黑水虻抗菌肽的研究進展
2021-12-13 03:17:06李瀟瀟侯梅芳
天然產物研究與開發 2021年11期
王 青,陳 俊,李瀟瀟,侯梅芳
上海應用技術大學,上海 201418
抗菌肽(antimicrobial peptides,AMPs)是昆蟲在受到微生物感染或意外傷害時,由脂肪體內合成并釋放到血淋巴中的一類小分子活性多肽,在體液免疫防御中發揮了重要作用。AMPs具有強堿性、熱穩定性和廣譜抗菌活性,并對真菌、病毒、寄生蟲及癌細胞也具有一定的殺滅作用[1-2]。由于AMPs具有廣泛的生物學活性,尤其是對某些耐藥性病原菌也有一定的殺滅作用,因此被認為是理想的抗生素替代物,在畜牧業、食品醫藥和化妝品等領域都具有巨大應用價值和市場潛力[3]。
黑水虻,英文名black soldier fly,又稱亮斑扁角水虻(HermetiaillucensL.),原產于美洲,目前是全世界廣泛分布的雙翅目水虻科昆蟲,生命周期短,繁殖速度快。其幼蟲主要靠取食腐爛有機物、動物糞便或餐廚垃圾等有機廢棄物獲取營養,并能夠高效轉化為昆蟲蛋白,經黑水虻處理過的糞便可以成為優質有機肥[4]。由于黑水虻幼蟲取食時機體暴露于高濃度有害物中,復雜的環境因子誘使其產生AMPs等免疫活性物質來抵抗病原微生物[5],但黑水虻蟲體中幾乎檢測不出沙門氏菌等病原菌,因此還被作為飼料使用[6]。隨著黑水虻養殖業的蓬勃發展,黑水虻體內強大的免疫系統引起了學術界的關注。
本文主要綜述了近年來包括黑水虻在內的昆蟲抗菌肽的種類、結構特征、抑菌活性及生產應用,分析了國內外在該領域取得的研究進展,為研究黑水虻抗菌肽的構效關系提供了思路,同時也為開發黑水虻抗菌肽產品提供了理論支持。
1 黑水虻抗菌肽的種類及結構特征
1974年,研究人員首次在接種了細菌的巨型蠶蛾蛹中觀察到昆蟲的抗菌活性,6年之后第一個昆蟲AMP(cecropin)從盲蝽蛹中純化出來,隨后幾十年的研究已經鑒定出多類型的AMPs[2],在AMPs數據庫APD(http://aps.unmc.edu/AP)列出的3 087種AMPs中有305種來自昆蟲[7]。昆蟲抗菌肽是帶有正電荷且具有兩親性的陽離子多肽,一般由幾十個氨基酸殘基組成,其空間結構為線性或帶環狀[8],二級結構主要由α-螺旋和β-折疊組成。