霍山石斛菌根真菌多樣性及差異性的研究
2021-12-13 03:16:56陳紹通劉旭宇韓邦興
天然產物研究與開發 2021年11期
劉 麗,陳紹通,張 騰,李 琳,劉旭宇,韓邦興,4*
1安徽中醫藥大學藥學院,合肥 230031;2皖西學院生物與制藥工程學院,六安 237012;3華南農業大學,廣州 510000;4安徽省中藥資源保護與持續利用工程實驗室,六安 237012
霍山石斛(DendrobiumhuoshanenseC.Z.Tang et S.J.Cheng)來源于蘭科石斛屬,為多年生草本植物,具有增強人體免疫力、抗氧化、修復肝損傷、制止心血管疾病發生等功效[1-3]。霍山石斛不僅有著極大的保健醫療價值,而且食用歷史悠久;近幾年,隨著越來越多的人開始關注保健產品,其市場需求量不斷增加[4]。霍山石斛常以三年生入藥,不同生長年限的霍山石斛,其藥材質量也有所差異。石斛屬植物的生長形式是合軸式生長,主軸生長到一定階段就會停止生長,開始發出側芽形成新軸,同樣到一定階段后,新軸上繼續發出側芽又形成新軸,依此往復,植物得以不斷生長。按照發芽時間的前后將霍山石斛分為三年莖(ⅢY3)、二年莖(ⅢY2)、一年莖(ⅢY1),分別對應根ⅢR3、ⅢR2和ⅢR1;同理,二年生的霍山石斛可分為一年莖(ⅡY1),二年莖(ⅡY2),各自對應根ⅡR1和ⅡR2,一年生的霍山石斛為ⅠY1,對應根ⅠR1[5]。研究發現,微生物對蘭科植物的影響不僅關乎種子萌發[6],還能促進生長發育[7]及成分積累等[8]。
