扁桃-冬小麥間作模式下不同樹形對冬小麥產量和葉片光合能力的影響
2021-11-04 09:04:58韓守安
新疆農業科學 2021年9期
謝 輝,韓守安,王 敏,張 雯
(新疆農業科學院園藝作物研究所,烏魯木齊 830091)
0 引 言
【研究意義】特色林果栽培是新疆南疆的主要種植模式[1]。新疆南疆果樹主要以間作的方式在耕地中種植,果麥間作面積達3.33×105hm2[2]。果糧間作已成為南疆主要農業生產模式之一[3]。扁桃(AmygdaluscommunisL.)是新疆特色果樹,具有重要的經濟價值[4],栽培面積已超過7.5×104hm2,主要以間作模式種植[5]。特別是隨著樹齡和樹冠體積的擴大,進入盛果期后果樹對下層農作物遮陰日益突出。提高光能利用率是小麥高產的關鍵。小麥植株積累量受光合面積、光合強度、光合時間和光合消耗共同影響[6]。研究果糧間作模式下扁桃遮陰程度對間作冬小麥籽粒產量和葉片光合能力的影響,對新疆南疆果糧間作模式的選擇和優化有重要意義。【前人研究進展】朱德群等[7]指出旗葉是小麥的主要功能葉,其合成光合產物對籽粒產量的貢獻率高到達43%,提高小麥旗葉的光合速率、延長其光合速率高值持續期對于小麥產量的形成具有重要意義。弱光導致單葉凈光合速率下降,Burkey和Wells[8]研究認為,弱光導致葉片凈光合速率降低主要與單位葉面積內葉綠體數目降低,葉綠體功能改變有關。Zhao DD[9]和Huang Q等[10]研究進一步證實弱光導致光系統Ⅱ(PSⅡ)的光化學效率提高、電子傳遞效率下降。光響應曲線模型是光合作用對環境因子響應模擬模型系統的核心組分,光響應曲線的確定,有助于了解光反應過程的效率[11]。Xu CL等[12](2016)通過對遮陰程度對冬小麥光合作用影響的研究結果表明,重度遮陰條件下植株光合能力顯著降低,而輕度遮陰條件下小麥凈光合速率升高。光響應曲線模型是光合作用對環境因子響應模擬模型系統的核心組分,光響應曲線的確定,有助于了解光反應過程的效率[11]。【本研究切入點】目前有關間作對作物光合能力的影響主要是通過遮陰模擬試驗進行的,較難準確反映出間作模式下樹冠不同生育時期遮陰差異及光陰交替的日變化動態對小麥光合能力的影響。以生產中廣泛應用且樹體結構存在較大差異的4個扁桃樹形及不同間作區域為處理,研究果樹樹冠截光程度對間作冬小麥葉片光合能力的影響。【擬解決的關鍵問題】以扁桃-冬小麥(新冬20號)間作模式為研究對象,設置主干分層形(DC)、開心形(OC)、高干形(HS)和小冠半圓形(SC)等樹體結構指標存在較大差異的4個樹形為處理,分析扁桃樹冠截光對間作冬小麥光合能力和產量的影響,為新疆南疆果糧間作模式的優化提供科學依據。
1 材料與方法
1.1 材 料
試驗地點位于新疆喀什地區,莎車縣阿熱勒鄉14村扁桃豐產示范園,屬暖溫帶大陸性干旱氣候,年平均氣溫11.4℃,年日照時數為2 965 h,年平均降水量56.6 mm,無霜期192 d;土壤類型為沙壤土,土壤有機質含量1.524 g/kg,全氮含量0.519 g/kg,全磷含量0.775 g/kg,全鉀含量19.584 g/kg,速效氮39.067 mg/kg,速效磷10.129 mg/kg,速效鉀110.333 mg/kg。