根據其氨基酸序列和結構特征大致可以分為三類,分別為無半胱氨酸殘基的線性α-螺旋肽(天蠶素類,cecropin and moricin),具有6~8個保守半胱氨酸殘基、由3或4個二硫鍵橋的β-折疊結構多肽(防御素類,defensinh和drosomycin),富含脯氨酸和甘氨酸等特定氨基酸殘基的抗菌肽(apidaecin、drosocin、lebocin、attacin和gloverin)[9]。在昆蟲抗菌肽中,天蠶素、防御素、脯氨酸肽和攻擊素是比較普遍的[2],常見的雙翅目昆蟲抗菌肽的名稱和其抑菌范圍見表1。
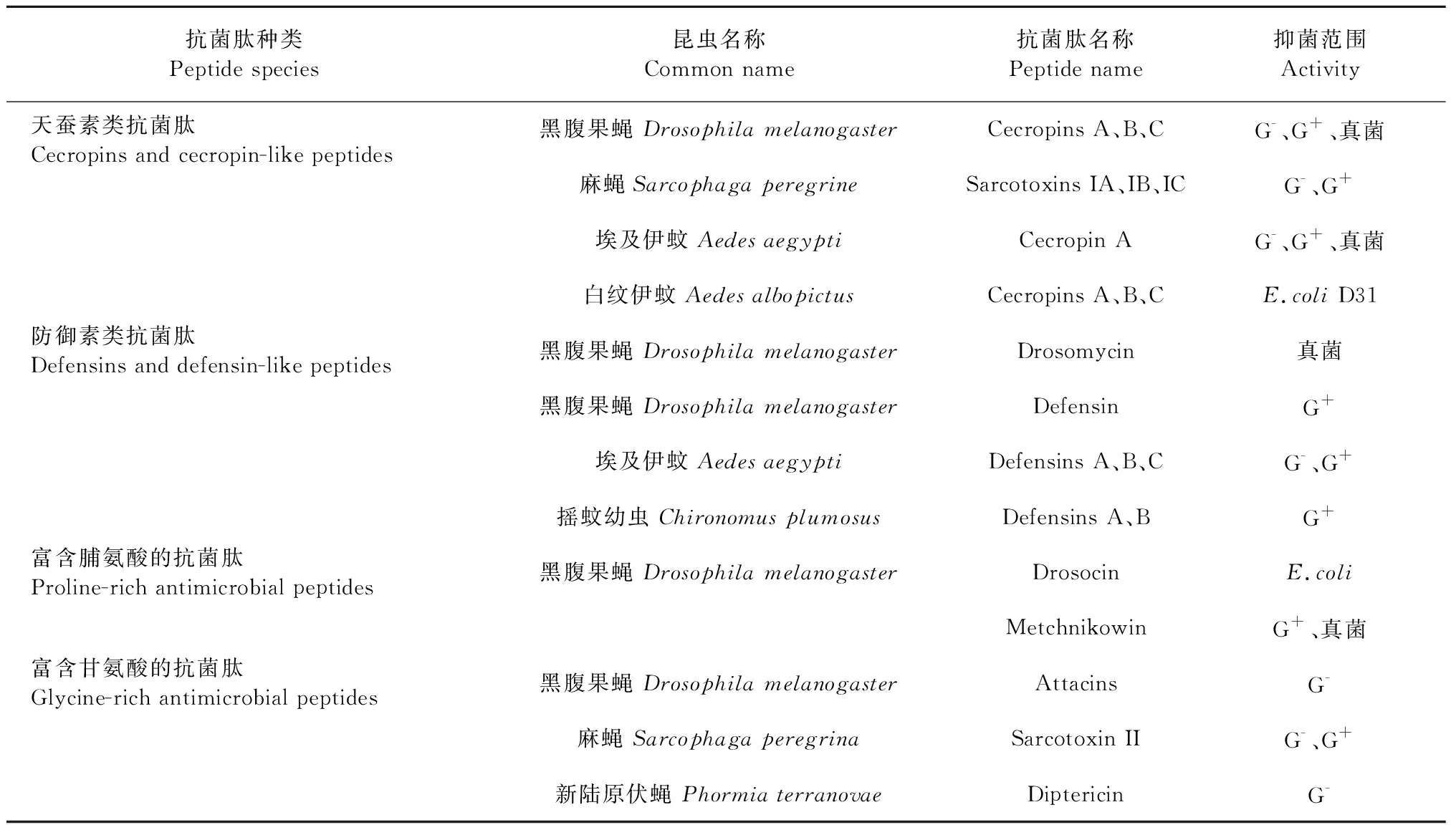
表1 雙翅目昆蟲抗菌肽的種類和抑菌作用[8]Table 1 Species and antimicrobial activity of antimicrobial peptides from Diptera [8]
近年來,越來越多的學者從事黑水虻幼蟲抗菌肽的研究,并分離出多種AMPs,如天蠶素[10]、防御素[11]和攻擊素[12],并驗證了其抗菌活性。
1.1 天蠶素類抗菌肽
天蠶素最初是從大蠶蛾(cecropia moth)的血淋巴中分離出來的[13],是一類陽離子AMPs,一般含有31~39個氨基酸殘基,不含半胱氨酸,所以不能形成分子內二硫鍵,大多數天蠶素在C端被酰胺化,酰胺化能促進天蠶素與脂質體的相互作用,酰胺化對其抗菌活性也具有重要作用[2]。其N端區域具有強堿性,且多含一個色氨酸殘基,可形成雙親螺旋結構[8]。Park等[10]從黑水虻幼蟲血淋巴中提取了天蠶素樣肽(CLP1),純化測定其分子量為4 840 Da。在NCBI BLAST比對中,發現CLP1的氨基酸序列與果蠅天蠶素C的同源性為60%,其特征為陽離子、線性、α-螺旋和兩親性多肽,并對革蘭氏陰性菌有抑菌活性。Park等進一步采用RACE-PCR方法對N-末端氨基酸測序鑒定的新型CLPs的cDNA序列進行分析,獲得了3個CLP基因(CLP1~3),將CLP1~3成熟肽的氨基酸序列與報道的雙翅目昆蟲天蠶素氨基酸序列比較顯示,相似程度為60%~65%(見圖1)。

圖1 黑水虻天蠶素CLP1~3氨基酸序列與雙翅目昆蟲天蠶素的比對[10]Fig.1 Multiple alignment of amino acid sequences of H.illucens CLP1-3 with dipteran cecropins [10]注:*相同氨基酸;黑色:保守的氨基酸序列,下同。Note:*Consensus amino acids;Black box:Conserved amino acids,the same below.