迄今為止對石斛菌根真菌的研究一般采用傳統培養分離法,已報道的有半知菌門(Deuteromycotina)的絲核菌屬(Rhizoctonia)、鏈格孢屬(Alternaria)[9,10],子囊菌門(Ascomycota)的毛殼菌屬(Chaetomium)、炭角菌屬(Xylaria)、刺盤孢屬(Colletotrichum)、鐮刀菌屬(Fusarium)[9,11],擔子菌門(Basidiomycota)的膠膜菌屬(Tulasnella)[9,12]、小菇屬(Mycena)、蠟殼菌屬(Sebacina)和角擔菌屬(Ceratobasidium)[9,13]等。向石斛屬植物接種菌根真菌不僅能夠促進植株生長,還具有不同程度的抗旱、抗病以及促進石斛堿和多糖的積累等作用[14-16]。雖然近幾年利用傳統培養分離方法對石斛屬植物內生菌的研究較多,但因為微生物培養具有很大的隨機性和盲目性,99%的微生物無法分離得到[17],目前關于一年生霍山石斛菌根真菌的研究尚未見報道。本文運用高通量測序技術以三種類型一年生霍山石斛菌根真菌為研究對象,目的是能夠系統了解其群落多樣性、差異性及結構組成,為指導霍山石斛合理采收及應用提供依據。
1 材料與方法
1.1 樣品采集
每隔兩個月于皖西學院藥用植物園設施栽培基地采集三種一年生類型(ⅠR1、ⅡR1和ⅢR1)霍山石斛樣品,方法為五點隨機取樣法,樣品經皖西學院韓邦興教授鑒定。本實驗以1年生霍山石斛的根為研究對象(從不同生長年限的植物叢中分得),ⅢR1取自ⅢY1莖下對應的根,ⅡR1取自ⅡY1莖下對應的根,ⅠR1取自ⅠY1莖下對應的根。具體測序樣品信息見表1。
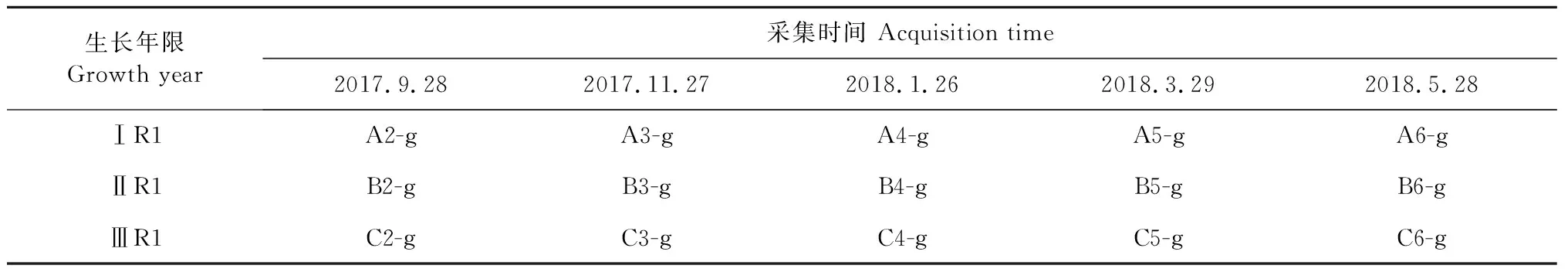
表1 三種類型一年生霍山石斛樣品Table 1 Three types of annual D.huoshanense samples
1.2 方法
1.2.1 樣品消毒處理