扁桃(AmygdaluscommunisL.)-冬小麥(TriticumaestivumL.)間作種植模式,扁桃品種為晩豐,2007年定植,株行距(6×7)m,南北行向;冬小麥品種為新冬20號,10月2日播種,采用寬窄行種植,窄行10 cm,寬行20 cm,播種密度525×104粒/hm2。間作小麥種植行寬度6 m,小麥沿果樹行向種植,邊行具樹干直距離50 cm。
1.2 方 法
1.2.1 試驗設計
設置主干分層形(DC)、開心形(OC)、高干形(HS)和小冠(SC)形4個樹形處理和大田100%光照對照(CK)處理。5株每小區(隨行向連續水平排列),3次重復。各樹形處理樹體結構指標。播種前,采用塑料布對果樹小麥根系交接區域進行隔離,分隔寬度80 cm,長度與調查區域長度相同,避免地下部根系水肥競爭影響小麥生長。表1
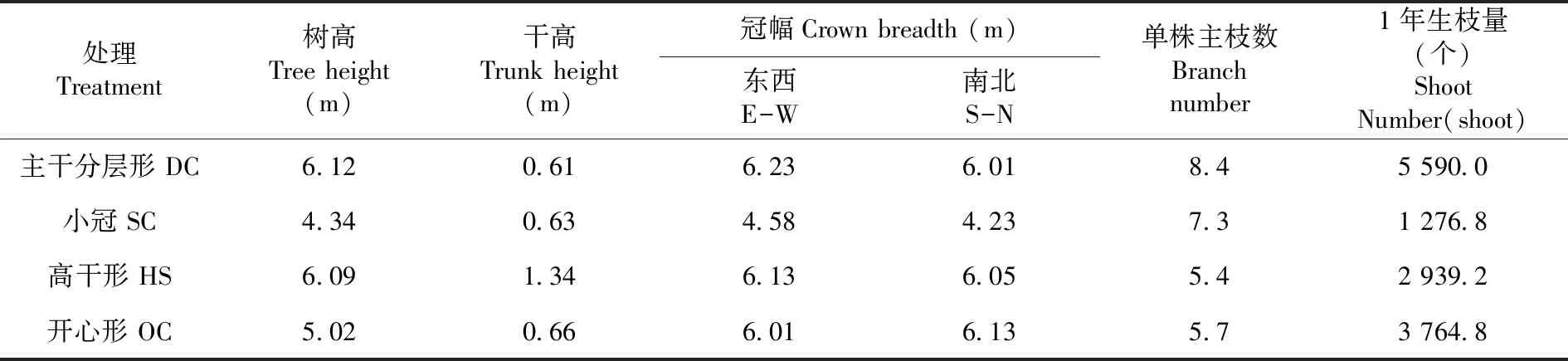
表1 盛果期扁桃不同樹形結構指標Table 1 The canopy structure index of different tree from at full bearing period
1.2.2 測定指標
分別于小麥抽穗期(4月21~23日)、揚花期(5月1~3日)、灌漿期(5月13~15日)和乳熟期(5月25~29日)4個關鍵生育時期隨機選擇Zone1(距樹干距離為1.5 m)、Zone2(距樹干距離為2.5 m)和Zone3(距樹干距離為3.5 m)3個間作區域內小麥旗葉對光響應曲線進行測定,采集旗葉樣品-80℃低溫保存,用于光合色素和可溶性蛋白含量的測定;小麥蠟熟期(6月10日),分區域進行測產和室內拷種。
1.2.2.1 旗葉光響應曲線
使用Li6400 便攜式光合測定系統(LICOR),于選取晴朗無云的晴天于太陽真時07:00~10:00測定,采用開放式氣路,測定旗葉的光響應曲線,選取各處理Zone3、Zone2和Zone1區生育進程一致的健康植株測定,每采用巡回測定,各測點重復3次。由高至低依次設置2 000、1 700、1 400、1 100、900、700、500、300、200、100、50、25和0 μmol/(m2·s)等13個光強梯度。
光響應曲線曲線的擬合:采用非直角雙曲線模型(non-Rectangular hyperbola equation)對光響應曲線進行擬合。
(1)
A為凈光合速率,Φ為表觀量子效率,Amax為最大凈光合速率,PPFD為光合有效輻射,k為光響應曲線曲角,Rday為暗呼吸速率。(具體方法參照:葉子飄,等2010)
1.2.2.2 光合色素和可溶性蛋白質含量
采用丙酮浸提法對旗葉光合色素含量進行測定,采用考馬斯亮藍比色法對旗葉可溶性蛋白含量進行測定。
1.2.2.3 產量及產量構成指標
各小區每個測點收獲0.5 m×0.5 m范圍內小麥全部植株進行測產,測定單位面積籽粒產量、單位面積有效穗數;同時從每小區各測點隨機選取20株進行室內拷種,分別測定穗粒數、千粒重。
1.3 數據處理
使用Excel2010、Origin2017和SAS數理統計軟件進行數據處理、統計分析和作圖。
2 結果與分析
2.1 果糧間作模式下樹體截光對間作冬小麥產量的影響
