生物被膜態和游離態臨床耐碳青霉烯類鮑曼不動桿菌轉錄組的差異分析*
2021-12-14 01:06:04劉巍巍國果吳兆穎毛成菊李忠旬賈利娜尚小麗彭建吳建偉
貴州醫科大學學報 2021年11期
劉巍巍,國果,吳兆穎,毛成菊,李忠旬,賈利娜,尚小麗,彭建**,吳建偉**
(1.貴州醫科大學 基礎醫學院 人體寄生蟲學教研室,貴州 貴陽 550025;2.貴州醫科大學 基礎醫學院 現代病原生物學特色重點實驗室,貴州 貴陽 550025;3.貴州醫科大學 生物工程學院,貴州 貴陽 550025)
鮑曼不動桿菌(Acinetobacterbaumannii,AB)是一種非發酵性革蘭陰性菌,屬于條件致病菌。碳青霉烯類抗生素是臨床上抗菌譜最廣,治療耐藥菌導致的嚴重感染的主要抗菌藥物[1]。目前,耐碳青霉烯類鮑曼不動桿菌(carbapenem-resistant AB,CRAB)的檢出率不斷上升,提示著臨床抗感染治療面臨重大挑戰[2],而AB極易形成生物被膜,使臨床治療更加棘手[3]。生物被膜是細菌粘附于生物或非生物體表面、被自身細胞分泌的基質(胞外多糖、蛋白質及DNA基質)所包裹而形成的特殊結構,可表達與游離態細菌完全不同的基因,在形態、理化性質及抗生素敏感性等方面都有明顯的不同[4]。有研究報道,細菌形成生物被膜后對抗生素的敏感性降低,抗生素的最低抑菌濃度將提高10~1 000倍[5]。因此,進行生物被膜及其形成機制的研究對臨床細菌感染的防控有重要意義。轉錄組是指細菌在某一特定功能或狀態下、所有轉錄RNA的總和,研究臨床CRAB游離態和生物被膜態細菌的生理性狀和基因表達狀況,將為AB生物被膜形成機制和耐藥菌感染的治療提供科學依據[6]。有學者對AB臨床分離株1656-2的生物被膜態菌和游離態菌進行了蛋白質組的比較,發現與處理環境信息、參與代謝、細菌耐藥及基因修復有關的蛋白發生了表達變化[7]。李帥[8]對耐藥AB菌株BJAB0868的生物被膜和對數生長期細菌進行了轉錄組分析,發現2種狀態下菌株的基因表達確實存在差異,有些基因在生物被膜態中顯著升高,在游離生長狀態中極少表達甚至被抑制。但是單個菌株的比較具有特異性,且不同地區的菌株特性存在差異[9]。因此,本研究選取4株不同的臨床CRAB作為4個生物學重復臨床樣本,對其生物被膜態和游離態進行轉錄組學測定和差異分析,以期進一步明確臨床CRAB生物被膜和游離態細菌的差異,為其生物被膜的產生機制以及藥物作用靶點的研究提供科學依據。
1 材料與方法
1.1 實驗材料