將采集的霍山石斛植株用無菌剪刀剪取根,用自來水沖洗干凈,置于超凈工作臺內晾干,樣品消毒:置于0.1%升汞中浸泡2~3 min,無菌水漂洗3~4次,再置于75%的乙醇中浸泡3 min,無菌水漂洗5~6次,最后用干燥無菌濾紙擦去表面附著的水分,并將須根兩端用無菌剪刀除去,保留中間段。將最后一次沖洗的無菌水均勻涂布于PDA培養基上,28 ℃黑暗培養3~7天,觀察是否有菌長出,以此驗證表面消毒的有效性。
1.2.2 DNA提取、PCR擴增及測序分析
將上述消毒完全的樣品,按照OMEGA試劑盒E.Z.N.ATM Mag-Bind Soil DNA Kit說明書的步驟進行DNA提取,并用瓊脂糖凝膠檢查DNA的提取效果。真菌ITS1-2擴增區域采用通用引物ITS1F和ITS2R進行擴增。反應體系見表2,擴增程序見表3。霍山石斛樣品DNA檢測合格之后,由上海生工生物工程股份有限公司利用Illumina MiSeqTM平臺進行測序。

表2 反應體系Table 2 Reaction system
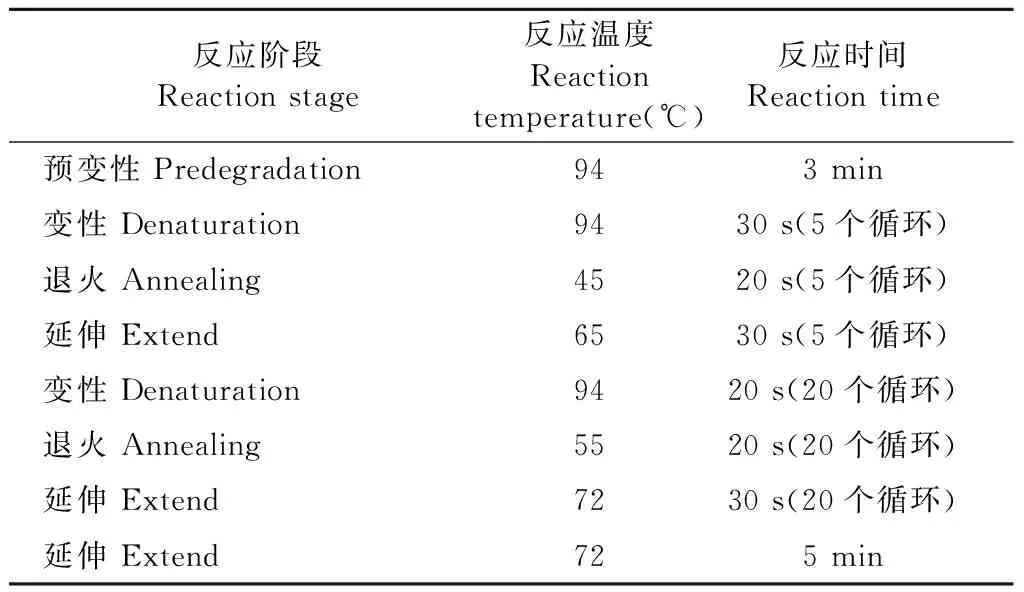
表3 擴增程序Table 3 Amplification procedure
1.2.3 數據處理及分析
利用QIIME1.8.1對所得到的基因序列進行拼接、過濾、去除嵌合體及非特異性擴增序列,得到各樣本高質量序列[18]。將高質量的序列與數據庫中代表性序列進行比對。通過Usearch將所有樣本序列根據序列間的距離進行聚類,然后按照97%的相似性將序列分成不同的操作分類單元(OTU),在此基礎上,選擇豐度最高的序列作為OTU的代表性序列,而后進行各類分析[19]。