研究表明,籽粒產量、千粒重、穗粒數和單位面積有效穗數等指標,樹形、區域及二者的交互作用均達到極顯著水平(P<0.001)。與單作對照相比DC、SC、OC和HS 4個不同樹形處理對應間作區域間作冬小麥籽粒產量和產量構成指標均不同程度降低,其中籽粒產量各處理均顯著低于單作(P<0.05),降低比例依次為63.68%、18.24%、35.43%和33.00%;千粒重各處理均顯著低于單作(P<0.05),降低比例依次為15.80%、3.23%、9.28%和5.72%;穗粒數,除SC與單作間的差異未達到顯著水平外,其它3個處理均顯著低于單作(P<0.05),與單作相比4個樹形的降低比例依次為32.45%、2.13%、16.16%和3.25%;單位面積有效穗數各處理均顯著低于單作(P<0.05),降低比例依次為35.24%、12.02%、15.21%和32.48%。間作對千粒重的影響程度范圍為-22.94%~10.36%,變幅為33.30%;對單位面積有效穗數的影響范圍為-7.92%~-44.42%,變幅為36.5%;對穗粒數的影響范圍為0.37%~42.13%,變幅為41.76%。產量除SC處理Zone3區域與CK無顯著差異外,其它樹形和間作區域均顯著低于CK;穗粒數與CK間的差異較小,其中SC處理3個間作區域、HS處理Zone1、Zone3等2個區域與CK間的差異均未達到顯著水平;單位面積有效穗數除SC處理Zone2、Zone3區域與CK無顯著差異外,其它樹形和間作區域均顯著低于CK;千粒重,SC處理Zone3區域與CK間的差異未達到顯著性水平,其它處理和區域均顯著低于CK。表2
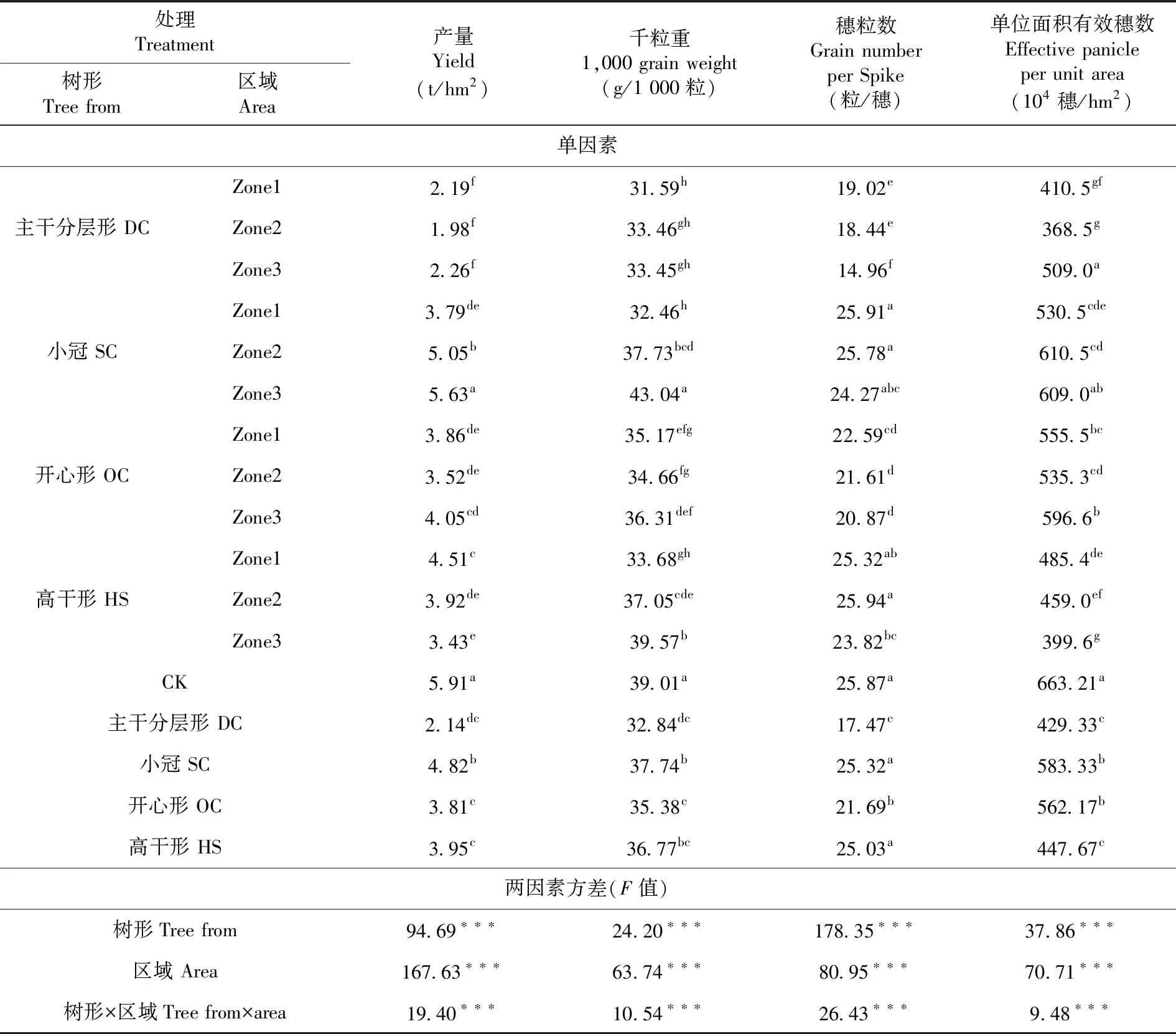
表2 不同樹形下間作冬小麥產量相關指標變化Table 2 Effect of tree shape on yield related indexes of intercropping winter wheat
2.2 果糧間作模式下樹體遮陰程度對間作冬小麥不同生育時期葉片質量的影響
研究表明,葉綠素a、b的含量和比值與葉片的光合能力具有密切的關系,不同樹形和區域間作冬小麥旗葉葉綠素a+b含量存在一定差異。抽穗期CK(單作)中葉綠素a+b含量為2.55 mg/g,與CK相比不同樹形處理表現出不同的變化趨勢,其中DC、SC較CK分別提高14.05%和6.46%,HS處理與CK的差異在1%以內,OC處理較CK降低3.25%;DC處理Zone3區域,SC處理3個區域和HS處理Zone1區域顯著高于CK,HS處理Zone2、Zone3區域和OC處理Zone3區域顯著低于CK,其它處理與區域與CK的差異均未達到顯著水平。CK為葉綠素a/b比值2.92,與CK相比不同樹形處理均不同程度降低,下降程度由高至低依次為DC、OC、HS和SC,降幅分別達到了14.57%、12.49%、10.95%和9.67%;除DC處理Zone1區域和OC處理Zone2區域與CK的差異未達到顯著水平外,其它處理和區域均顯著低于CK。
揚花期,CK旗葉葉綠素a+b含量為3.42 mg/g較抽穗期提高了34.12%,與CK相比不同樹形處理均不同程度降低,降幅由高至低依次為SC、DC、HS和OC,降幅分別為20.27%、12.09%、10.82%和6.63%;除DC處理Zone1區域和OC處理Zone3區域與CK無顯著差異外,其它處理和區域均顯著低于CK;CK旗葉葉綠素a/b為2.25,與CK相比不同樹形處理均不同程度提高,其中OC平均增幅最高達到處理32.44%;除DC處理Zone2、Zone3區域與CK不存在顯著差異外,其它處理和區域均顯著高于CK。
