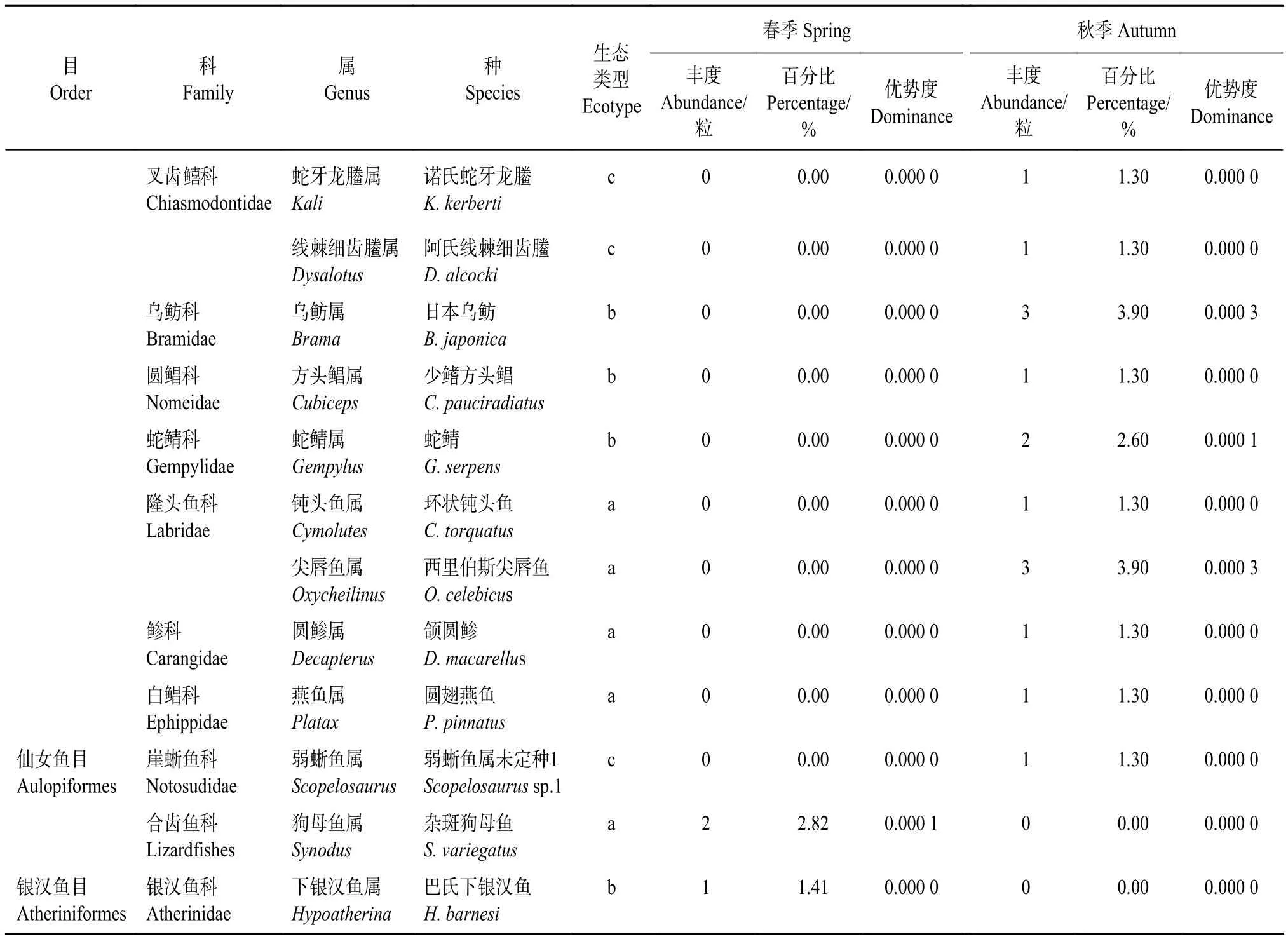
續表2 to be continued
鱸形目魚卵種類和數量占比最大,魚卵種類數為34種,占比91.89%,序列總數為145條,占比97.31%,其余為仙女魚目2種,銀漢魚目1種。根據生態分布,可分為三大類型[24]:礁棲類群魚卵 (28種),大洋性中上層類群魚卵 (6種) 以及深海類群魚卵 (3種)。
本調查中發現頜圓鲹 (Decapterus macarellus)、圓翅燕魚 (Platax pinnatus)、環狀鈍頭魚(Cymolutes torquatus)、玫鰭頭齒天竺鯛 (Pristicon rhodopterus)、西里伯斯尖唇魚 (Oxycheilinus celebicus)、巴氏下銀漢魚 (Hypoatherina barnesi) 等產卵種類在珊瑚礁海域的捕撈調查中未見記錄。表明基于分子鑒定的魚卵資源調查具有更好的發現隱秘種的能力。
2.3 魚卵種類組成的季節變化
根據數量分布統計結果,不同季節的種類數量組成有所差異,春季魚卵種類為13種,秋季為26種,春秋季均出現的魚卵種類有青星九棘鱸(Cephalopholis miniata) 和黑斑緋鯉 (Upeneus tragula) 2種,隸屬于鱸形目。春季特有魚卵種類11種,分別屬于燈籠魚目、銀漢魚目和鱸形目;秋季特有的魚卵種類24種,均屬于鱸形目;春秋兩季魚卵種類更替率為94.59%。獲得的37種魚卵中鱸形目豐度最高,其在春季和秋季魚卵組成中分別占95.83%和98.70%。
春季優勢種是黑帶鱗鰭梅鯛 (Pterocaesio tile,優勢度0.001 9,豐度百分比11.27%)、犬牙錐齒鯛(Pentapodus caninus,優勢度0.031 5,豐度百分比46.48%)、黑斑緋鯉 (優勢度0.002 3,豐度百分比12.68%) 和青星九棘鱸 (優勢度0.001 4,豐度百分比9.86%) 4種,而秋季優勢種是黃鰭金槍魚 (Thunnus albacares,優勢度0.012 8,豐度百分比27.27%)和條紋眶棘鱸 (Scolopsis taenioptera,優勢度0.004 2,豐度百分比15.58%) 2種 (表2)。