使用R的Venn Diagram包做出Venn圖,以此統計樣本中共有及特有OTU的數量;通過Mothur計算Alpha多樣性指數,如Shannon指數、Simpson指數、Chao1指數及覆蓋率(coverage)[20];采用Mothur對相似水平是97%的OTU進行稀釋性分析,檢測樣品的取樣深度。采用R對物種分類學統計結果作圖,計算樣品中所含何種微生物以及各微生物的相對豐度。采用統計學方法進行差異特征發現和顯著性檢驗,LEfSe(linear discriminant analysis effect size,線性判別分析及影響因子)通過將統計顯著性的標準檢驗與編碼生物一致性和效應相關性的附加檢驗相結合,確定最能解釋組間差異的基因[21]。
2 結果
2.1 物種組成(OTU)分析
對真菌ITS1-2區域進行高通量測序,共得到898 663條高質量序列,基于≥97%的相似度水平,通過聚類分析共獲得1 928個有效OTUs(見圖1),其中ⅠR1中有787個OTUs,ⅡR1中有1 018個OTUs,ⅢR1中有903個OTUs,200個OTUs共同存在于三組。ⅠR1中特有389個OTUs,ⅡR1特有516個OTUs,ⅢR1中特有443個OTUs。隨著15個霍山石斛根部樣品的序列數不斷增加,其稀釋性曲線越來越趨于平緩,說明即使繼續增加取樣量也只能產生極少量新的OTU(見圖2),說明取樣量合理,能夠比較準確地反映霍山石斛菌根真菌群落。
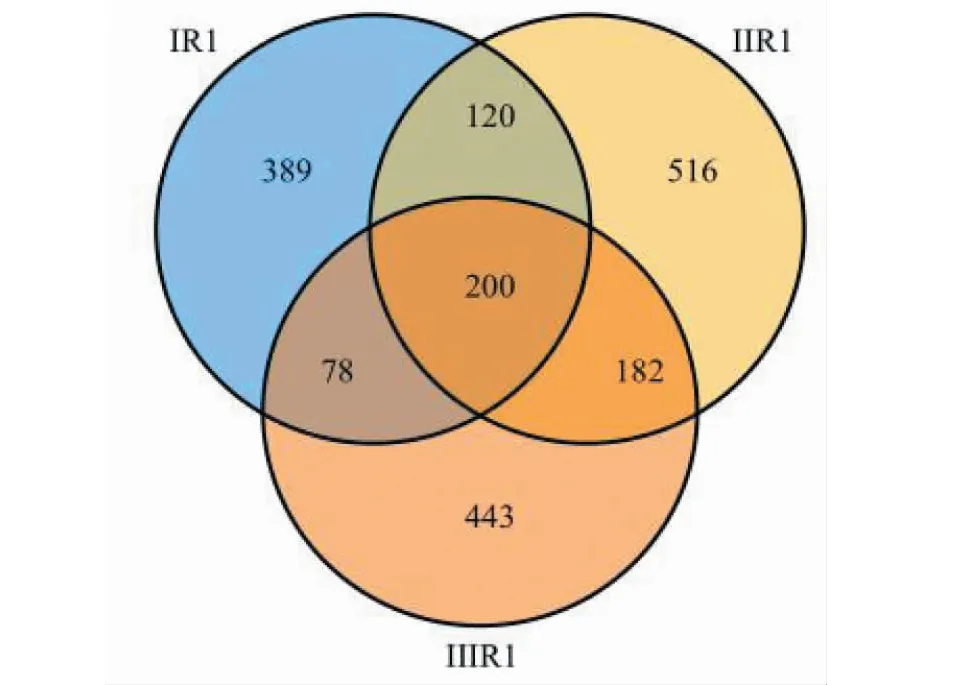
圖1 霍山石斛菌根真菌物種(OTUs)組成Venn圖Fig.1 Venn diagram of species composition of mycorrhizal fungi (OTUs) of D.huoshanense
