乙型肝炎病毒對miR-10b表達的影響及其生物信息學分析*
2021-11-20 09:45:54梁俐蘭劉莉莉覃超梅石明連劉永明蘇何玲
重慶醫學 2021年21期
梁俐蘭,劉莉莉,覃超梅,梁 斌,石明連,劉永明,蘇何玲
(桂林醫學院生物化學與分子生物學教研室,廣西桂林 541100)
乙型肝炎病毒(hepatitis B virus,HBV)感染引起的急、慢性肝炎,肝硬化和肝細胞癌(hepatocellular carcinoma,HCC)是全球性的主要衛生問題。全球慢性乙型肝炎患者約2.57億人,每年約887 000人死于HBV感染導致的肝硬化和HCC。近年來,HBV引起的宿主微小RNA(micro RNA,miRNA)表達變化及其在肝臟疾病發生、發展中的作用已成為HBV感染相關疾病發病機制研究的熱點[1-2]。miRNA是一類長度為19~25個核苷酸的非編碼單鏈小分子RNA,可通過與靶基因mRNA的3′非翻譯區部分序列同源性結合而誘導基因沉默,在基因表達調控中發揮著重要作用。有研究表明,miRNA可影響HBV在宿主細胞內的復制和基因表達,而HBV則可通過調節宿主miRNA表達為其提供生存與復制的有利環境[3-6]。這種miRNA介導的HBV與宿主的相互關系是HBV感染相關疾病發病機制的重要基礎。
miR-10b是新近被發現與多種疾病的發生、發展密切相關的一種疾病相關性miRNA,在一些病毒性疾病的發病機制中具有關鍵作用[7-9]。本研究分析美國生物技術信息中心(national Center for Biotechnology Information,NCBI)基因表達綜合數據庫中的GSE19980和GSE121248數據發現,miR-10b在穩定表達HBV的HepG2.2.15細胞中表達明顯上調。進而提取HepG2.2.15細胞及其對照HepG2細胞的總RNA進行熒光定量聚合酶鏈反應(polymerase chain reaction,PCR)檢測,證實了HBV對miR-10b表達的上調作用。在此基礎上,對HBV上調miR-10b表達在肝臟疾病中的作用進行生物信息學分析,以期為研究miR-10b在HBV感染及其致病機制中的作用提供理論與實驗依據。
1 材料與方法
1.1 實驗材料
人肝癌細胞HepG2細胞株及穩定表達HBV的HepG2.2.15細胞均購自上海中國科學院細胞庫。胰蛋白酶、高糖杜氏改良Eagle培養基(Dulbecco′s modified Eagle medium,DMEM)、青霉素鏈霉素及胎牛血清均購自美國Gibco公司;RNA提取試劑TRI Reagent購自美國Sigma公司;All-in-OneTMmiRNA qRT-PCR Detection試劑盒購自美國GeneCopoeia公司,反轉錄引物為試劑盒中的Oligo-dT adaptor primer, PCR反應所用特異性引物由上海生工生物工程公司合成。普通PCR儀及熒光定量PCR儀購自美國BIO-RAD公司。
1.2 方法
1.2.1miRNA芯片GEO2R分析
從NCBI基因表達綜合數據庫(https://www.ncbi.nlm.nih.gov/)下載miRNA表達芯片GSE19980。GSE19980 miRNA表達芯片數據集有12個樣本,6個細胞株HepG2(樣本編號:GSM499316~GSM499321)和6個表達HBV細胞株HepG2.2.15樣本(樣本編號:GSM499322~GSM499327)。采用GEO2R工具(https://www.ncbi.nlm.nih.gov/geo/geo2r/)對GSE19980數據進行分析,定義HepG2細胞為1組,HepG2.2.15細胞為2組,按P值從小到大排序,選取前250個具有顯著表達差異的miRNA執行分析。將分析結果置于Echart(http://www.ehbio.com/ImageGP/)中制作火山圖,篩選差異miRNA。