因為C5站點未獲得有效的物種信息,故只以9個站點的物種種類構建相對豐度的熱圖。結果顯示 (圖2),春季4個站點相對豐度最高的為犬牙錐齒鯛 (C1, C2)、青星九棘鱸 (C2)、黑斑副緋鯉(C3)、黑帶鱗鰭梅鯛 (C4);秋季5個站點相對豐度最高的魚卵物種為圓翅燕魚 (Q1)、六線巨齒天竺鯛 (Q2)、條紋眶棘鱸 (Q3, Q4)、黃鰭金槍魚 (Q4,Q5)。其中,圓翅燕魚是Q1站點唯一獲得的魚卵物種信息,因而不具有代表性。
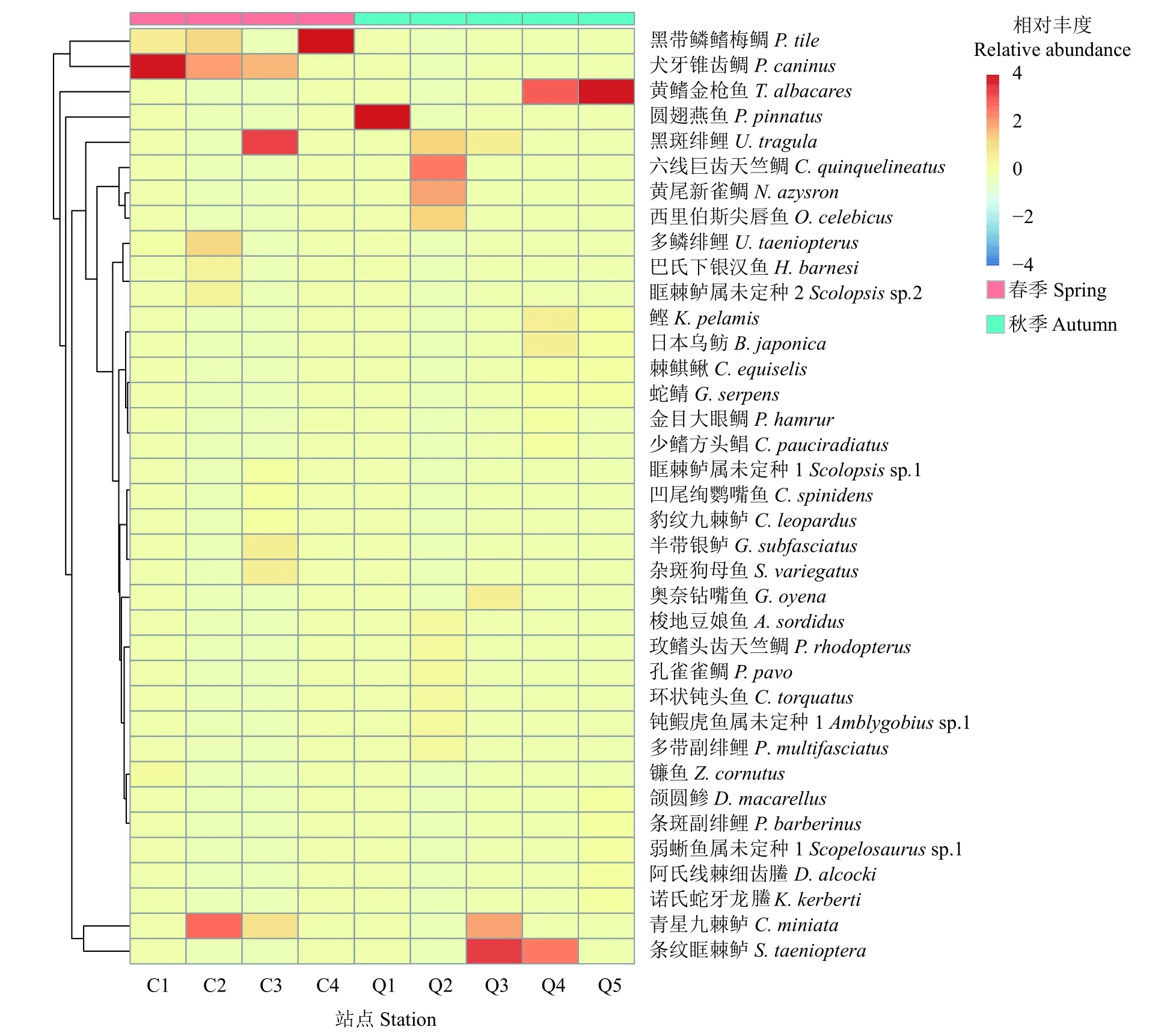
圖2 2019年美濟礁春、秋季魚卵種類組成和相對豐度的比較因C5站點沒有獲得有效序列,只選取9個站點的物種信息進行分析。以站點 (列) 為樣本進行標準化,比較各物種相對豐度差異,以0為基準,4為相對豐度最大值。Figure 2 Comparison of species composition and relative abundance of fish eggs collected from Meiji Reef between spring and autumn in 2019Since there was no valid sequence in C5 site, only nine sites were selected for species information analysis. The data of each station (column) was used as one sample for standardization, and the relative abundance of each species was compared, with 0 as a benchmark and 4 as the maximum.
優勢度和相對豐度的結果共同表明,美濟礁春、秋季的魚卵優勢種差異較大,產卵種類呈現季節性變動。
美濟礁春季魚卵的Δ+為53.63,Λ+為111.45;秋季魚卵的Δ+為50.77,Λ+為31.03 (圖3)。表明美濟礁春季產卵種類之間的親緣關系比秋季產卵種類要更遠,并且物種分類關系的均勻度遠低于秋季。