灌漿期,CK中葉綠素a+b含量為3.36 mg/g,不同處理表現出不同的變化趨勢,其中DC和HS處理與CK相比分別降低8.63%和18.16%,SC和OC處理分別較CK提高5.16%、2.08%和5.26%;DC和OC處理的Zone3區域、HS處理的Zone2和Zone3區域顯著低于CK,SC處理Zone1區域和OC處理Zone1、Zone2區域顯著高于CK,其它處理和區域與CK的差異均未達到顯著水平。CK旗葉葉綠素a/b比值為3.04,與CK相比不同樹形處理均不同程度降低,降幅由高至低依次為DC、SC、OC和HS,降幅依次達到16.67%、15.79%、15.02%和7.78%;各處理和區域均顯著低于CK。
乳熟期,CK旗葉葉綠素a+b含量為2.74 mg/g較灌漿期降低18.45%,不同樹形處理表現出不同的變化趨勢,其中DC處理較CK提高16.79%,SC、HS和OC處理分別較CK降低4.75%、5.96%,OC處理與CK基本一致;DC處理的Zone1和Zone2區域、OC和HS處理的Zone1區域顯著高于SC和OC處理的Zone3區域顯著低于CK,其它處理和區域與CK的差異均未達到顯著水平。CK旗葉葉綠素a/b比值為2.56,不同樹形處理表現出不同的變化趨勢,其中DC處理較CK降低7.81%,SC、和OC處理分別較CK提高8.33%和5.86%,HS處理與CK基本一致;DC處理的Zone3區域、SC處理的Zone2、Zone3區域和OC處理的Zone2區域顯著高于CK,HS和DC處理的Zone1區域顯著低于CK,其它處理和區域與CK的差異均未達到顯著水平。圖2,圖3
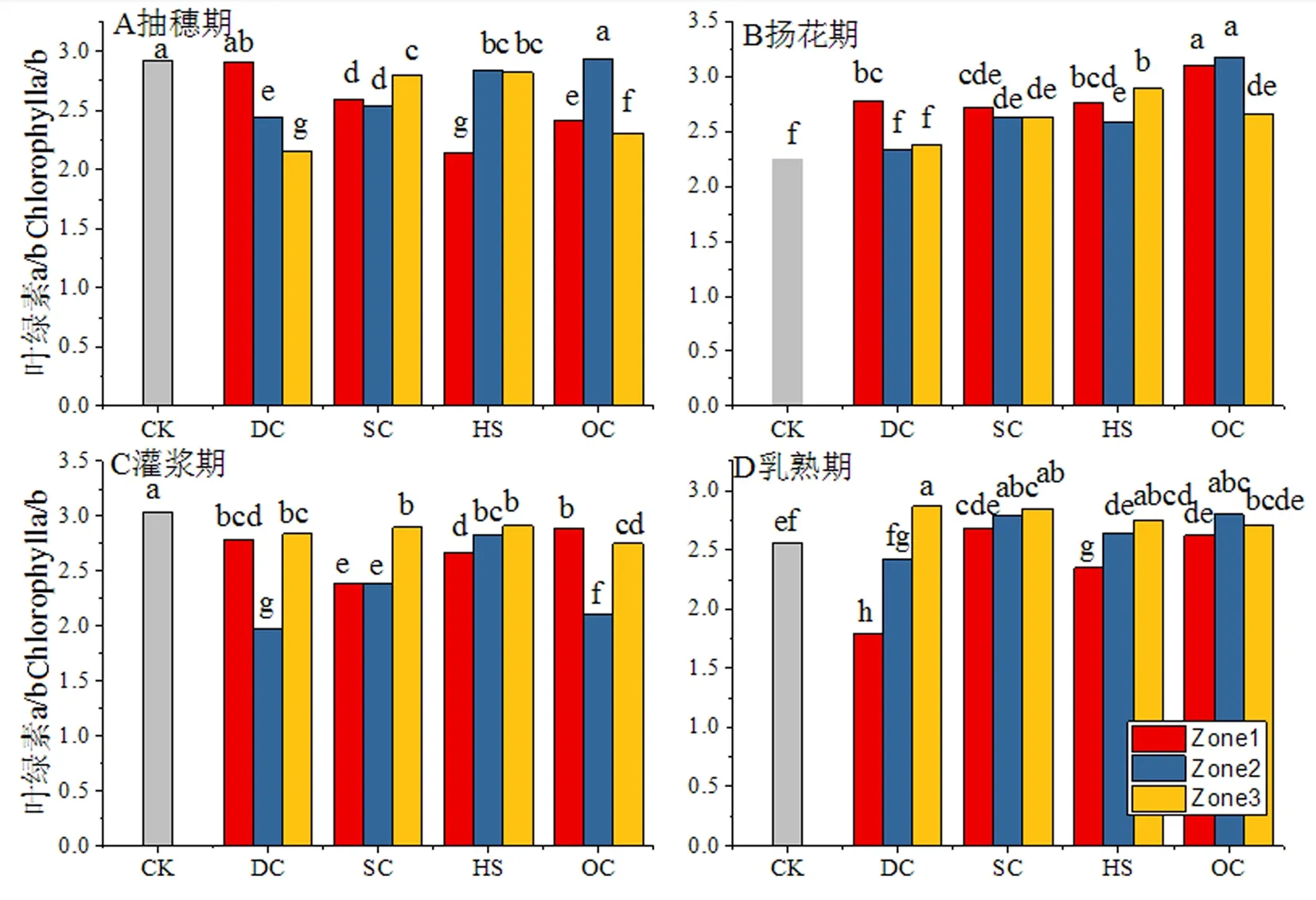
圖2 不同樹形下間作冬小麥不同生育時期旗葉葉綠素a/b變化Fig.2 Effect of tree shape on chlorophyll a/b ratio of flag leaf of intercropping winter wheat in different growth stages
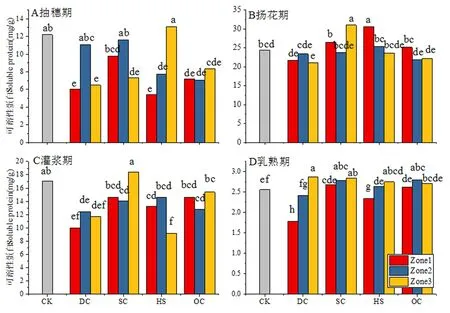
圖3 不同樹形下間作冬小麥不同生育時期旗葉可溶性蛋白含量變化Fig.3 Effect of tree shape on soluble protein content of flag leaf of intercropping winter wheat in different growth stages
研究表明,抽穗期,CK旗葉中可溶性蛋白含量12.19 mg/g,與CK相比不同樹形處理均不同程度降低,降幅由高至低依次為OC、DC、HS和SC,降幅分別達到38.20%、35.55%、28.22%和21.33%;除DC處理Zone2區域,SC處理Zone1、Zone2區域和與HS處理Zone3區域與CK間的差異未達到顯著水平外,其它處理和區域均顯著低于對照。
揚花期,CK旗葉可溶性蛋白含量為24.44 mg/g,較CK提高100.49%。與CK相比不同樹形處理表現出不同的變化趨勢,其中SC和HS處理較CK分別提高10.82%和8.52%,DC處理和OC處理較CK分別降低9.77%和5.70%;除SC處理Zone3和HS處理Zone1區域顯著低于CK外,其它處理和區域與CK間的差異均未達到顯著水平。
灌漿期,CK中可溶性蛋白含量為17.06 mg/g,較揚花期降低30.20%,與CK相比不同樹形處理均不同程度降低,降幅由高至低依次為DC處理33.18%、HS處理27.75%、OC處理16.39%和SC處理7.97%;除SC和OC處理的Zone1和Zone3區域及HS處理的Zone3區域與CK無顯著差異外,其它各處理和區域均顯著低于CK。