從NCBI下載GSE121248 mRNA表達芯片數據集,GSE121248數據集是以慢性乙型肝炎誘導的HCC及其鄰近正常組織中分離出的組織,并提取總RNA進行Affymetrix基因微陣列分析芯片,共有107個樣本,其中70個HCC樣本(樣本編號:GSM3428716~GSM3428785),37個鄰近正常組織樣本(樣本編號:GSM3428786~GSM3428822)。采用GEO2R對GSE121248數據進行分析,定義70個HCC樣本為1組,37個鄰近正常組織樣本為2組,按P值從小到大排序,選取P<0.001的差異表達基因進行后續分析。
1.2.2細胞培養
HepG2細胞和HepG2.2.15細胞采用含10%胎牛血清、1%青霉素鏈霉素溶液的高糖DMEM、37 ℃、5%二氧化碳培養箱中培養。
1.2.3miR-10b表達的熒光定量PCR檢測
利用TRIzol提取細胞總RNA,Nanodrop 2000測量總RNA濃度及純度。miRNA逆轉錄按All-in-OneTMmiRNA qRT-PCR Detection試劑盒說明書操作,合成的第1鏈cDNA稀釋10倍后于Bio-Rad CFX96實時熒光定量PCR儀進行擴增。PCR條件為95 ℃ 5 s、63 ℃ 30 s、72 ℃ 30 s進 行35個循環。以U6作為內參。hsa-miR-10b上游引物序列:5′-ACACTCCAGCTGGGTACCCTGTAGAACC-3′,內參U6上游引物序列:5′-CTCGCTTCGGCAGCACA-3′,qPCR檢測的通用下游引物為試劑盒中的Universal Adaptor PCR Primer。以2-ΔΔCt方法評估miR-10b的相對表達水平。
1.2.4miR-10b的靶基因預測
選擇TargetScan (http://www.targetscan.org/vert_72/)、miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/php/index.php)、miRDB (http://www.mirdb.org/)、DIANA TOOLS (http://diana.imis.athena-innovation.gr/DianaTools/index.php) 4種在線靶基因預測工具篩選出miR-10b的靶基因,使用在線工具Venny 2.1(https://bioinfogp.cnb.csic.es/tools/venny/index.html)繪制4個數據庫預測靶基因的韋恩圖,選取并集及交集靶基因用于后續分析。
1.2.5miR-10b靶基因的生物信息學分析
采用在線工具DAVID(https://david-d.ncifcrf.gov/)對miR-10b的交集靶基因進行基因本體(gene ontology,GO)分析,選擇物種為Homo sapiens,背景也選擇 Homo sapiens,包括生物過程、分子功能、細胞組分3種類型。以P<0.05為顯著性閾值,富集分數為-lg P,采用在線工具STRING (https://string-db.org/)對miR-10b的交集靶基因進行KEGG通路富集分析。
1.2.6miR-10b靶基因PPI網絡構建
將上述4個miRNA靶基因預測網站分析的miR-10b靶基因并集與GSE121248數據集GEO2R分析結果進行Venny分析以獲得共同基因,并將結果上傳在線工具STRING (https://string-db.org/),選擇物種為 human species,蛋白質相互作用評分大于0.4執行PPI 網絡分析,將PPI 網絡分析結果導入Cystoscape,選取節點度大于10的蛋白與miR-10b的靶基因4個數據庫預測交集結果進行Venny 2.1分析,獲得核心蛋白用于下一步分析。
1.3 統計學處理
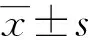