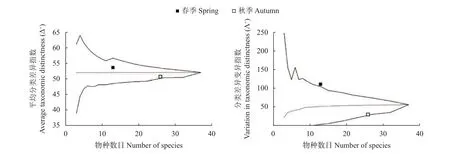
圖3 美濟礁春、秋季魚卵平均分類差異指數 (Δ+) 和分類差異變異指數 (Λ+) 95%置信漏斗曲線Figure 3 Funnel plot for average taxonomic distinctness (Δ+) and variation in taxonomic distinctness (Λ+) with 95% confidence limit of eggs species in Meiji Reef
3 討論
3.1 美濟礁魚卵種類組成
此次調查共獲得37種魚卵,包含礁棲性魚類、大洋性中上層魚類和深海魚類。根據中國水產科學院南海水產研究所1998—2018年的調查數據,美濟礁礁棲魚類達166種,其中鱸形目魚類占77%[31]。在此次調查中,鱸形目魚卵為主要組成部分,數量占總數的97.31%,符合南海魚類群落構成特征。與研究資料對比發現,凹尾絢鸚嘴魚 (Calotomus spinidens) 和犬牙錐齒鯛在2012—2018年美濟礁魚類資源調查中未發現[31],而在本研究中再次發現了以上2個物種的存在,并新記錄了多種南海珊瑚礁產卵種類。一般認為氣候變化和人類活動會引起資源變動[32],但在珊瑚礁資源調查中,由于采樣方法和采樣工具的限制,獲得的魚類種類可能具有選擇性。絕大多數硬骨魚類產出的魚卵于海水中受精發育,魚卵數量巨大,且大多具有浮性[24]。受精卵在水中隨水流漂浮,該特性使其易于被采集。因此利用魚卵對珊瑚礁魚類種類組成進行調查,可作為傳統采樣調查方法的有效補充手段。
另外,本研究首次在美濟礁發現了黃鰭金槍魚的魚卵。黃鰭金槍魚具有重要經濟價值,其在赤道和熱帶海洋性氣候的海域全年均可繁殖,產卵行為多發生于28 ℃[33-34],而美濟礁的海表溫度在27.6~29.2 ℃,平均為28.4 ℃[35],符合其繁殖條件。早期有研究報道了黃鰭金槍魚仔稚魚在南海的分布,3—8月在南海中、西沙均有捕獲[36];Zhang等[37]采用回聲測深儀對南海黃鰭金槍魚幼魚進行了調查,漁獲的幼魚體長337~458 mm;李炎璐等[38]曾在美濟礁附近海域捕獲52尾黃鰭金槍魚幼魚。上述研究雖然都捕獲到了幼魚,但由于金槍魚游泳能力強,難以確定其產卵場的具體范圍。本研究為南沙美濟礁珊瑚礁海域作為黃鰭金槍魚產卵場之一提供了直接證據。