1.1.1菌株 CRAB 4、55、78及117分離于住院患者痰液、血液臨床樣本,大腸埃希菌ATCC25922和銅綠假單胞菌ATCC27853為質控菌,所有菌株由病原生物學實驗室保存。
1.1.2主要試劑與儀器 Mueller-Hinton(MH)肉湯培養基、Trypticase Soy Broth(TSB)肉湯培養基及Luria-bertani(LB)固體培養基(北京索萊寶),溶菌酶(上海生工),SYBR Premix Ex TaqTMⅡ(大連寶生物),TRIZOL(美國Invitrogen);恒溫培養箱(上海博訊實業),細菌比濁儀(上海昕瑞),高壓蒸汽鍋(日本ALR),微量加樣槍、聚合酶鏈式反應(polymerase chain reaction,PCR)擴增儀及速冷凍離心機(德國Eppendorf),ABI7300熒光定量PCR儀(美國ABI),NanoDrop ND-2000分光光度計(美國Thermo)。
1.2 實驗方法
1.2.1臨床菌株鑒定及生物膜態和游離態菌株制備 CRAB 4、55、78及117作為4個生物學重復,采用法國生物梅里埃VITEK-2全自動微生物分析儀進行分析鑒定與藥敏試驗;同時,擴增菌株16SrRNA和rpoB基因測序和分析,進一步確定菌株鑒定的準確性。挑取單菌落加入MH培養基中,于37 ℃、220 r/min搖床培養至對數期,用TSB培養基稀釋至106cfu/L,取菌液4 mL加6孔培養板,置37 ℃恒溫箱培養24 h,棄游離菌液,PBS清洗3次,細胞刮刀刮取生物被膜態菌(A組);挑取單菌落加入MH肉湯培養基中,于37 ℃、220 r/min搖床培養至對數期,收集游離態菌體(B組)。
1.2.2Trizol法提取RNA與質量檢測 參考文獻[10]方法,取A組和B組樣品各1 mL于4 ℃、8 000 r/min離心1 min;棄上清,加400 mg/L溶菌酶100 μL,震蕩混勻,室溫酶解5 min;4 ℃、3 000 r/min離心5 min,棄上清,加TRIZOL溶液1 mL,震蕩裂解,冰上靜置15 min;加氯仿200 μL,顛倒混勻,冰上靜置5 min;4 ℃、12 000 r/min離心15 min,吸取上清液,轉入新的離心管中。加入等體積的異丙醇,輕輕顛倒混勻,室溫靜置10 min;4 ℃、12 000 r/min離心10 min,棄上清,加預冷的用焦炭酸二乙酯(diethyl pyrocarbonate,DEPC)水配制75%乙醇1 mL,輕輕吹打混勻,溶解沉淀;4 ℃、7 500 r/min離心5 min,棄上清,室溫干燥10 min;加DEPC水30~50 μL溶解沉淀,分裝,置于-80 ℃保存;瓊脂糖凝膠電泳檢測RNA降解情況,分光光度計測定RNA濃度。