2 結 果
2.1 HBV感染上調miR-10b的表達
hsa-miR-10b在穩定表達HBV的HepG2.2.15細胞中表達明顯上調,差異有統計學意義(P<0.05),見圖1A。HepG2細胞miR-10b的相對表達水平[(0.980±0.079)]明顯低于穩定表達HBV的HepG2.2.15細胞[(2.682±0.937)],差異有統計學意義(P<0.05),見圖1B。
2.2 miR-10b基因位點與序列保守性
在miRBase數據庫中檢索人miR-10b的位置及其他物種miR-10b成熟序列,人miR-10b定位于2q31.1(chr2:176150303-176150412),其成熟序列為5′-UACCCUGUAGAACCGAAUUUGUG-3′。將10種其他物種miR-10b成熟序列與人類相比較發現,不同物種miR-10b具有高度保守性,見表1。提示 miR-10b具有潛在的重要生物學功能。
2.3 miR-10b的靶基因預測
使用Targetscan、miRTarBase、miRDB、DIANA TOOLS 4種在線工具預測miR-10b的靶基因,分別為340、325、352、464個,見圖2。用Venny2.1軟件繪制韋恩圖篩選獲得23個交集靶基因,見圖3。作為后續分析的基因數據集見表2。

A:差異表達miRNA火山圖;B:HepG2與HepG2.2.15細胞中miR-10b的表達水平比較;*:P<0.05,與HepG2.2.15細胞比較。

表1 不同物種的miR-10b的成熟序列
2.4 miR-10b靶基因數據集的GO及KEGG信號通路分析
miR-10b的23個靶基因參與了轉錄因子復合體、囊泡、核質部分等細胞組分;主要富集在轉錄調控、RNA代謝、基因表達等生物學過程,以及轉錄調控、轉錄因子活性和DNA結合等分子功能,見圖3。miR-10b的靶基因明顯富集在癌癥中蛋白多糖、癌癥中miRNA、晝夜節律及cAMP信號通路等相關通路中,差異均有統計學意義(P<0.05)。

表2 交集的靶基因數據集
2.5 miR-10b靶基因編碼蛋白的相互關系網絡
miR-10b靶基因與慢性乙型肝炎誘導的HCC及其鄰近的正常組織基因芯片GSE121248數據集GEO2R分析結果進行Venny分析獲得573個共同基因,并將此573個基因上傳在線工具STRING (https://string-db.org/),分析573個基因表達蛋白之間的相互作用,去掉無相互作用的蛋白質,獲得501個相互作用的蛋白,將STRING分析結果導入Cystoscape后執行分析,選取節點度大于10共獲93個蛋白基因,將此93個基因與miR-10b 4個網站預測23個交集的靶基因進行Venny 2.1分析獲得CREB1、GATA3、NCOA6、NCOR2、PIK3CA、MAPRE1、TIAM1 7個核心蛋白基因。

圖2 miR-10b的靶基因預測

A:細胞組分分析;B:生物學過程分析;C:分子功能分析;D:KEGG pathway分析。
3 討 論
盡管越來越多的研究表明,miR-10b在一些病毒性疾病的發病機制中具有關鍵作用,但其在HBV感染所致疾病中的作用仍鮮見文獻報道。本研究通過分析NCBI數據庫中的基因芯片數據發現,miR-10b在HBV感染肝細胞株HepG2.2.15中的表達明顯升高。熒光定量PCR檢測結果證實,HepG2.2.15細胞miR-10b的表達水平明顯高于HepG2細胞。表明HBV感染可上調細胞miR-10b的表達。