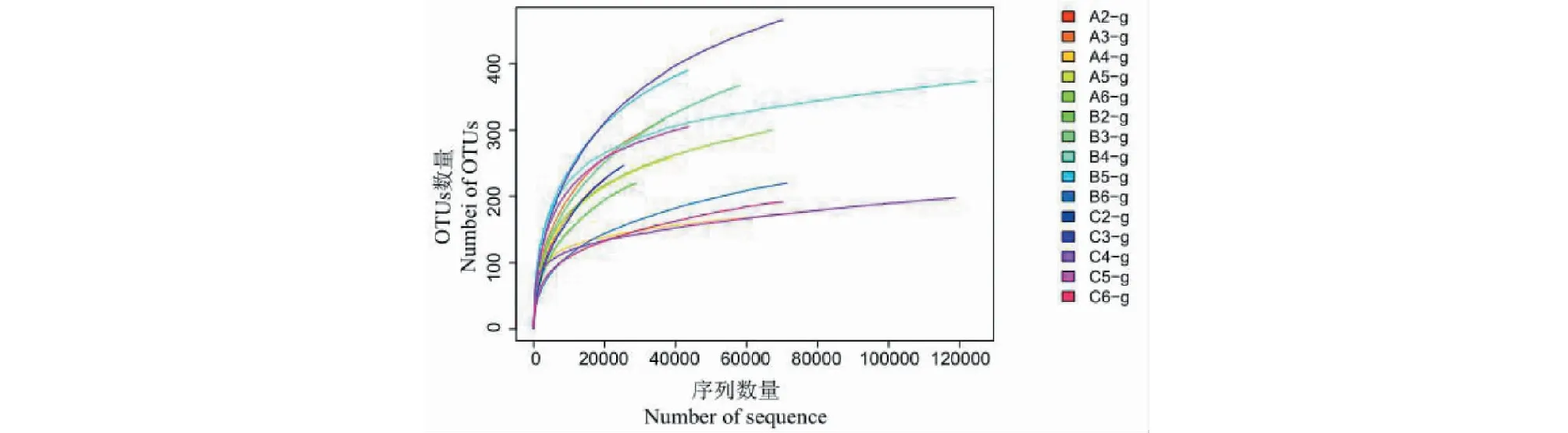
圖2 樣品真菌測序的稀釋曲線Fig.2 Dilution curve of fungal sequencing
2.2 多樣性指數分析
三組樣品的Alpha多樣性指數如表4所示。用Shannon指數量化微生物群落多樣性。ⅡR1、ⅢR1、ⅠR1組的平均Shannon指數分別為2.79、2.74、2.42,IIR1>ⅡIR1>IR1。用Chao1算法評估物種豐富度大小。ⅠR1、ⅡR1、ⅢR1三組群落分布豐度的Chao1指數分別為320.7、394.27、365.81;同樣,ⅡR1的菌根真菌豐富度最大,ⅠR1最小。霍山石斛各樣品的coverage值達到1.00,表明該測序結果能夠代表樣本的真實情況。Alpha多樣性評價箱式圖評估了三組樣本的離散及分布情況(見圖3),Shannon指數的箱式圖表明三組樣品的菌落的均勻度基本一致(P>0.05),Chao1指數的箱式圖表明三組樣品的物種豐富度基本一致(P>0.05)。
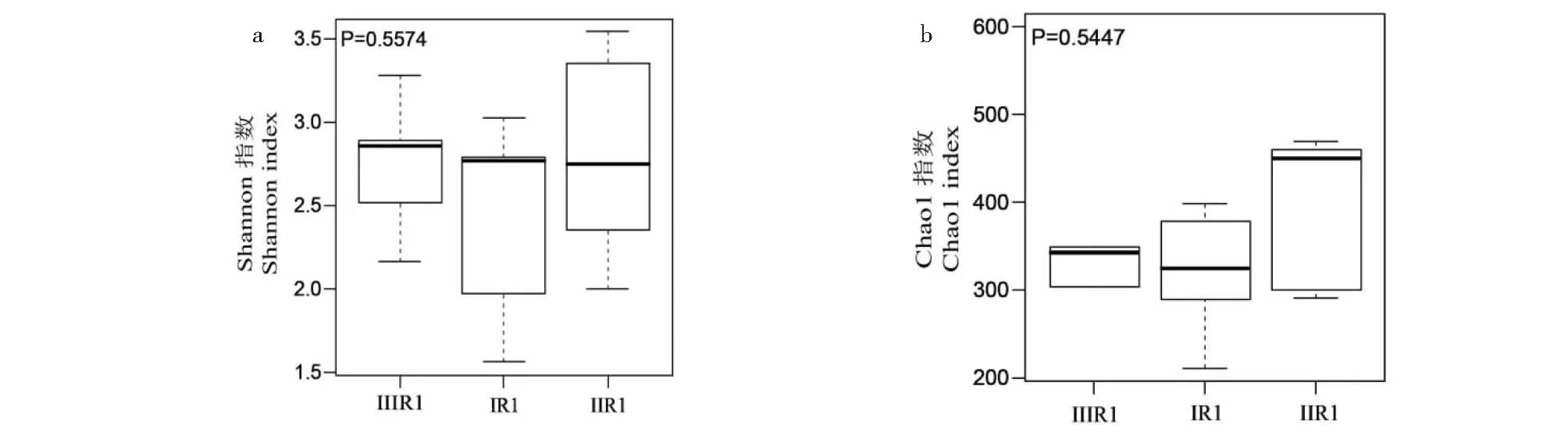
圖3 Alpha多樣性評價菌根真菌菌群Fig.3 Alpha diversity evaluation of mycorrhizal fungi flora注:a:Shannon指數多樣性;b:Chao1指數多樣性。Note:a:Shannon index diversity;b:Chao1 index diversity
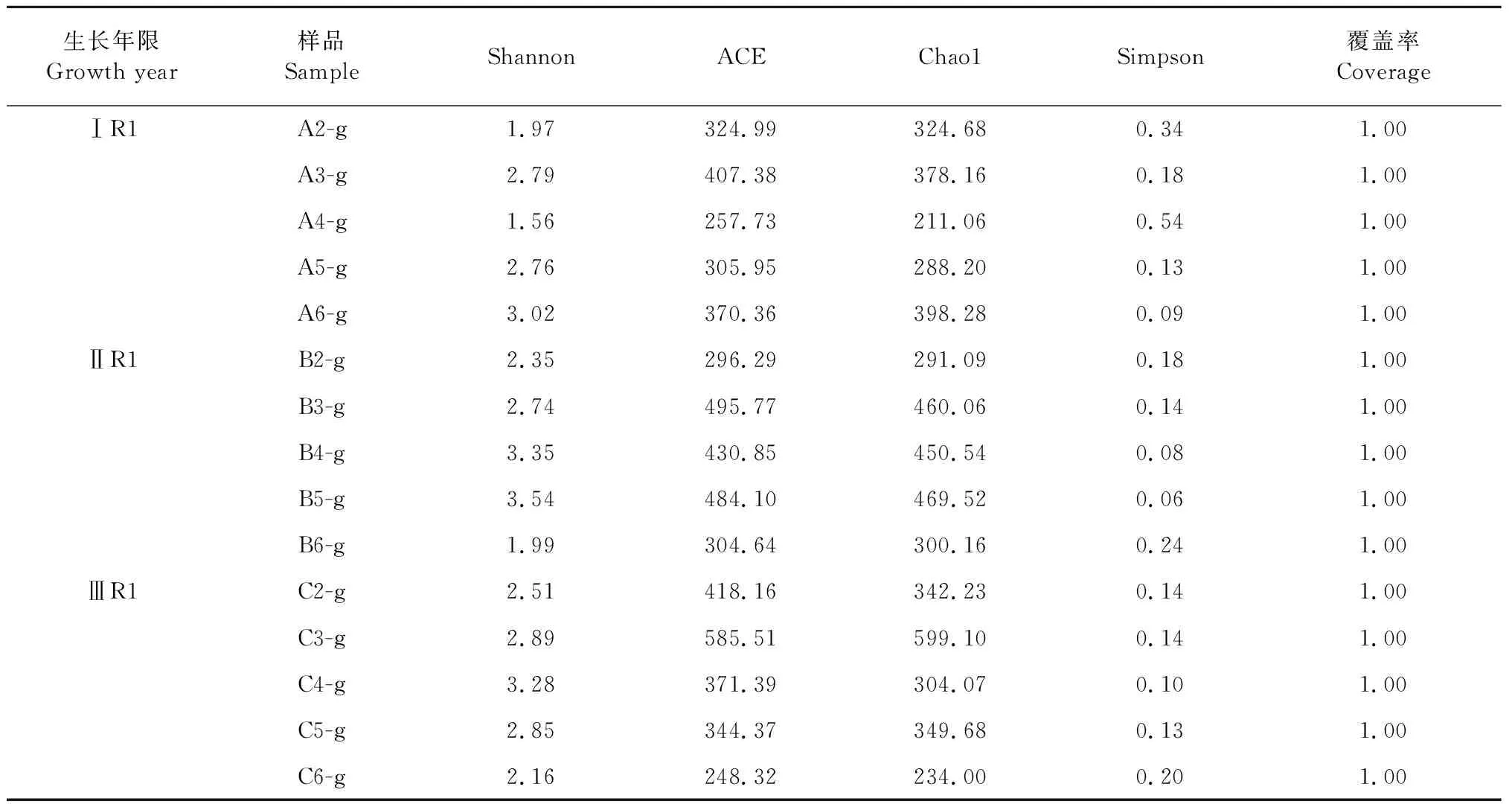
表4 菌根真菌的Alpha多樣性指數表Table 4 Alpha diversity index of mycorrhizal fungi