1.2.3文庫構建和測序 提取到的RNA經檢測合格后,去除Total RNA中的核糖體RNA(rRNA),獲得mRNA。隨后加入fragmentation buffer,將得到的mRNA隨機打斷成短片段,按照鏈特異性建庫的方式建庫[11],即以片段化的mRNA為模板,隨機寡核苷酸為引物,在M-MuLV逆轉錄酶體系中合成cDNA第一條鏈;隨后用RNaseH降解RNA鏈,并在DNA polymerase I體系下,以dNTPs(將dNTP中的dTTP用dUTP取代)為原料合成cDNA第二條鏈(使第2條鏈中包含A/U/C/G);再用AMPure XP beads篩選250~300 bp的cDNA,之后進行cDNA末端修復、加A尾、連接測序接頭,然后使用USERTM(尿嘧啶-特異性切除試劑)酶降解含U的cDNA第二鏈,PCR擴增獲得文庫。文庫構建完成后,使用Qubit2.0 Fluorometer進行初步定量,使用Agilent 2100 bioanalyzer檢測文庫的insert size,符合預期后用實時熒光定量PCR(quantitative real-time PCR,qRT-PCR)對文庫有效濃度進行準確定量,以保證文庫質量。庫檢合格后,把不同文庫按照有效濃度及目標下機數據量的需求pooling后進行Illumina測序。
1.2.4讀取映射到參考基因組 以AB 030(NZ_CP009257.1)的基因組作為參考基因組,用Bowtie2軟件將過濾后的序列進行基因組定位分析[12]。
1.2.5差異表達基因(differentially expressed genes,DEGs)分析 HTSeq v0.6.1用于計算映射到每個基因的讀數。使用DESeq軟件進行A組與B組間的差異表達分析(每個組4個生物學重復)。使用Benjamini和Hochberg的方法來調整所得P值,設置差異基因篩選的標準[13]:校正后的P<0.05,且丨Log2(fold change)丨>1。
1.2.6DEGs的基因本體(gene ontology,GO)和京都基因與基因組百科全書(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析 通過GOSeq R軟件實現差異基因的GO富集分析,并以校正P<0.05的GO Term作為差異表達基因顯著富集,通過KOBAS軟件分析差異基因在KEGG通路中的富集。