1.2 防御素類抗菌肽
此類AMPs主要有防御素(defensins)和果蠅毒素(drosomycins)[8],昆蟲防御素一般由29~34個氨基酸組成的富含二硫鍵的陽離子型多肽,氨基酸序列中都含有6個保守半胱氨酸和3個分子內二硫鍵,其中2個二硫鍵連接C端β-片層和α-螺旋,第3個二硫鍵連接N端和第二個β-片層形成回路,即單個α-螺旋和兩個反向平行的β-片層結構域,構成穩定的發夾狀結構[14]。Park等[11]通過注射金黃色葡萄球菌誘導黑水虻幼蟲產生抗菌肽,提取血淋巴并使用固相萃取和反相色譜法純化得到新型防御素樣肽(defense-like peptide 4,DLP4),分子量為4 267 Da,DLP4對革蘭氏陽性菌表現出較強的抗菌活性,對革蘭氏陰性菌沒有活性,新型防御素(DLP)是第一個從黑水虻體內分離出來的抗菌肽。為了分析N端氨基酸序列中新發現的DLP的cDNA序列,Park等采用RACE-PCR方法對黑水虻的AMP基因進行全序列分析,得到4個不同的DLP基因(DLP1-4),通過DLP1-3和DLP4相比較,ORF中成熟肽和堿性氨基酸數量存在顯著性差異,該氨基酸被認為是AMP抗菌活性的關鍵氨基酸。進一步將DLP1-4成熟肽的氨基酸序列與報道的雙翅目昆蟲防御素氨基酸序列比較表明,相似程度為72.5%~75%(見圖2),特別是半胱氨酸的位置與雙翅目昆蟲是相同的,N端回路、α-螺旋、β鏈的氨基酸的序列是相當保守的。

圖2 黑水虻防御素DLP1-4的氨基酸序列與雙翅目昆蟲防御素比對[11]Fig.2 Multiple alignment of amino acid sequences of H.illucens DLP1-4 with dipteran defensins [11]
1.3 富含特殊氨基酸的抗菌肽
此類AMPs一般指富含脯氨酸和甘氨酸的抗菌肽,富含脯氨酸的抗菌肽主要包括蜜蜂抗菌肽(apidaecins),果蠅素(drosocins)和家蠶抗菌肽(lebocins)等,由14~39個氨基酸殘基,根據分子大小可分為兩個亞族:短鏈亞族(小于20個氨基酸殘基)和長鏈亞族(大于20個氨基酸殘基)[15],其中一個是高度保守的結構域,具有抗菌活性;另一個結構域有更多的變量,起到特異性靶向的作用[8],并且對革蘭氏陰性菌有較強的抗菌活性[16]。此外,富含甘氨酸的抗菌肽,分子量大小在8~30 kDa,含14%~22%的甘氨酸殘基,具有O-糖基化,如攻擊素(attacins)、雙翅肽(diptericins)、肉蠅毒素(sarcotoxins Ⅱ)、天蠶葛老素(gloverins)等都屬于這一類[17,18]。Shin等[12]通過對黑水虻免疫脂肪體制備的cDNA文庫進行篩選,獲得編碼類攻擊素(attacin)蛋白的cDNA并進行克隆得到新型黑水虻攻擊素(HI-attacin),預測由169個氨基酸組成,分子量為17.7 kDa。HI-attacin對大腸桿菌和耐甲氧西林金黃色葡萄球菌(MRSA)均具有抗菌活性。與其他雙翅類attacins相比,HI-attacin的N端G1結構域短于其它雙翅目昆蟲,含46個氨基酸;而C端G2結構域較G1區保守,包含120個氨基酸(見圖3)。

圖3 黑水虻攻擊素HI-attacin的氨基酸序列與雙翅目昆蟲攻擊素比對[12]Fig.3 Multiple alignment of amino acid sequences of H.illucens attacin with dipteran defensins [12]