3.2 美濟礁魚卵種類組成與分布特征的季節變化
美濟礁魚卵的資源密度季節變化較小,春季魚卵密度 (0.95 粒·m-3) 是秋季 (0.54 粒·m-3) 的 1.76倍。王九江等[39]對大亞灣魚卵的調查顯示,春季魚卵密度約為0.98粒·m-3,夏季為1.21粒·m-3,秋季為0.17粒·m-3,冬季為0.09粒·m-3,結果顯示大亞灣海域春季魚卵密度是秋季的5.76倍,夏季魚卵密度是冬季的13.44倍,認為溫度、溶解氧、鹽度和pH對魚卵有重要影響。美濟礁年均氣溫變化幅度在1 ℃左右,全年溫度差異不明顯[35]。與近海相比,美濟礁珊瑚礁海域處于熱帶,具有充足的光照和穩定的氣候條件,且潟湖內鹽度條件較為穩定。Isari等[40]對紅海珊瑚礁附近海域幼魚的調查顯示,近岸的幼魚豐度具有明顯的季節變化。與紅海珊瑚礁的片狀分布不同,美濟礁的礁盤呈環狀,礁盤內環境相對更穩定。Zhao等[41]對南沙美濟礁和渚碧礁珊瑚覆蓋率進行研究,發現兩島礁的珊瑚平均覆蓋率為21%,表明南沙群島南部的珊瑚群落處于相對健康的狀態。因此認為美濟礁的氣候環境全年適宜魚類繁殖。
美濟礁春、秋兩季的產卵種類差異很大,且春、秋季各站點魚卵種類組成不均。秋季,Q4和Q5采樣點所獲得的魚卵大部分屬于大洋性種類和深海類群,剩余部分為鱸形目種類;靠近這2個點的Q3和Q1種類和數量均較少,均為礁棲類群的魚卵;而Q2有較豐富的種類,均屬礁棲魚類。春季,與Q4和Q5相近的采樣點中,未發現大洋性種類和深海類群,且魚卵的種類數與其他點無明顯差別。推測大洋性魚類和深海種類的繁殖行為可能會導致礁棲魚類產卵地點的轉移甚至遷出,但具體關系有待持續觀察并進一步研究以提供更多信息。
優勢度和密度的結果共同表明,不同季節的魚卵優勢種種類組成差異較大。與優勢度相似,相對豐度分析可顯示群落優勢物種,且結果十分明了,但在樣本數量極少的情況下,相對豐度結果可能會與實際數量產生差異。因此,使用相對豐度分析優勢種時,在有條件的情況下,應盡量豐富樣本總量,從而避免誤差。