1.2.7實時熒光定量PCR驗證差異基因 從轉錄組測序鑒定得到的DEGs中挑取8個差異基因(表1),利用qRT-PCR進行驗證。按之前所述方法提取RNA后,用逆轉錄試劑盒將提取的總RNA逆轉錄成cDNA。PCR擴增總反應體系20 μL:含cDNA模板2.0 μL、SYBR?Green Realtime PCR Master Mix 10 μL、ROX Reference Dye(50×)0.4 μL、ddH2O 6.0 μL、上下游引物各0.8 μL。反應條件:95 ℃預變性30 s,95 ℃變性5 s、60 ℃退火31 s,共40個循環。反應在ABI7300熒光定量PCR儀上進行,以AB 16SrRNA為內參基因,目的基因的相對表達量采用2-ΔΔCt方法計算。
2 結果
2.1 RNA質量
總RNA條帶完整清晰,無雜帶,樣本無污染(圖1);通過Nano Photometer spectrophotometer檢測RNA純度,OD260/280介于1.8~2.2,OD260/230>1.8,樣品滿足后續建庫和試驗要求。見圖1和表2。
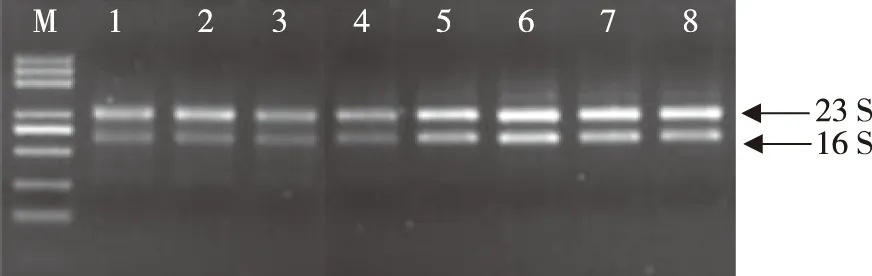
注:M為marker條帶,1~4為A組總RNA,5~8為B組RNA。圖1 RNA瓊脂糖凝膠電泳結果Fig.1 RNA agarose gel electrophoresis

表2 RNA樣品質量信息Tab.2 RNA sample quality information
2.2 測序數據
樣品測序統計結果顯示錯誤率為0.03%,可認為測序結果可靠;每組樣品的clean reads 數據>1 G,測序深度足夠覆蓋所有可能存在的基因。Q20和Q30分別為計算Phred 數值大于20和30的堿基占總體堿基的百分比,結果顯示Q20>95%且Q30>90%。見表3。
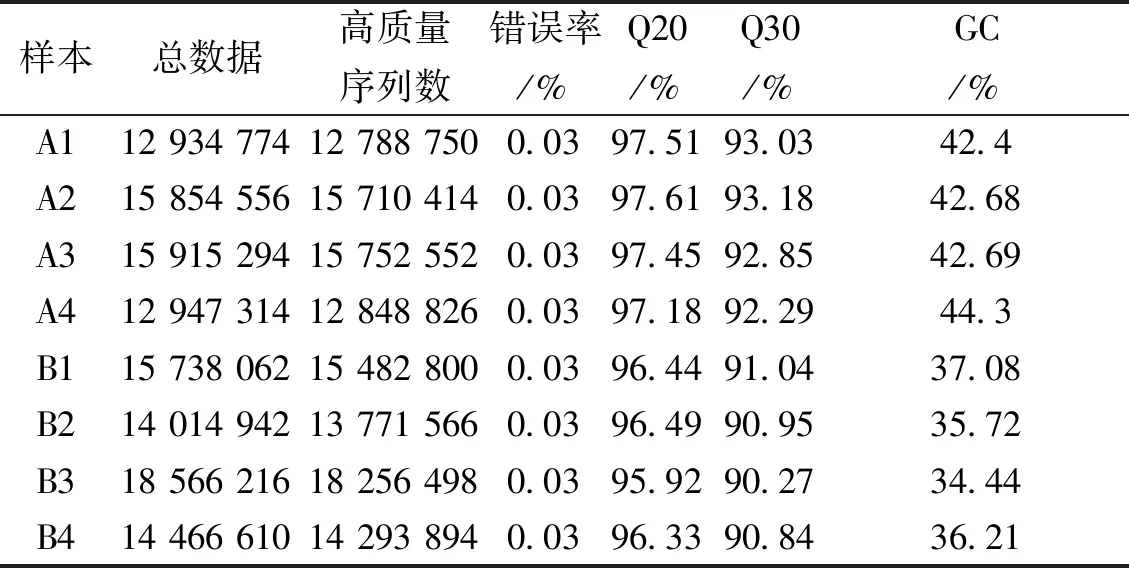
表3 測序數據匯總Tab.3 Sequencing data
2.3 參考序列比對分析
A組所產生的測序reads成功比對到基因組的總映射比例高于80%,其中具有多個定位的測序序列占總體的百分比低于6%;B組所產生的測序reads成功比對到基因組的總映射比例為60%左右,系樣品處理導致的變化,不影響測序效果。見表4。
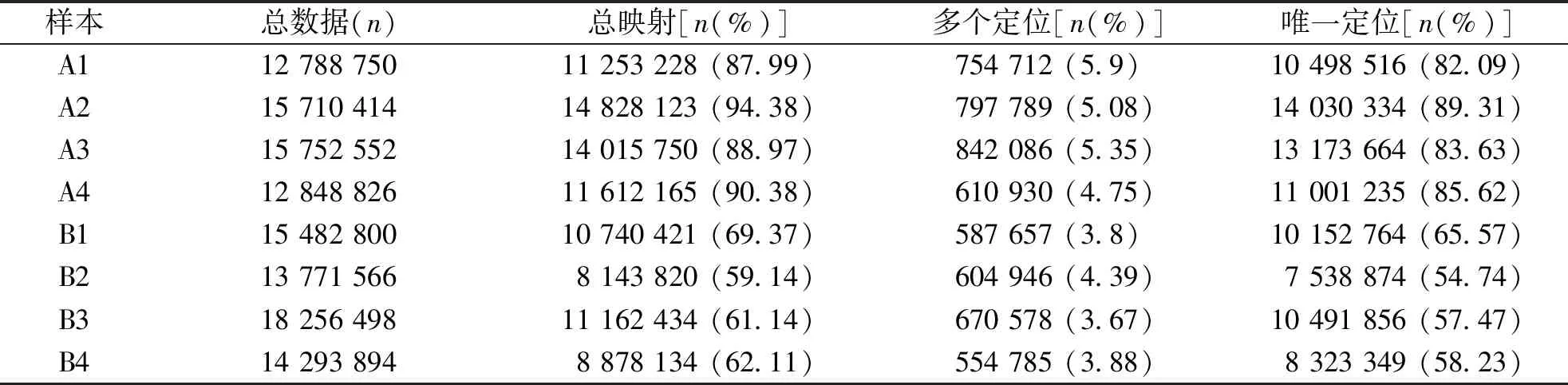
表4 樣本與參考基因組比對情況統計Tab.4 Statistics comparison of samples and reference genome