2 作用機制
AMPs作用機制是非常復雜的,可以通過多種機制殺死細菌,包括膜破壞,干擾細菌代謝以及靶向細胞質成分[19]。關于AMPs作用機制主要有離子通道學說、抑制細胞呼吸學說、抑制細菌細胞壁的形成學說、胞內損傷假說等幾種觀點來解釋[20]。不同AMPs的作用機制可能不一致,尚有待進一步研究。
2.1 作用于細胞膜機制
由于AMPs帶陽離子,Ongey等[21]認為AMPs可通過靜電或疏水與微生物陰離子表面的相互作用和插入磷脂雙分子層發揮作用,以改變膜的通透性。AMPs可以在細菌細胞質膜上穿孔而形成離子孔道,造成細菌細胞膜結構破壞引起胞內水溶性物質大量滲出,而最終導致細菌死亡。因此,AMPs與膜之間的相互作用與AMPs的抗菌活性直接相關。目前,主要有四種作用模型用于描述AMPs作用于細胞膜:桶形壁模型、地毯模型、環形孔模型和凝聚模型[22](圖4)。例如,天蠶素和天蠶素樣肽(cecropins)對G-抗菌活性要強于G+,它與細胞膜相接觸是以其長軸α-螺旋結構域平行于脂質雙分子層表面,極性殘基與磷脂相互作用,非極性殘基埋入膜的疏水核心。在高濃度時,cecropins形成地毯狀結構破壞細胞膜,具有類似洗滌劑的特性。在低濃度時,cecropins形成孔隙影響細胞電解質平衡,并最終導致細菌死亡[7]。

圖4 抗菌肽作用細胞膜機制模型[22]Fig.4 Mechanism model of antibacterial peptide acting on cell membrane [22] 注:A.毯式模型;B.桶板模型;C.環孔模型;D.聚集模型。Note:A.The carpet model;B.The barrel-stave model;C.The toroidal pore model;D.The aggregate model.
2.2 細胞內殺傷機制
富含脯氨酸的抗菌肽(pyrrhocoricin、drosocin和apidaecin)與其他類型的抗菌肽不同,主要是抗G-。其作用方式不涉及膜的溶解,而是通過進入易感細胞在細胞內部產生功效。通過定位于細胞內相應位點阻礙或抑制細胞組分的合成,從而對細胞造成殺傷作用[23]。AMPs在細胞內部的靶點主要是70 kDa熱休克蛋白DnaK[3]。它們還可以通過與核酸結合而干擾DNA和RNA的合成[24](圖5),這一特性使它們成為一種新型潛在的細胞穿透肽。
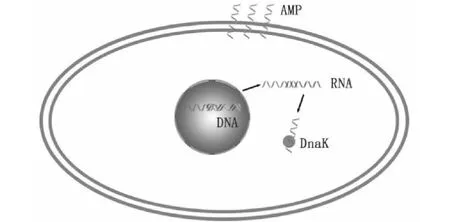
圖5 抗菌肽細胞內殺傷機制模型[23]Fig.5 Model of intracellular killing mechanism of antimicrobial peptides [23]
3 黑水虻抗菌肽的生產制備
目前AMPs主要通過3種途徑獲取,一是直接從昆蟲體內分離純化;二是化學合成;三是生物工程法[25]。根據近幾年的研究報道,黑水虻抗菌肽主要采用從蟲體直接分離提純和異源表達。Park等[26]最初研究黑水虻抗菌肽時,直接將黑水虻幼蟲凍干粉用甲醇萃取,再使用高效液相色譜法從幼蟲提取物中分離出具有廣譜抗菌活性的黑水虻抗菌肽并用Edman降解法鑒定防御素的氨基酸序列。由于直接提取分離得到的AMPs產量不高,而化學合成法的提取工藝復雜,且成本高、耗時長,因此這兩種途徑都不足以達到規模化工業生產,從而限制了AMPs的產業化應用[18]。近年來,國內外研究人員開始利用基因工程的方法生產制備黑水虻抗菌肽,通過AMPs基因克隆到細菌和真菌載體進行重組融合表達[27]。重組表達制備AMPs的工藝在不斷優化,但仍遠高于傳統藥物的制造成本。