乳熟期,CK中可溶性蛋白含量降低至2.67 mg/g,與其它生育時期相比含量大幅降低,各樹形處理和區域均不同程度降低,但降低幅度存在較大差異,導致乳熟期冬小麥旗葉中可溶性蛋白含量存在較大差異。
不同處理和區域可溶性蛋白含量均高于CK,幅度由高至低依次為DC處理190.51%、HS處理189.23%、OC處理186.52%和SC處理162.92%;除SC和HS處理的Zone3區域與CK間的差異未達到顯著水平外,其它處理和區域均顯著高于CK。隨著生育進程的推進小麥旗葉葉片開始衰老,葉綠素含量降低,遮陰可以延緩葉片衰老,保持較高的葉綠素和可溶性蛋白含量。圖1~3
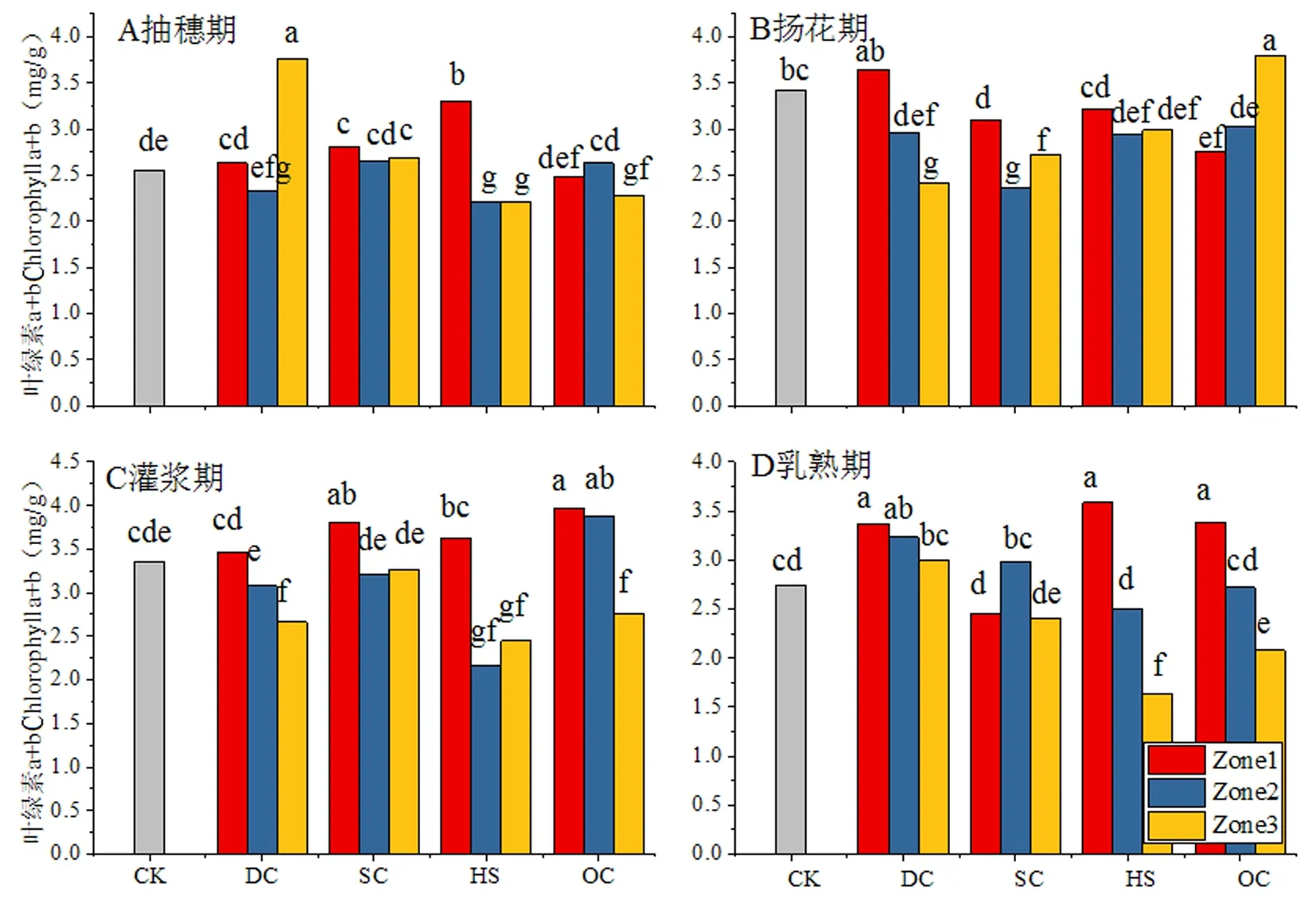
圖1 不同樹形下間作冬小麥不同生育時期旗葉葉綠素含量變化Fig.1 Effect of tree shape on chlorophyll content of flag leaf of intercropping winter wheat in different growth stages
2.3 果糧間作模式下樹體遮陰程度對間作冬小麥不同生育時期旗葉光合能力的影響
研究表明,R2均大于0.969 3,擬合效果較好。CK最大凈光合速率(Pmax)為19.68 μmol/m2s,光飽和點為(Lsp)為1 723.94 μmol/m2s,DC處理Zone1~Zone3區域與CK相比,Pmax分別降低48.88%、50.35%和30.28%,Lsp分別降低9.56%、22.86%和9.26%;SC處理Zone1~Zone3區域與CK相比,Pmax分別降低15.39%、5.89%和1.68%,Lsp分別降低6.58%、4.67%和6.68%;OC處理Zone1~Zone3區域與CK相比,Pmax分別降低35.41%、30.94%和11.59%,Lsp與CK差異較小均在2%以內;HS處理Zone1~Zone3區域與CK相比,Pmax分別降低11.13%、9.91%和3.35%,Lsp較CK分別提高5.83%、4.95%和14.64%。圖4
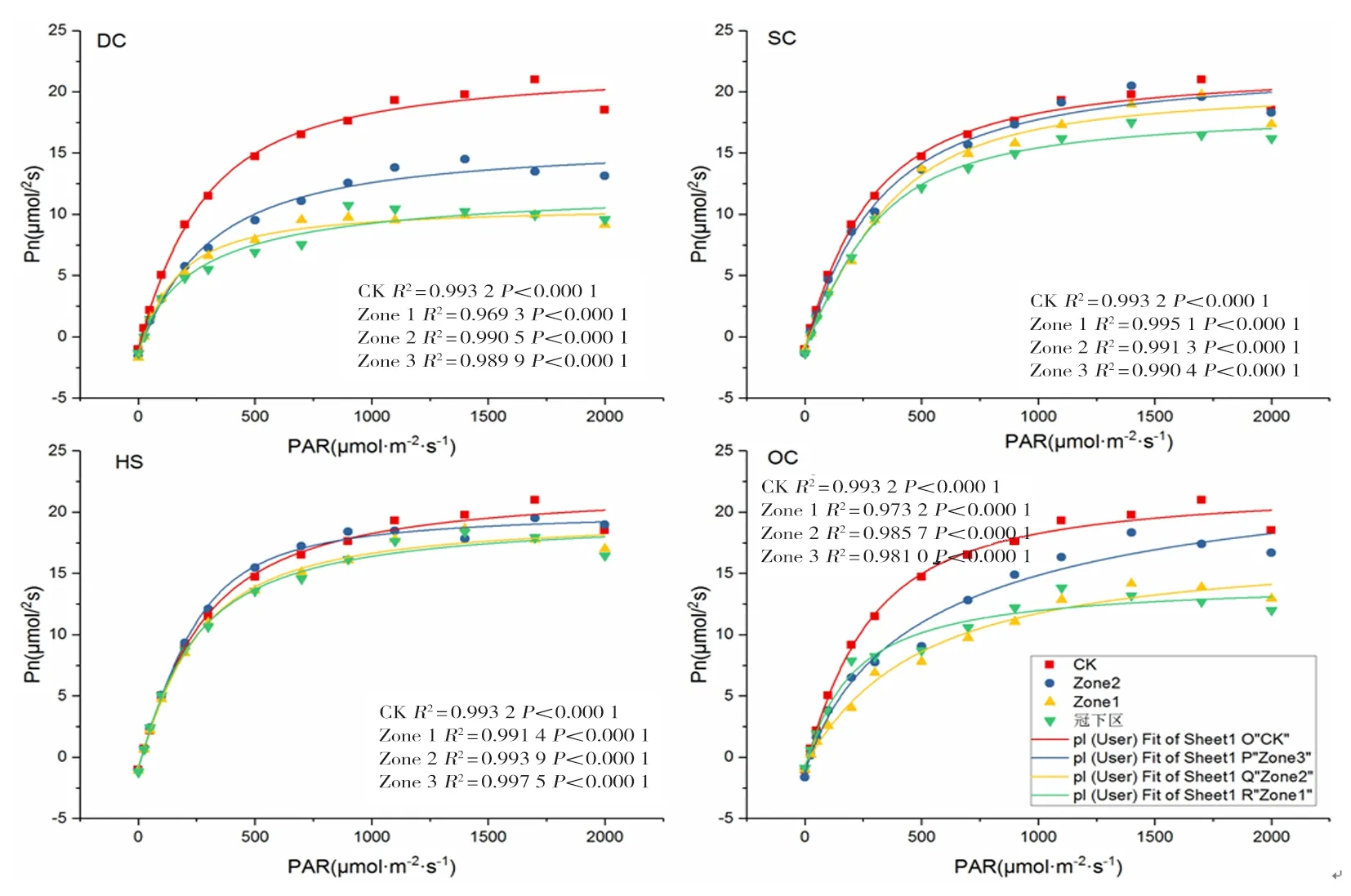
圖4 扁桃冬小麥間作體系中冬小麥抽穗期旗葉光響應曲線Fig.4 Light response curve of flag leaf at heading stage of winter wheat in almond-winter wheat intercropping system
R2均大于0.969 3,擬合效果較好。CK旗葉Pmax為24.18 μmol/m2s(較拔節期提高18.61%),Lsp為1 838.19 μmol/m2s,DC處理Zone1~Zone3區域與CK相比,Pmax分別降低48.88%、44.21%和33.87%,Lsp分別降低22.83%、23.77%和18.80%;SC處理Zone1~Zone3 3個間作區域Pmax分別較CK降低22.16%、18.24%和18.69%;Lsp,Zone1、Zone2等2個間作區域較CK降低8.60%和5.23%、Zone3區域較CK提高19.55%;OC處理Zone1~Zone3區域與CK相比,Pmax分別降低40.90%、40.07%和35.61%,Lsp分別降低15.77%、14.58%和14.64%;HS處理Zone1~Zone3區域與CK相比,Pmax分別降低42.02%、33.83%和27.21%,Lsp較CK分別降低15.77%、14.58%和14.56%。圖5