2.4 基因表達水平
不同表達水平區間的基因數量統計結果表明,各樣本的FPKM>1的基因數量約占總體的74%以上,提示有基因表達。見表5。
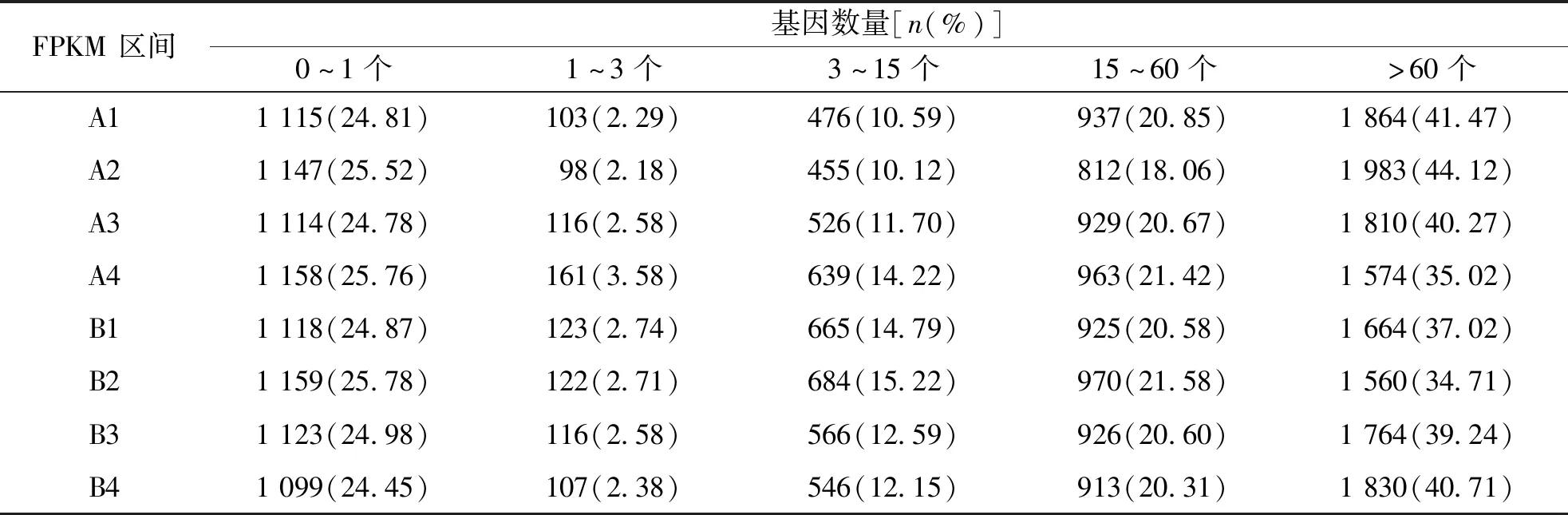
表5 不同表達水平區間的基因數量統計Tab.5 Statistics of gene numbers in different expression levels
2.5 DEGs的篩選
DEGs的火山圖顯示,臨床CRAB A組與B組相比有407個基因差異表達,包括212個上調和195個下調基因。見圖2。
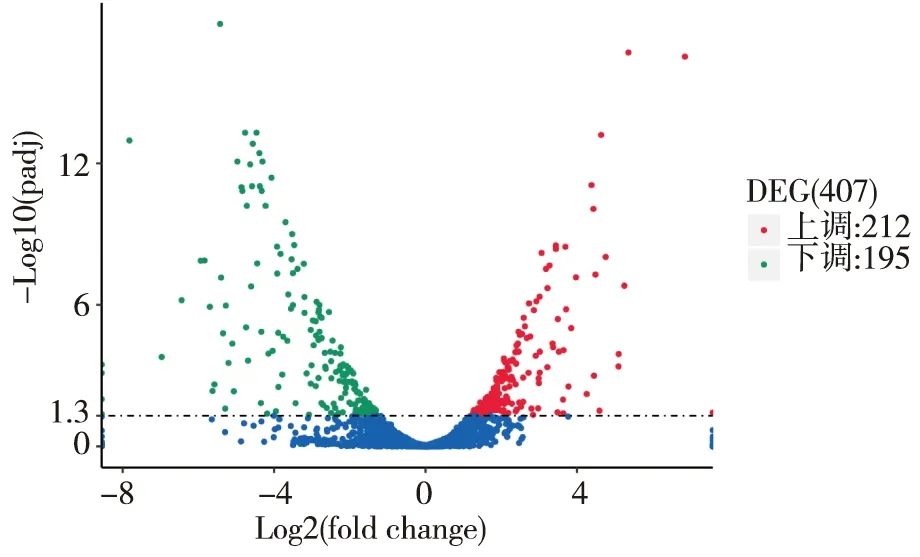
注:紅、綠點分別為上調和下調的DEGs,藍點為不顯著DEGs。圖2 DEGs火山圖Fig.2 Volcanoplot of DEGs
2.6 差異表達關鍵基因分析
上調的差異基因涉及鐵的攝取和轉運、核酸和生化代謝酶類、菌毛合成相關基因等,其中,TonB依賴的鐵載體受體(IX87_RS16650)、鐵氧還蛋白還原酶(IX87_RS14470)、鐵載體生物合成蛋白(IX87_RS18700)、亞鐵轉運蛋白A(IX87_RS15450)、鐵螯合酶(IX87_RS16135)等基因的Log2(fold change)值為1.5~5.1;NAD(P)依賴性氧化還原酶(IX87_RS20015)、乙酰輔酶A C-酰基轉移酶(IX87_RS06095)的Log2(fold change)值分別為3.7和3.4;菌毛合成相關基因CsuA(IX87_RS06910)的Log2(fold change)值為1.89。下調的差異基因涉及鎂、硫的轉運,無機物質的ABC轉運蛋白相關基因、生化代謝酶等,其中,磺酸鹽ABC轉運蛋白底物結合蛋白(IX87_RS14420)、硫酸鹽ABC轉運蛋白底物結合蛋白(IX87_RS09105 )、氨基酸ABC轉運蛋白通透酶(IX87_RS03165)的Log2(fold change)值分別為-5.9、-5.8及-4.9;α/β水解酶(IX87_RS09110)的Log2(fold change)值為-5.4,NADH氧化酶(IX87_RS04460)為-4.3。
2.7 差異基因GO富集分析
GO富集分析結果表明最顯著的30個GO條目,酰基輔酶A脫氫酶活性、輔助因子結合、維生素結合以及物質跨膜轉運蛋白活性等富集于分子功能類別,核苷轉運、氧化還原過程、脂質代謝過程、氨基酸的生物合成及金屬離子的轉運等富集于生物過程。見圖3。
