酶解對猴頭菇多糖結構特征及免疫活性的影響
2021-11-02 11:18:08余蕊宏孫夢珂陳仕雄張浚文劉曉盼黃一帆福建農林大學動物科學學院蜂學學院福建省獸醫中藥與動物保健重點實驗室福建福州350002
中國獸醫學報 2021年9期
關鍵詞:質量
余蕊宏,孫夢珂,雷 博,陳仕雄,張浚文,劉曉盼,任 喆,黃一帆,秦 韜 (福建農林大學 動物科學學院(蜂學學院) 福建省獸醫中藥與動物保健重點實驗室,福建 福州 350002)
猴頭菇(Hericiumerinaceus)屬擔子菌綱、多孔菌目、猴頭屬,是一種傳統的食用菌和藥用菌,具有滋補身體、助消化等功效[1]。多糖(polysaccharide)作為猴頭菇中的主要活性成分之一,是一類廣泛存在于動植物中的天然高分子活性物質[2]。大量研究表明,猴頭菇多糖(Hericiumerinaceuspolysaccharide,HEP)具有增強免疫力[3]、抗氧化[4]、降血脂[5]等生理功能,具有較高的藥用價值和保健價值。
多糖的生物活性與其結構密切相關,包括相對分子質量、構象、糖鏈類型等[6]。相對分子質量是影響多糖物理特性和生理活性的主要因素[7],不同相對分子質量的多糖具有不同的結構特征及功能特性,但大相對分子質量的多糖由于結構復雜、水溶性差等缺點,極大地限制了多糖的開發和應用。研究發現,經過降解后的多糖,其相對分子質量和黏度均得到降低,往往表現出更強的生物活性[8]。酶解法是多糖降解的一種常見方法,主要通過改變分子的結構特征來提高其生物活性,具有專一性強、高效性、降解條件溫和等優點[9]。目前,國內外研究表明[10-11],酶解修飾不僅可以改變多糖的相對分子質量大小,還可以顯著增強其生物活性,但關于猴頭菇酶解多糖(enzymatic hydrolysis ofHericiumerinaceuspolysaccharide,EHEP)的生物活性尚不清楚。
巨噬細胞(macrophages)屬于免疫細胞,具有識別、吞噬及清除病原微生物和凋亡的機體細胞,是機體抵抗感染的第1道防線[12]。當病原微生物等外源物質侵入機體時,巨噬細胞會產生大量具有生物活性的分子,比如一氧化氮(NO)、活性氧和細胞免疫因子等[13],從而發揮免疫調節作用。鑒于巨噬細胞在機體免疫系統中的重要地位,本研究采用酶解法制備EHEP,以巨噬細胞RAW 264.7為靶細胞,深入研究EHEP對巨噬細胞的生物學功能及其在機體免疫調節中的作用,為酶解多糖的開發與應用奠定理論基礎。
1 材料與方法
1.1 試驗材料HEP由實驗室制備[14](糖含量為81.3%);內切-1,5-α-阿拉伯聚糖酶(10 U/mg)購自Megazyme公司;PBS緩沖液購自Solarbio公司;DMEM培養液和雙抗(青、鏈霉素)均購自Hyclone公司;胎牛血清(FBS)購自Gibco公司;NO檢測試劑盒和噻唑藍(MTT)均購自碧云天生物技術有限公司;二甲基亞砜(DMSO)、脂多糖(LPS)、FITC-dextran均購自Sigma公司;FITC-CD40和FITC-CD86抗體均購自eBioscience公司。
1.2 主要儀器ABT 320-4M電子天平購自科恩公司;真空冷凍干燥機購自CHRiST公司;傅里葉紅外光譜儀、Multiskan Mk3酶標儀購自Thermo公司;臺式高速離心機購自BECKMAN公司;NovoCyte流式細胞儀購自艾森生物有限公司;二氧化碳恒溫培養箱購自西德Haraneus公司。
1.3 EHEP的制備用pH 4.0的緩沖液將內切-1,5-α-阿拉伯聚糖酶配制成質量濃度為5 g/L的酶溶液,同法配制4 g/L的HEP溶液,再按酶溶液:HEP溶液體積比1∶10的比例混勻。靜置5 min后,40℃反應1 h,沸水浴10 min后靜置1 h,4 000 r/min離心10 min,收集上清液置于100~500 kDa透析袋中透析。24 h后,冷凍干燥,即得EHEP。