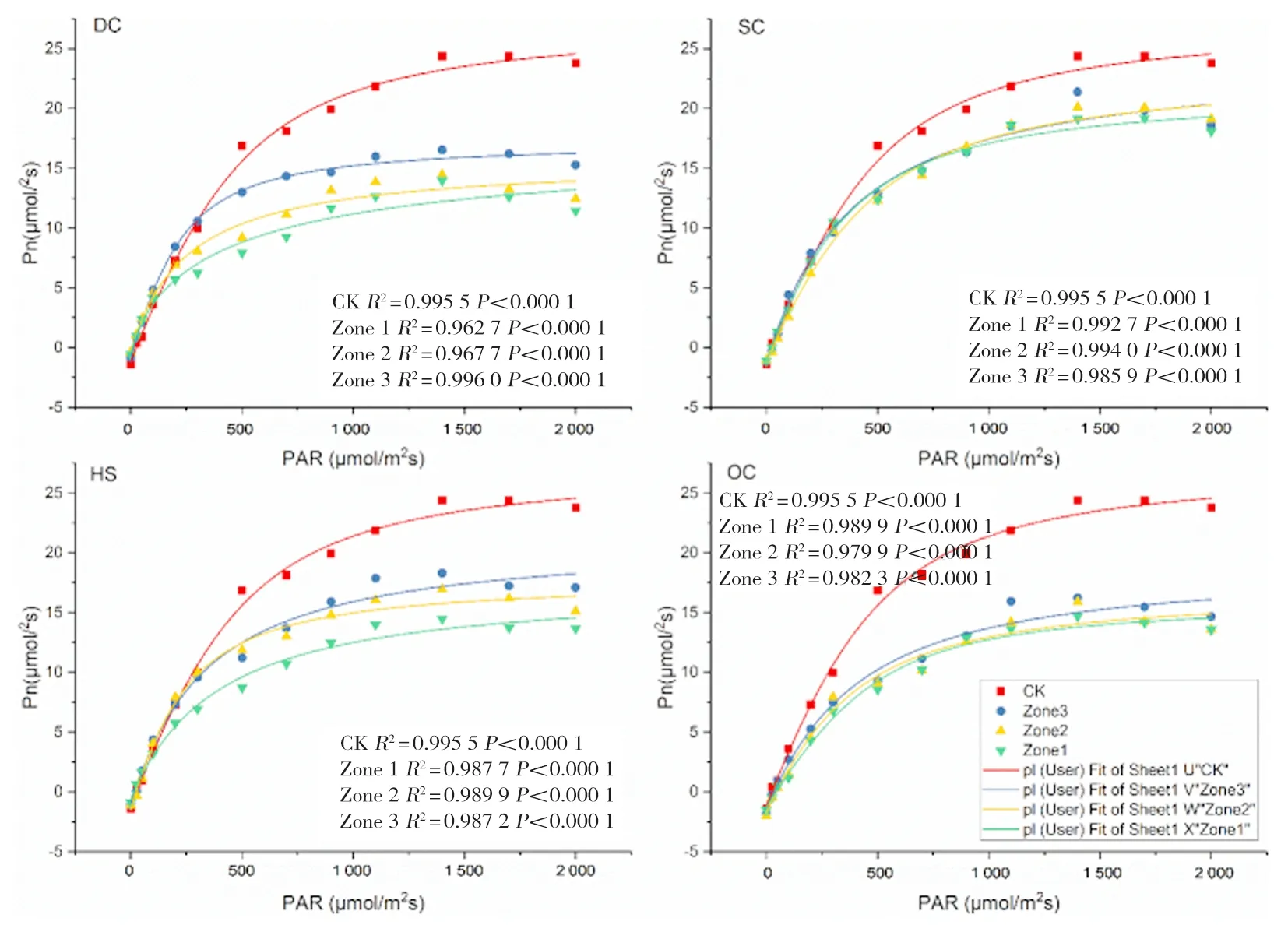
圖5 扁桃冬小麥間作體系中冬小麥揚花期旗葉光響應曲線Fig.5 Light response curve of flag leaf at flowering stage of winter wheat in wlmond-winter wheat intercropping system
研究表明,CK旗葉Pmax為23.98 μmol/m2s(較花期降低0.80%),Lsp為1 755.39 μmol/m2s,DC處理Zone1~Zone3區域與CK相比,Pmax分別降低54.30%、56.38%和52.29%,Lsp分別降低20.77%、22.17%和15.98%;SC處理Zone1~Zone3 3個間作區域Pmax分別較CK降低23.85%、23.85%和15.55%;Lsp,Zone1和Zone2區域較CK降低7.06%和1.89%、Zone3區域較CK提高2.23%;OC處理Zone1~Zone3區域與CK相比,Pmax分別降低41.45%、28.82%和25.15%,Lsp分別降低12.57%、0.57%和3.96%;HS處理Zone1~Zone3區域與CK相比,Pmax分別降低42.02%、33.83%和27.21%,Lsp較CK分別降低8.00%、7.01%和6.37%。圖6
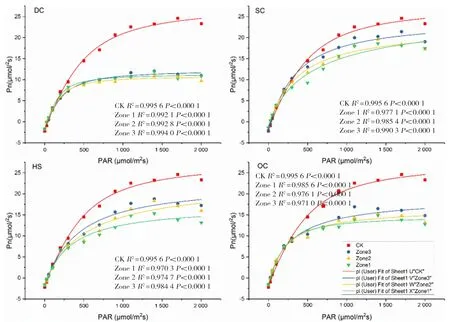
圖6 扁桃冬小麥間作體系中冬小麥灌漿期旗葉光響應曲線Fig.6 Light response curve of flag leaf at grain filling stage of Winter Wheat in Almond-winter wheat intercropping system
研究表明,CK旗葉Pmax為7.32 μmol/m2s(較花期降低69.48%),Lsp為1 369.98 μmol/m2s(較灌漿期降低21.97%),DC處理Zone1~Zone3區域與CK相比,Pmax分別提高26.91%、18.17%和18.72%,Lsp分別提高3.57%、5.97%和5.40%;SC處理Zone1區域較CK提高25.68%,Zone2和Zone3區域分別較CK降低4.37%和20.22%;Lsp提高2.44%、8.55%和8.08%;OC處理Zone1~Zone3區域與CK相比,Pmax分別降低Zone1、Zone1 2個區域分別較CK提高28.55%和9.97%,Zone3較CK降低2.59%;Lsp,Zone1~Zone3等3個間作區域較CK分別提高5.62%、5.50%和16.55%;HS處理Zone1~Zone3區域與CK相比分別降低13.66%、0.68%和4.51%;Lsp分別降低17.49%、21.78%和23.11%。圖7
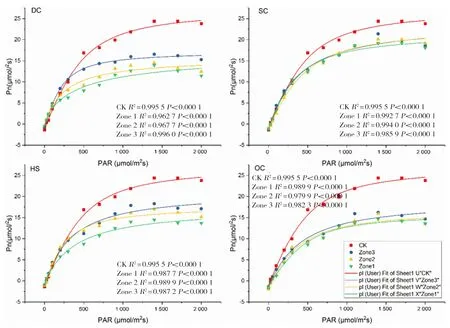