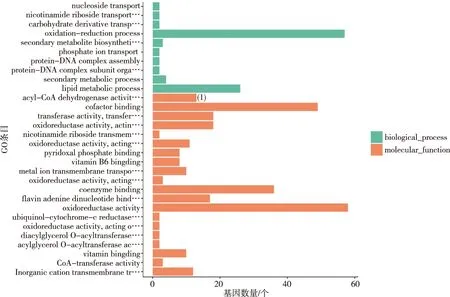
注:(1)P<0.05。圖3 DEGs的GO富集分析Fig.3 GO enrichment analysis of DEGs
2.8 差異基因KEGG富集分析
KEGG富集分析結果表明,DEGs參與的KEGG代謝通路中最顯著的通路有20條,分別為硫代謝(sulfur metabolism)、三磷酸腺苷(adenosine triphosphate,ATP)結合盒(ATP binding cassette,ABC)轉運系統(ABC transporters)、苯甲酸酯降解(fluorobenzoate degradation)、芳香化合物的降解(degradation of aromatic compounds)、纈氨酸、亮氨酸和異亮氨酸的降解(valine,leucine and isoleucine degradation)、β-丙氨酸代謝(beta-alanine metabolism)、卟啉和葉綠素代謝(porphyrin and chlorophyll metabolism)、丙酸酯代謝(propanoate metabolism)、丙酮酸代謝(pyruvate metabolism)、甲烷代謝(methane metabolism)、不飽和脂肪酸的生物合成(biosynthesis of unsaturated fatty acids)、糖酵解/糖異生(glycolysis/gluconeogenesis)、不同環境中微生物代謝(microbial metabolism in diverse environments)、甘油酯代謝(glycerolipid metabolism)、硒化合物代謝(selenocompound metabolism)、核黃素代謝(riboflavin metabolism)、碳代謝(carbon metabolism)、酮體的合成與分解(synthesis and degradation of ketone bodies)、丁酯酸代謝(butanoate metabolism)。另外,還涉及到萬古霉素耐藥(vancomycin resistance)、群體感應(quorum sensing)、陽離子抗菌肽耐藥[cationic antimicrobial peptide (CAMP)resistance]和細菌分泌系統(bacterial secretion system)等。見圖4。
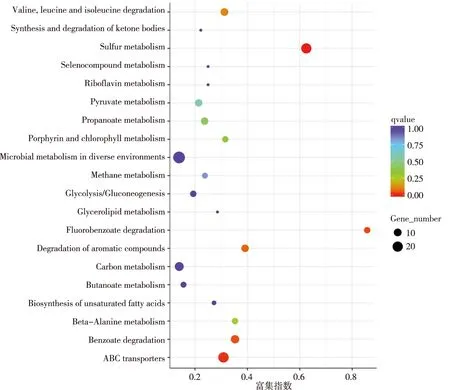