2.3 群落組成分析
基于OTUs聚類分析得到9個門,30個綱,71個目,149個科,244個屬。在門水平,霍山石斛根真菌主要集中在子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)和毛霉門(Mucoromycota)三個門,占霍山石斛菌根真菌總量的90%左右(見表5);同時5月份的三個樣品A6、B6、C6中子囊菌門的豐度最大,擔子菌門的豐度最小;9月份的兩個樣品A2、C2中擔子菌門的豐度最大,子囊菌門的豐度最小。在屬水平,unclassified Serendipitaceae、unclassified Auriculariales、Zymoseptoria、unclassified Pleosporales、Fusarium、unclassified Agaricomycetes在三種類型一年生霍山石斛菌根真菌中占優勢(見表6)。unclassified Serendipitaceae在ⅠR1中豐度最大(>45%),在ⅢR1中豐度最小(<5%),相反Fusarium在ⅠR1中豐度最小(<5%),在ⅢR1中豐度最大(>20%);Zymoseptoria在ⅡR1中豐度最大(>10%),而在ⅠR1、ⅢR1上均小于5%。
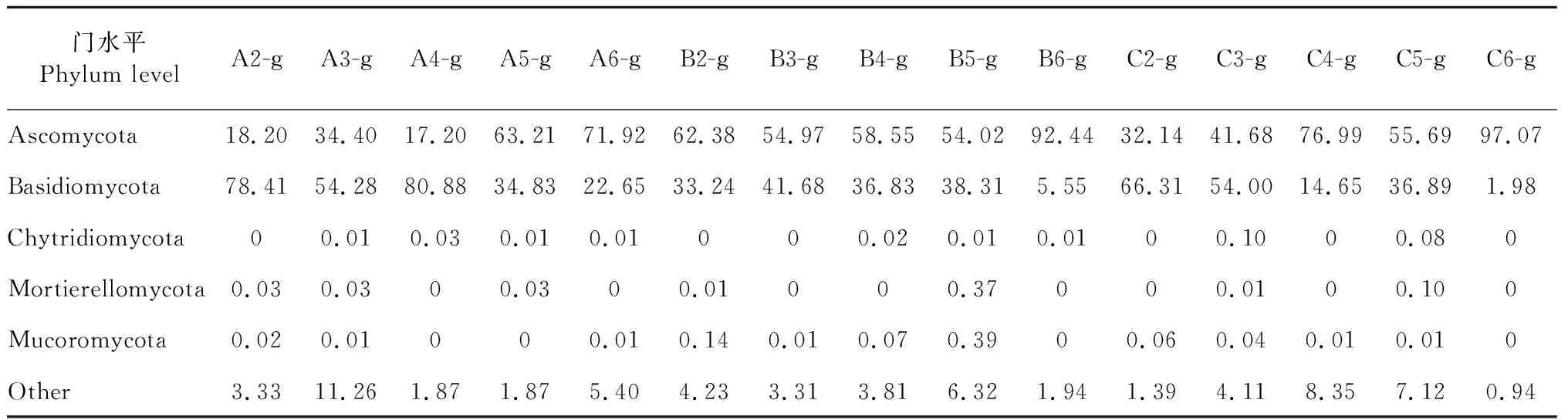
表5 門水平各樣品菌群豐度Table 5 Abundance of fungus in each sample at phylum level (%)
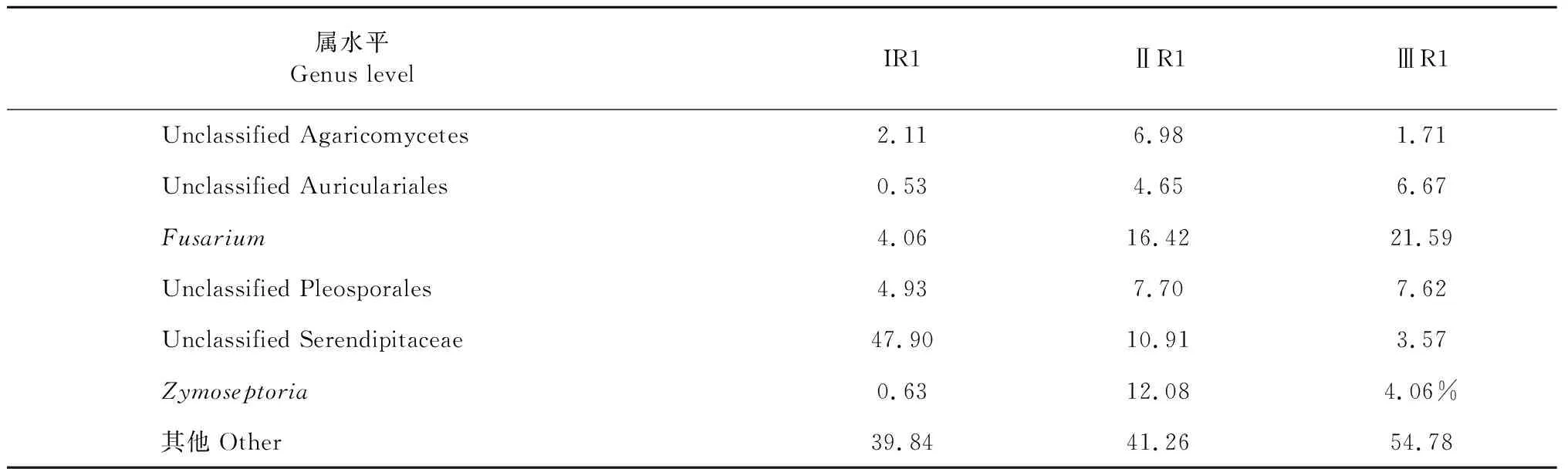
表6 屬水平各組樣品菌群豐度Table 6 Abundance of fungus in each group of the genus level (%)