3.1 黑水虻抗菌肽的重組表達
3.1.1 大腸桿菌表達系統生產抗菌肽
從20世紀80年代開始,人們就運用大腸桿菌表達系統表達AMPs,具有遺傳背景清楚、成本低、表達量高和表達產物分離純化相對簡單等優點。在所有基因工程菌生產AMPs的方法中,大腸桿菌表達系統是最常見的。但是AMPs對宿主細胞的毒性抑制了其在大腸桿菌體內的表達量,因此多采用融合蛋白方式來降低昆蟲抗菌肽對大腸桿菌菌株的毒性,表達后再經化學裂解或者酶解處理后得到有抗菌活性的表達產物[20]。Elhag等[28]從黑水虻中分離出189 bp新型AMPs基因stomoxynZH1,利用PCR技術克隆出AMPs基因并連接到表達載體pET32a,然后轉入大腸桿菌BL21(DE3)中表達,結果表明基因工程菌株的表達產物重組肽(Trx-stomoxynZH1)對大腸桿菌和金黃色葡萄球菌均有抑制作用。
3.1.2 枯草芽孢桿菌表達系統生產抗菌肽
枯草芽孢桿菌是一種革蘭氏陽性細菌,具有非致病性和分泌蛋白能力強的特性,是目前原核表達系統中表達和分泌外源蛋白的理想宿主,成為原核表達系統中的一種重要的模式菌株[29]。Xu[30]利用大腸桿菌-枯草芽孢桿菌穿梭載體PMA5質粒和PAX01質粒構建整合型大腸桿菌-枯草芽孢桿菌穿梭載體質粒PAX01-KANA,成功構建含有黑水虻幼蟲抗菌肽 DLP4、沙蛾抗菌肽SMFP和天蠶素融合基因Cecropin AD三段抗菌肽基因的枯草芽孢桿菌工程菌。結果顯示,工程菌株發酵液對大腸桿菌有明顯的抑菌活性,工程菌在LB 液體培養基中生長良好。Ji等[31]將CAM-W、EDDIE和信號肽SacB融合基因轉移到由啟動子Pg/v控制的pDM03載體中,轉化到枯草芽孢桿菌內,形成高效表達的重組枯草芽孢桿菌WB700表達系統,在上清發酵液中Cecropin A-melittin濃度高達159 mg/L。
以上兩種表達系統是目前基因工程領域研究應用最廣泛、遺傳表達圖譜最清楚的原核表達系統。雖然大腸桿菌表達系統應用較多,但表達的蛋白質以包含體形式存在,與大腸桿菌內毒素不易分離,缺少蛋白質翻譯后修飾和加工過程,且雜蛋白多,純化步驟復雜[30]。枯草芽孢桿菌的表達系統研究雖然起步較晚,但其本身具有一些獨特的優勢:安全無毒、較強的蛋白分泌能力和發酵工藝成熟[29],因此也受到研究人員的關注。此外,還有真核表達系統和桿狀病毒表達系統等,隨著研究的不斷深入有望成為滿足黑水虻抗菌肽工業化生產需求的表達系統[20]。
4 黑水虻抗菌肽的應用前景
昆蟲抗菌肽具有抗菌活性、抗病毒活性、抗寄生蟲活性、抗腫瘤細胞活性和抗炎活性等生物學活性[8],因此可以應用在農牧業、醫藥、食品、化妝品以及保健品等領域。目前,黑水虻幼蟲主要集中用于畜禽糞便等有機垃圾的無公害處理[32],研究人員也在加強黑水虻抗菌肽及其他系列產品的開發。
4.1 在飼料添加劑領域的應用
黑水虻含有較豐富的粗蛋白質且氨基酸種類較齊全,不飽和脂肪酸、礦物質和類胡蘿卜素等,還能提取幾丁質、AMPs等生物活性物質,是一種優質蛋白質飼料原料[33]。Yu等[34]研究發現,添加黑水虻幼蟲可以提高豬肉的品質。黑水虻幼蟲抗菌肽可以作為飼料添加劑提高畜禽免疫力,提高畜禽生長性能。Ren等[35]報道,在飼料中添加500 mg/kg的AMPs可顯著提高斷奶仔豬血液中免疫球蛋白IgG、IgM、IgA與血清總補體CH50等免疫因子的含量。Dong等[36]研究在飼養鯉魚時添加AMPs,可以降低血清中的甘油三酸酯水平,增強抗氧化性并提高鯉魚的免疫力。結果表明,適當濃度的AMPs作為鯉魚飼料中的補充劑可以提高生長性能。由于AMPs擁有良好的理化特性,所以在飼料加工過程中也不會丟失活性,還可以提高飼料的品質和延長保質期。