美濟礁春、秋季的Δ+和Λ+結果反映了春季產卵種類之間的親緣關系比秋季要更遠,而物種分類關系的均勻度遠低于秋季,兩者均低于1998—2018年的年平均值[31]。與附近生境永暑礁相比(表3),美濟礁產卵種類中,不僅有典型的珊瑚礁礁棲魚類,還存在大洋性中上層種類和深海種類,產卵種類之間的親緣關系要更遠,故而美濟礁的Δ+要更高。
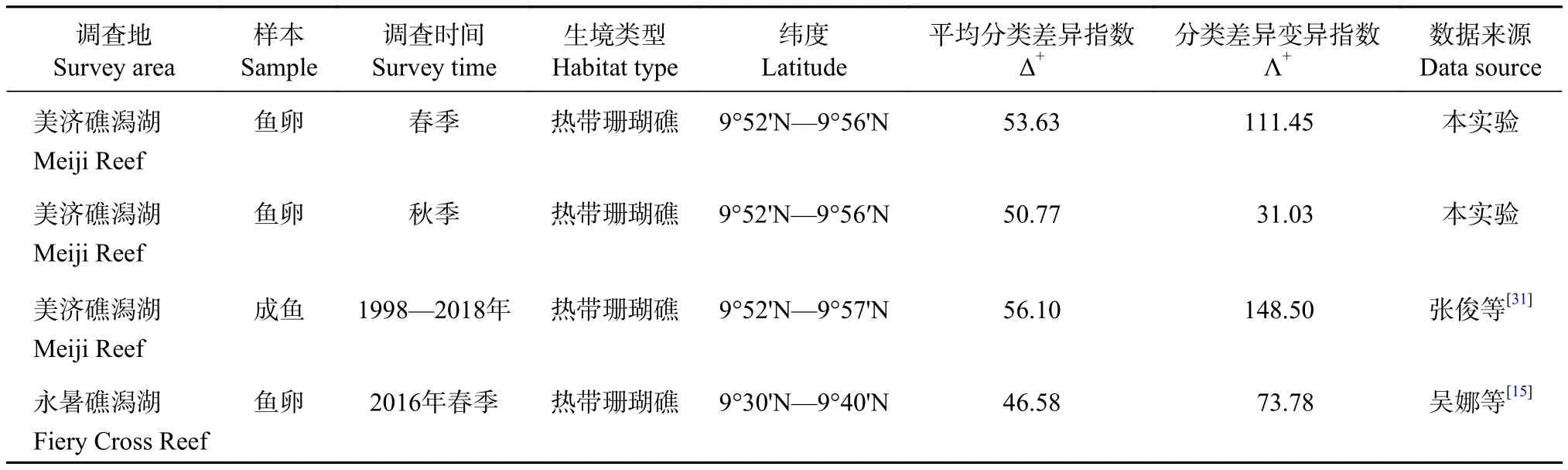
表3 相似生境的魚卵平均分類差異指數 (Δ+) 及分類差異變異指數 (Λ+)Table 3 Average taxonomic distinctness (Δ+) and variation in taxonomic distinctness (Λ+) in similar economical systems
4 結論
基于DNA條形碼的美濟礁春、秋季的魚卵種類組成和多樣性分析發現,美濟礁產卵種類多樣性較豐富,魚卵密度季節性變動較小。美濟礁的魚類產卵行為具有一定的季節偏好性,產卵物種更替現象明顯,而南海魚類繁殖行為是否同樣存在季節偏好性,需要今后進行長期、持續性調查研究。南海美濟礁的產卵魚類除了有礁棲性魚類外,還有多種深海魚類和大洋性中上層魚類,說明南海珊瑚礁生態系統在維護深海和大洋生態系統穩定性中起重要作用,建議將美濟礁作為珊瑚礁魚類重要產卵場進行全年保護。
另外,本實驗中序列擴增的成功率并不高,可能由于部分魚卵腐敗,導致DNA降解,可考慮在船上即時進行魚卵核酸的提取,從而減少核酸降解帶來的影響。相比依賴于手釣、延繩釣、流刺網等傳統采樣方法,基于分子鑒定的魚卵采集調查方法對環境和生物資源破壞更小,且有利于發現隱秘種和未知種,是一種有效的資源調查補充手段。
致謝:感謝廣東海洋大學侯剛老師給予的技術指導,感謝蔡研聰、楊玉滔和“南鋒”號船員在樣品采集上的幫助,感謝王棟梁、方藝菲和李媛潔等在論文寫作上給予的幫助!
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