2.4 菌群差異分析
應用LEfSe對三種類型一年生的霍山石斛樣品的菌根真菌進行分析,發現44個差異豐富的分類學分支(a = 0.01,LDA score≥2.0)(見圖4)。在屬水平(見圖4a),ⅠR1組的擬棘殼孢屬(Pyrenochaetopsis)、青霉菌屬(Penicillium)與其他兩組存在顯著差異,且在該組豐度最高;ⅡR1組刺球菌屬(Chaetosphaeria)、柱霉屬(Scytalidium)、鞘孢屬(Chalara)、Zymoseptoria、紅酵母屬(Rhodotorula)、Kockovaella與其他兩組存在明顯豐度差異;ⅢR1組Zasmidium、unclassified Stachybotryaceae、阿太菌屬(Athelia)、糙孢菌屬(Trechispora)與其他兩組存在明顯豐度差異。
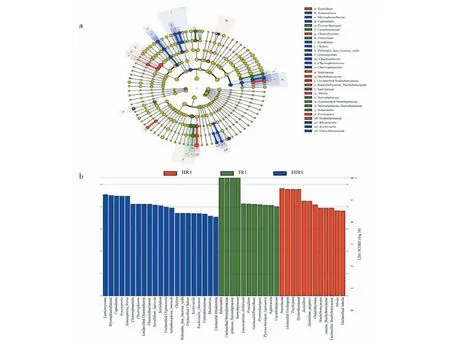
圖4 三種類型一年生霍山石斛菌根真菌的LEfSe結果Fig.4 LEfSe results of three types of mycorrhizal fungi of D.huoshanensis
3 討論與結論
植物根系微生物不僅能夠通過調控基因表達來促進化學成分合成[22],還可以增加次生代謝產物的積累[23]。近年來對石斛屬植物根系微生物研究普遍,如石斛屬植物菌根真菌的特異性與寄主植物系統發育的關系[24],研究發現石斛屬植物鐵皮石斛的根中廣泛分布子囊菌門、擔子菌門真菌[25,26]等。