圖7 扁桃冬小麥間作體系中冬小麥乳熟期旗葉光響應曲線Fig.7 Light response curve of flag leaf at milk ripe stage stage of winter wheat in almond-winter wheat intercropping system
2.4 扁桃冬小麥間作模式下不同生育時期旗葉光合能力與產量指標的相關性
研究表明,小麥抽穗期至灌漿期小麥旗葉最大凈光合速率與產量、千粒重、單位面積有效穗數和穗粒數的相關性均呈顯著或極顯著正相關。抽穗期小麥旗葉最大凈光合速率與花期干物質積累量、抽穗期至灌漿期最大凈光合速率與成熟期干物質積累量均呈顯著正相關。其中產量、千粒重和穗粒數3個指標與灌漿期旗Pmax相關性最高,相關系數分別達到達到0.873、0.825和0.765;與抽穗期旗葉Pmax也有很高的相關性,相關系數分別達到0.829、0.818和0.730。葉花前干物質轉移量對子里貢獻率與花后干物質積累量對籽粒貢獻率均分別與灌漿期旗葉最大光合速率呈顯著負相關和正相關。旗葉光合能力和群體光合速率均與小麥產量密切相關,花后干物質積累量對籽粒貢獻率降低主要是由于灌漿期光合產物合成量下降引起的,間作花前干物質轉移量對籽粒貢獻率提高主要是由于花后光合產物積累量降低而導致的相對提高。表3
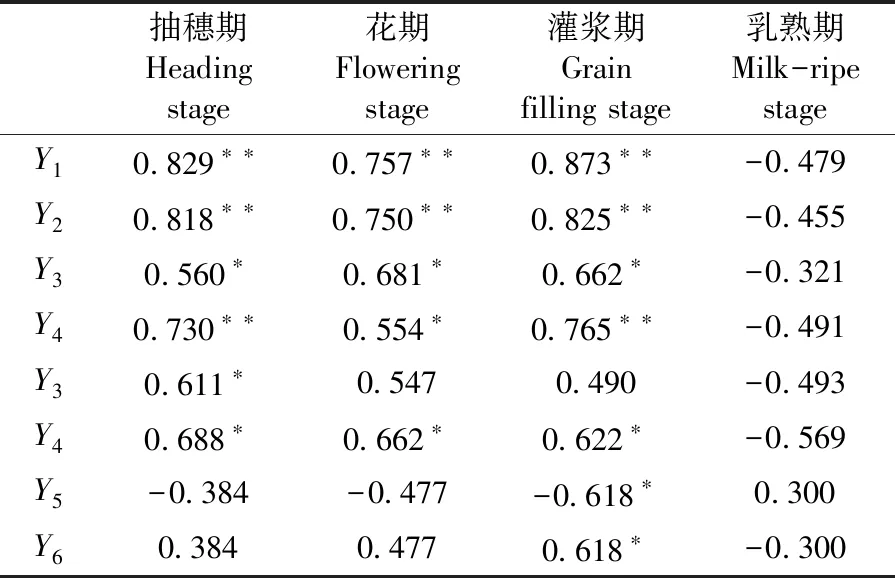
表3 扁桃-冬小麥間作不同生育時期旗葉最大凈光合速率與產量構成指標的相關性Table 3 Correlation between maximum net photosynthetic rate(Pmax)of flag leaves and yield component index of wheat at different stage in almond-winter wheat intercropping system
3 討 論
3.1 扁桃冬小麥間作體系中樹形對冬小麥旗葉質量的影響
葉綠素是光合作用的基礎,在光能的吸收、傳遞和轉換中起著重要作用[13],直接影響了小麥光合作用的進行。前人關于弱光對葉綠素組分影響的研究結果存在一定的差異,部分研究證實遮光導致小麥葉片中葉綠素含量增加,但葉綠素a/b比值降低,并指出這種轉變有利于葉片對光能的吸收[14,15]。李文陽等[16]認為,小麥花后弱光脅迫導致總葉綠素含量和葉綠素a/b比值降低。研究表明,扁桃冬小麥間作模式下,抽穗期和花期小麥旗葉葉綠素含量不同處理和區域間表現出不同的變化趨勢,至灌漿期間作模式下小麥旗葉葉綠素a/b值均顯著降低,但葉綠素含量不同處理間表現出不同的變化趨勢,分析認為可能與不同處理和區域間遮陰強度不同有關。
小麥旗葉可溶性蛋白80%以上為Rubisco酶[17],Rubisco是葉片光合作用的最關鍵酶,對于葉片光合具有重要意義。牟會榮等[18]關于拔節至成熟期遮光對小麥光合特性影響的研究結果表明,遮光導致小麥旗葉可溶性蛋白含量顯著降低了。有關遮陰對番茄光合能力的研究也得到了類似的結果。該文研究結果表明,間作導致小麥灌漿期葉片可溶性蛋白含量顯著降低,與前人研究結果一致。
3.2 扁桃冬小麥間作體系中樹形對冬小麥旗葉光合能力的影響