MiR-10b在生物學方面具有重要功能。比對人類、小家鼠、獼猴、黑猩猩和牛等不同物種miR-10b序列的結果顯示,其序列高度保守。實際上GO分析顯示,miR-10b的靶基因一是生物過程方面與轉錄調控、RNA代謝和基因表達等相關;二是分子功能方面與轉錄調控、轉錄因子活性和DNA結合等相關;三是細胞組分方面與轉錄因子復合體、囊泡和核質部分等相關。KEGG信號通路分析提示,miR-10b的靶基因明顯富集在癌癥中蛋白多糖、癌癥中miRNA、晝夜節律及cAMP信號通路等相關通路中。以GEO2R分析慢性乙型肝炎誘導的HCC及其鄰近正常組織基因芯片GSE121248數據集差異表達基因與miR-10b靶基因進行Venny 2.1分析,獲得了與HBV感染相關的miR-10b靶基因,進而STRING分析基因表達蛋白的相互作用確定了7個核心蛋白基因,分別為CREB1、NCOA6、NCOR2、PIK3CA、GATA3、MAPRE1和TIAM1。
CREB是正常細胞中許多重要信號傳導通路的最終調節因子,在小鼠胰島素分泌細胞INS-1中敲除CREB則上調caspase-3基因表達,促進細胞凋亡[10]。CREB可能通過誘導細胞保護因子——胰島素受體底物-2(insulin receptor substrate-2,IRS-2)和神經元PAS結構域蛋白4(neuronal PAS domain protein 4,NPAS4)表達促進β細胞增殖[11]及存活[12-13]。此外,CREB可增強核心囊泡跨膜蛋白IA-2基因的表達促進胰島素分泌[14]。NCOA6是一種轉錄共激活因子,NCOA6可調節胰腺β細胞中Nampt基因刺激胰島素分泌[15]。磷酸肌醇-3激酶(phosphoinositide 3-kinase,PI3K)/磷脂酰3,4,5三磷酸(phosphatidylinositol 3,4,5-trisphosphate,PIP3)是胰島素信號作用的重要信號途徑[16-19],介導了所有細胞對胰島素的反應。活化的PI3K亞基可解除糖原合成酶激酶3(glycogen synthase kinase 3,GSK-3)對糖原合成酶的抑制作用,促進肝糖原的合成,PI3K-C2γ敲除則導致實驗小鼠肝臟糖原積累嚴重降低[20]。PI3K可通過促進細胞內囊泡中的葡萄糖轉運蛋白4(glucose transporter 4,GLUT4)向細胞膜的轉運,從而促進肌肉及脂肪組織葡萄糖的吸收[21-24]。miR-10b對CREB1和NCOA6的靶向作用可能降低胰島β細胞的活性、下調胰島素基因的表達及抑制胰島素的分泌。miR-10b對PIK3CA的靶向作用可能會促進肝臟中糖原的分解,并阻止骨骼肌和脂肪組織中的葡萄糖攝取,從而導致機體糖穩態受損。這些研究結果顯示,miR-10b參與了調節胰島素分泌及葡萄糖攝取的生理過程。近年來,國內外研究表明,HBV感染可誘發2型糖尿病或增加2型糖尿病發病風險[25-26]。新近有研究發現,HBV感染與妊娠糖尿病相關,可增加妊娠糖尿病發病率[27-28]。慢性HBV感染合并非酒精性脂肪肝患者普遍具有胰島素抵抗[29]。這些研究結果與miR-10b靶基因的生物信息學分析相吻合。miR-10b的其他靶基因編碼蛋白如GATA3被報道在脂肪形成和炎癥中具有重要作用[30]。新近有研究指出,其可作為胰島素抵抗和2型糖尿病的潛在治療靶標[31]。而miR-10b靶基因Tiam1和Mapre1編碼蛋白TIAM1和MAPRE1分別被報道與腫瘤的侵襲性有關[32]和促進肝癌細胞的細胞周期進程[33]。
綜上所述,HBV感染可上調細胞miR-10b的表達,而異常表達的miR-10b可能通過靶向沉默CREB1、NCOA6、NCOR2、PIK3CA、GATA3、MAPRE1、TIAM1等蛋白基因損害機體糖穩態調節,產生組織炎癥及促進腫瘤發生、發展,從而在HBV感染的肝臟疾病中發揮著重要作用。因此,HBV感染對miR-10b表達的影響及其在肝臟疾病的作用值得深入研究。
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中國生殖健康(2019年2期)2019-08-23 08:12:10
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