由于微生物培養的方法研究內生菌存在局限性,本文采用高通量測序技術發現霍山石斛根部內生真菌豐富多樣,子囊菌門、擔子菌門為霍山石斛根部內生真菌的主要真菌門,ⅠR1、ⅡR1、ⅢR1有各自的優勢菌,與子囊菌門、擔子菌門在丹參、石斛屬等藥用植物內生真菌中也占主導地位報道基本一致[27,28]。通過Alpha多樣性分析發現三組樣品不管是在物種豐富度還是群落多樣性比較上均無顯著差異。而LEfSe進行組間內生真菌菌群差異分析發現,ⅠR1組有3個科與其他兩組存在顯著差異,且在該組豐度最高;ⅡR1組有5個科與其他兩組存在明顯豐度差異;ⅢR1組有4個科與其他兩組存在明顯豐度差異。
已有研究表明,中藥茅蒼術夏季的菌根內生真菌多樣性高于春季和秋季[29]。本研究也對不同季節的霍山石斛根部內生真菌進行了考察,發現5月份霍山石斛根中子囊菌門的豐度最大,擔子菌門的豐度最小,9月份擔子菌門的豐度最大,子囊菌門的豐度最小,說明季節可能是影響霍山石斛菌根真菌群落結構的重要因素。在后續研究中,擬將石斛根際微生物結合化學成分,重點研究根系微生物對霍山石斛有效成分的影響,為提高霍山石斛品質、開發菌肥提供科學依據等。
綜上所述,本研究應用Illumina MiSeq高通量測序方法對霍山石斛菌根真菌ITS1-2區進行測序,得到898 663條ITS rRNA序列,1 928個OTUs,基于OTU聚類分析得到9個門,30個綱,71個目,149個科,244個屬,其中子囊菌門、擔子菌門和毛霉門為霍山石斛根部內生真菌的主要真菌門,在5月份子囊菌門的豐度最大,擔子菌門的豐度最小;LEfSe分析發現三種類型一年生霍山石斛菌根真菌有44個差異豐富的分類學分支(a = 0.01,LDA score≥2.0);通過Alpha多樣性分析發現無論是物種組成還是多樣性方面,均是ⅡR1>ⅢR1>ⅠR1,但是Alpha多樣性評價箱式圖反映了三組并無顯著差異。