圖4 DEGs的KEGG富集分析Fig.4 KEGG enrichment analysis of DEGs
2.9 DEGs的驗證
從轉錄組差異基因中挑選8個基因,其中上調和下調表達的基因分別為5和3個;利用qRT-PCR進行驗證,該結果與轉錄組結果趨勢一致,表明轉錄組數據可靠。見圖5。
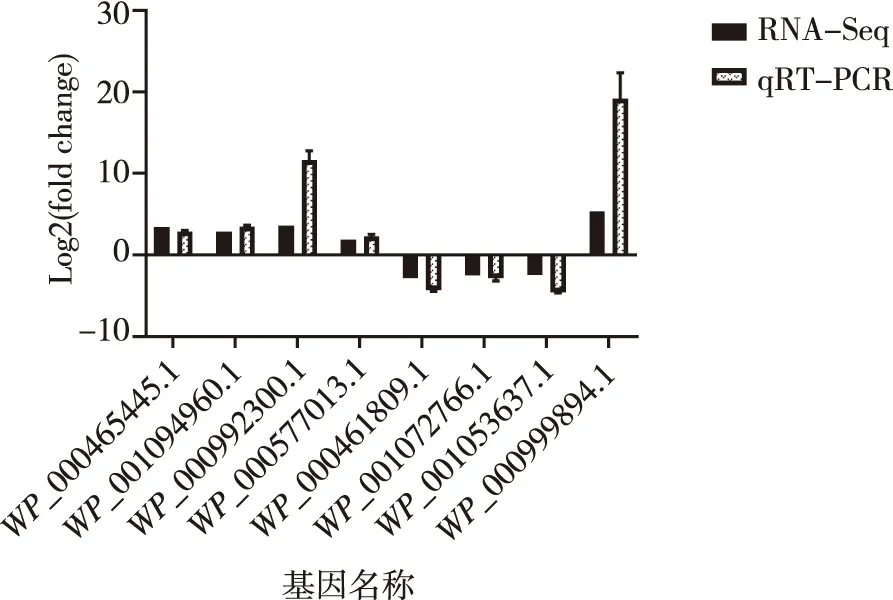
圖5 RNA-Seq和qTR-PCR的8個差異基因表達水平Fig.5 Analysis of eight differential genes expression levels between RNA-Seq and qTR-PCR
3 討論
傳統觀念上,細菌是獨立游離的個體,然而有研究表明,只有不到0.1%的微生物種群處于游離生長模式,大多數細菌以更復雜的生物被膜形式生存[14]。研究表明,細菌生物被膜可引起至少65%人類感染,以生物被膜形式存在的細菌對抗菌藥物、外界環境壓力和宿主免疫系統的抵抗能力增強,生物被膜相關感染的治療是臨床的重大難題[15]。本研究中,通過轉錄組學測序比較臨床CRAB生物被膜態細菌和游離態細菌的差異,探究臨床CRAB生物被膜形成的機理。以往研究中,一般選取同1株菌作為樣本,且只設3個生物學重復[8,10]。而生物學重復對測序實驗的設計及實驗數據的解讀和分析都非常重要,測序的樣本越多,越能夠降低背景差異,增強結果的可靠性[16]。本實驗選取4株來源不同,但形態及理化特性相似、生物被膜形成能力強的臨床分離CRAB作為4個生物學重復的樣本進行測序,得到的測序數據更具可靠性。測序結果表明,與游離態菌相比,CRAB生物被膜菌中有212個DEGs上調、195個DEGs下調,這些差異基因主要涉及鐵、鎂、硫的攝取和轉運、菌毛合成相關基因、生化代謝酶類、轉錄調節因子、核酸及無機物質的ABC轉運蛋白相關基因等。有文獻報道,鐵濃度與生物被膜的形成有關,在低濃度的鐵-Ⅲ的存在下,檢測到較高含量的群體感應自誘導信號分子酰基高絲氨酸內酯[17]。鐵的攝取和轉運等DEGs的變化證明細菌生物被膜的形成過程中需要鐵的參與。本研究中DEGsTonB依賴的鐵載體受體、亞鐵轉運蛋白及鐵螯合酶等顯著上調,表明生物被膜形成后,細菌對鐵的需求增強,鐵的轉運和攝取對生物被膜有重要影響。菌毛CsuA的表達上調,提示菌毛加速合成,促進生物被膜的形成。關于生物被膜形成的耐藥機制,有一種觀點是營養限制學說,生物被膜內營養限制,細菌生長速度變慢和代謝減緩,導致生物被膜態的細菌對抗菌藥物的敏感性降低[18-19]。轉錄組測序結果中生化代謝酶類相關基因的表達下調驗證了這個觀點。此外,也有一些未知基因(假設蛋白)在轉錄組測序比較中出現明顯上調或下調,這些基因在其中的作用還需進一步探究。