1.4 相對分子質量的測定將質量濃度為1 g/L的HEP與EHEP溶液,通過高效凝膠色譜法測定其相對分子質量。色譜條件為:色譜柱選用安捷倫KS-805、KS-804和KS-802三根串聯,流動相為雙蒸水,流速為1 mL/min,檢測器為安捷倫1260 Infinity液相色譜儀配示差折光檢測器,柱溫為70℃,檢測器溫度為50℃。
1.5 紅外光譜分析稱取1 mg的HEP和EHEP樣品,分別與100 mg KBr粉末混合,研磨均勻后壓片。將制好的壓片放入紅外光譜儀內進行掃描,掃描范圍為4 000~400 cm-1,并對所得的紅外吸收圖譜進行分析。
1.6 掃描電鏡分析取少量HEP和EHEP分別黏在帶有導電膠的處理片上,用洗耳球吹去浮樣,經噴金處理后,置于掃描電鏡的樣品室中掃描分析,觀察HEP與EHEP的表面形貌。
1.7 原子力電鏡分析將樣品HEP和EHEP分別配制成質量濃度為10 mg/L的溶液,滴在新剝離的云母片上,經氮氣干燥后被置于原子力顯微鏡上觀測。
1.8 HEP和EHEP對RAW 264.7細胞的免疫調節活性
1.8.1RAW 264.7細胞的培養與傳代 小鼠巨噬細胞系RAW 264.7采用DMEM完全培養液(含10% FBS、1%雙抗)于37℃、5% CO2恒溫培養箱中進行培養,待細胞密度達80%以上時,PBS洗滌2次,加入0.25%的胰蛋白酶作用30 s,最后用DMEM完全培養液終止消化,并接種于新的細胞培養瓶中進行傳代培養,用于后續試驗。
1.8.2MTT法檢測RAW 264.7細胞的增殖活性 取對數生長時期的RAW 264.7細胞接種于96孔板中,細胞密度為1×105/mL,于37℃、5% CO2培養箱中培養24 h后,每孔加入含有HEP和EHEP的培養液(終質量濃度為0.391,0.781,1.563,3.125,6.250,12.500,25.000 mg/L),另設空白組。孵育24 h 后,除去上清后,每孔加入10 μL MTT溶液(5 g/L),繼續孵育4 h后,棄上清,每孔加入150 μL DMSO,振蕩10 min使沉淀完全溶解,最后于酶聯免疫檢測儀測定490 nm處的D值,并計算細胞增殖率。
細胞增殖率=加藥組D值/空白組D值×100%。
1.8.3試驗分組 根據1.8.2中RAW 264.7細胞增殖的試驗結果,分為以下4組:Blank組、LPS組(1 mg/L)、HEP組(0.391~6.250 mg/L)、EHEP(0.391~6.250 mg/L)。
1.8.4RAW 264.7細胞表面分子CD40和CD86的檢測 取對數生長時期的RAW 264.7細胞接種于6孔板中,細胞密度為1×106/mL,于37℃、5% CO2培養箱中培養24 h后,加入含有HEP和EHEP的培養液(終質量濃度為0.391,0.781,1.563,3.125,6.250 mg/L)。孵育24 h后,收集上清液備用,用PBS清洗2次,每孔各加入1 μL FITC-CD40和FITC-CD86,4℃避光孵育30 min后,PBS清洗2次,棄上清,用500 μL PBS重懸后,上流式細胞儀檢測。
1.8.5RAW 264.7細胞吞噬功能的檢測 參照1.8.4方法接種細胞于6孔板中。孵育24 h后,收集上清液備用,并用PBS清洗2次,每孔加入400 μL FITC-dextran,37℃避光孵育30 min后,加入PBS清洗3次,棄上清,用500 μL PBS重懸后,上流式細胞儀檢測。