弱光導致了小麥光合色素含量和比例的變化,進而對葉片的光合能力造成影響[19],導致凈光合速率的降低和熒光動力學參數的變化。研究普遍認為弱光會導致小麥凈光合速率的下降,Xu C L等[12]通過對遮陰程度對冬小麥光合作用影響的研究結果表明,重度遮陰條件下植株光合能力顯著降低,而輕度遮陰條件下小麥凈光合速率升高。研究表明,扁桃小麥間作模式下,遮陰程度較輕的區域小麥旗葉光合速率與對照基本一致,部分光照強度下略高于CK,但遮陰較重的區域旗葉光合能力顯著降低,研究結果與Xu C L一致。但隨著生育進程的推進,花期和灌漿期各間作區域小麥旗葉光合能力均降低。郭峰等[15]研究表明,遮陰導致小麥旗葉凈光合速率、光飽和點出現不同程度的下降,該文研究結果表明,灌漿期各處理和區域小麥旗葉最大凈光合速率、光飽和點均降低。
牟會榮等[20]研究指出,長期遮陰能夠延長葉片光合速率高值持續期延長,延緩葉片衰老,但同時也指出由于長期遮光改變了光合系統活性的活性,盡管拔節至成熟期遮光條件下小麥旗葉的衰老有所延緩,但其光合速率(特別是后期)卻明顯低于對照。研究獲得了相似的結果,乳熟期間作小麥葉綠素含量、可溶性蛋白含量、Pmax的降幅均較小,但Pmax與CK并無較大差異,說明間作能夠延緩冬小麥旗葉衰老,但光合速率仍低于CK。
3.3 扁桃冬小麥間作體系中冬小麥產量與與葉片光合能力的相關性
單位面積有效穗數、穗粒數和粒重是構成小麥產量的重要指標,小麥籽粒產量的高低是由三者共同決定的,3個產量構成指標即相互聯系又相互制約。間作對小麥產量的影響受樹種、冠幅、間作區域均樹冠距離等多種因素共同影響。Xu Qiao等[21]通過對新疆南疆不同果糧間作模式的研究發現,與單作對照相比杏-麥、核桃-小麥間作模式下單位面積穗數、穗粒數、千粒重和單位面積產量均顯著降低,而棗麥間作模式下小麥產量與單作未達到顯著差異水平。小麥產量受桐麥間距的影響,小麥距樹行越近,產量越低[22]。研究表明,扁桃-冬小麥間作模式下樹形和區域對冬小麥產量及構成指標的影響均達到顯著水平,且二者存在明顯的交互作用。
小麥籽粒產量的形成與源光合產物供應能力密切相關,源光合產物的供應能力取決于小麥冠層光能截獲量[9,23]和光合系統的效率[24]。同時由于遮陰改變了光合產物在不同器官間的分配比例,進一步導致產量花前光合產物對籽粒貢獻率的提高[25]。研究表明,抽穗期、花期、灌漿期旗葉最大凈光合速率與小麥千粒重和產量均呈極顯著正相關關系,花前干物質積累量與抽穗期旗葉最大凈光合速率呈顯著正相關;花后干物質積累量、花前干物質轉移量對籽粒貢獻率及花后干物質積累量與灌漿期的旗葉光合能力均達到顯著水平。
4 結 論
扁桃-冬小麥間作系統中,小麥產量及構成指標受樹形和間作區域的共同影響,產量由高至低依次為小冠半圓形(SC)、高干形(HS)、開心形(OC)和主干分層形(DC),與單作相比降幅分別達到18.24%、33.00%、35.43%和63.68%;揚花期和灌漿期4個樹形對應間作區域小麥旗葉光合能力均顯著降低,與單作相比揚花期和灌漿期降幅分別在18.24%~48.88% 和15.55%~56.38%;抽穗期、揚花期、灌漿期小麥旗葉最大凈光合速率與小麥產量、千粒重、穗粒數和單位面積有效穗數的相關性均達到顯著水平。扁桃-冬小麥間作體系內,間作導致小麥揚花期和灌漿期旗葉光合能力的下降與籽粒產量降低密切相關。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年9期)2021-11-26 07:41:24
今日農業(2021年14期)2021-11-25 23:57:29
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