4.2 在生物醫藥領域的應用
目前很多企業已在AMPs藥物方面做了大量的研究,并有一些含AMPs成分的產品上市。由于抗生素濫用導致多重耐藥菌的產生,因此AMPs被認為是理想的抗生素替代品[27]。此外,AMPs的抗癌效果也被國內外學者所關注。Lee等[37]分離出黑水虻幼蟲的血淋巴中的抗菌肽HP/F9,證實該肽在體外有效抑制肺炎克雷伯菌的生長,并有效保護小鼠免受肺炎克雷伯菌的感染,這項研究為HP/F9可用作新型抗菌物質,用于治療細菌感染。夏嬙課題組對黑水虻幼蟲抗菌肽的醫用價值做了大量研究,發現其可以抑制鼻喉癌腫瘤細胞生長[38]。目前已有大量不同治療目的肽類藥物進入臨床或批準上市,如GRN-1201、WT-2725、ITK-1等[39]。昆蟲的抗菌肽的藥用價值在我國中醫藥中很早就有記載,如將家蠅幼蟲洗凈曬干后與其他中藥配制,用于治療感染性疾病[40]。
4.3 在食品安全領域的應用
食品安全中最主要的問題就是微生物污染和化學防腐劑的添加,AMPs作為天然的防腐劑,具有廣闊的應用前景[41]。AMPs在食品安全領域的應用包括兩個方面,一是在食品原料控制環節應用,減少或替代抗生素的使用,提高食品加工原料的安全品質;二是在食品防腐保鮮環節,作為高效、安全生物防腐劑應用于食品加工、儲運等諸多環節,解決食品防腐劑的殘留問題,同時也減少環境耐藥性菌株的產生,進而改善食品安全環境條件[42]。Secci等[43]用黑水虻幼蟲粉替代大豆粉飼養母雞,雞蛋的質量明顯提高,保障了食品原材料的安全。Choi等[44]從黑水虻幼蟲中提取到抗菌肽ME,對導致食品污染的宋內志賀氏菌有較強的抑制活性。此外,不同種類的黑水虻抗菌肽還對導致食源性污染的大腸桿菌[10]、金黃色葡萄球菌[11]也有抑制作用。
4.4 在化妝品添加劑領域的應用
AMPs由于其獨特的抗菌、無毒和無刺激性等優點,也被用于化妝品生產。昆蟲抗菌肽不僅可作為營養成分,更主要的是其抗菌和美容作用具有獨特的優勢和廣泛的市場潛力。Tonk等[45]提出將具有抗皮膚癌效果的AMPs和個人護理產品,可以在皮膚癌在癥狀變得明顯之前通過消除新生癌細胞來預防和增強人體健康。已證實多種抗菌肽可以對抗痤瘡丙酸桿菌而具有治療痤瘡的潛能[46,47]。黑水虻抗菌肽CLP1對銅綠假單胞菌有抑制作用[10],再加上AMPs的抗炎作用[48],也為開發黑水虻抗菌肽消炎祛痘的化妝品提供了新的思路。
5 總結
黑水虻作為一種新型“資源昆蟲”,近年來備受國內外學者關注。黑水虻體內可生產抗菌肽等具有高附加值的產品,對黑水虻養殖業帶來了新的發展。黑水虻抗菌肽不僅具有廣譜抗菌效果,還具備抗病毒、抗寄生蟲、抗腫瘤細胞和抗炎等活性,與傳統抗生素相比有很大的優勢,在飼料、食品、醫藥與化妝品領域具有廣泛的應用前景,成為了目前生物活性肽的研究熱點之一。但黑水虻抗菌肽天然提取難度較大,化學合成成本過高,基因工程表達表達量偏低,其制備技術制約了其大規模的生產,此外,黑水虻抗菌肽體內活性機制也有待進一步研究明確。隨著基因工程技術和蛋白質組學技術的進步,我們相信黑水虻抗菌肽在不遠的將來會實現產業化發展,并帶來更大的經濟效益和社會效益。