對DEGs進行GO富集和KEGG富集,有助于進一步探究在生物被膜形成過程中起作用的信號通路[20]。結果表明,硫代謝、ABC轉運及苯甲酸酯降解等通路顯著富集。值得注意的是,DEGs也富集到雙組份系統、萬古霉素耐藥、核苷酸切除修復、群體感應、β-內酰胺抗性及抗生素的生物合成這些通路中。ABC轉運系統將ATP水解與各種底物的主動運輸結合在一起,有利于細菌耐藥性的轉移及生物被膜態細菌的物質交流[21-22]。ABC轉運蛋白是已知的最大蛋白家族之一,并廣泛存在于細菌中,將ATP水解與各種底物(例如離子、糖、脂質、固醇、肽、蛋白質及藥物)的主動運輸結合在一起[23-24]。游離態細菌與生物被膜態細菌的區別可能涉及各種物質參與的能量交換,以及物質的合成與代謝。
萬古霉素耐藥通路的變化提示生物被膜的形成增強細菌對抗生素的抵抗力。李帥[8]報道多重耐藥AB在生物被膜狀態下,相關耐藥基因上調,幫助細菌獲得更強的抗藥性。糖肽類抗生素萬古霉素可抑制細菌細胞壁產生過程中肽聚糖的合成,通過與五肽的D-Ala-D-Ala C末端結合而起作用,從而阻止亞基向肽聚糖骨架的添加[25]。與生物被膜態細菌相比,游離態細菌在萬古霉素耐藥通路下調,可能表明生物被膜的形成增強了細菌對萬古霉素的耐藥性。陽離子抗菌肽信號通路在宿主防御細菌感染中起著重要作用,并且是先天免疫反應的關鍵組成部分[26]。游離態與生物被膜態細菌的轉換過程中陽離子抗菌肽耐藥通路發生了上調。群體感應系統允許細菌感知細胞密度并調節基因表達,群體感應控制的過程包括細菌毒力、運動、孢子形成及生物被膜的形成等[27]。本研究中RNA伴侶蛋白、DNA結合應答調節因子PmrA、雙組份傳感器組氨酸激酶等DEG富集到群體感應信號通路中,表明生物被膜的形成與群體感應密切相關。
綜上,本研究對臨床CRAB生物被膜態細菌和游離態細菌的轉錄組進行比較分析,獲得大量差異基因;這些DEGs富集到硫代謝、ABC轉運及苯甲酸酯降解等通路中,初步揭示臨床CRAB曼不動桿生物被膜形成的機理,為藥物作用靶點的研究及臨床抗感染治療提供一定的理論依據。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
科學大眾(2020年12期)2020-08-13 03:22:22
航空世界(2020年10期)2020-01-19 14:36:20