1.8.6NO的測定 收集1.8.4或1.8.5中各組的細胞上清液,每組取出50 μL上清液轉移至另一新的96孔板中,并用完全培養液稀釋NaNO2標準品質量濃度為0,10,20,40,60,80,100 mg/L。在各孔中依次加入Griess Reagent Ⅰ、Griess Reagent Ⅱ試劑后,用酶標儀檢測540 nm波長處的D值。最后,以NaNO2標準品的濃度為橫坐標,D值為縱坐標繪制標準曲線,計算出各組上清液中NO的含量。
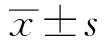
2 結果
2.1 HEP和EHEP的相對分子質量分析多糖相對分子質量的大小是影響其生物活性的主要因素之一,不同相對分子質量的多糖具有不同的生物活性,因此采用高效凝膠色譜法測定HEP和EHEP的相對分子質量。葡聚糖標準曲線方程為y=53.9-8.3x+0.53x2-0.015x3+0.000 17x4,將HEP和EHEP的洗脫體積代入到方程中,得到HEP的相對分子質量為4.3×104kDa,EHEP的相對分子質量為1.25×104kDa。
2.2 HEP和EHEP的紅外光譜分析紅外光譜可用于來鑒定化合物和官能團結構。如圖1所示,HEP與EHEP都包含3 600~3 200 cm-1的O-H吸收峰以及3 000~2 800 cm-1和1 410 cm-1的C-H伸縮振動和變角振動特征吸收峰[4],這3種峰均屬于多糖的特征峰,表明HEP與EHEP均為多糖類物質。HEP和EHEP分別在1 573.55,1 572.99 cm-1處有吸收峰,可能是有結晶水的存在;在575.85,620.39 cm-1的吸收峰表明HEP和EHEP均具有α-糖苷鍵。1 200~1 000 cm-1范圍出現的吸收峰是由于C-OH的變角振動引起的。與HEP紅外圖譜相比[4],EHEP在928.03 cm-1處有吸收峰,此峰屬于β型吡喃糖伸縮振動峰。

A.HEP紅外譜圖;B.EHEP紅外譜圖圖1 HEP和EHEP的紅外譜圖
2.3 HEP和EHEP的掃描電鏡和原子力電鏡分析掃描電鏡和原子力電鏡能觀察到多糖的微觀形貌,進一步分析多糖的結構特征。HEP和EHEP的掃描電鏡圖如圖2A所示,HEP的形態呈塊狀,菱角分明且表面光滑;EHEP呈不規則的形態,凹凸不平且表面粗糙,同時還出現帶裂痕的褶皺形貌。HEP和EHEP的原子力電鏡觀察結果如圖2B所示,兩者的空間結構均呈近似紡錘體形狀[4],且緊密排列在一起。HEP的高度范圍為-10.1~9.5 nm,EHEP的高度范圍為-10.3~10.7 nm。

A.掃描電鏡圖;B.原子力電鏡圖圖2 HEP和EHEP的掃描電鏡圖和原子力電鏡圖
2.4 HEP和EHEP對RAW 264.7細胞的免疫調節活性
2.4.1細胞增殖活性 MTT法是一種快速簡單檢測細胞增殖率的方法,可根據D值來判斷細胞的活性,從而間接反映細胞的增殖能力。由圖3可知,HEP和EHEP分別在0.391~3.125,0.391~25.000 mg/L質量濃度范圍的內細胞增殖率均顯著高于空白組(P<0.05),且呈現劑量依賴性,表明0.391~3.125 mg/L HEP和0.391~25.000 mg/L EHEP均可以顯著促進RAW 264.7細胞的增殖(P<0.05)。而當HEP的質量濃度增加至12.500,25.000 mg/L時,細胞的增殖率均顯著低于空白組(P<0.05)。

小寫字母相同的表示差異不顯著(P>0.05),不同的表示差異顯著(P<0.05)。下同圖3 HEP和EHEP對RAW 264.7細胞增殖的影響
2.4.2細胞表面分子CD40和CD86的表達 CD40和CD86屬于免疫球蛋白超家族成員,在刺激T細胞活化的過程中發揮重要作用,其表達量的多少與巨噬細胞的抗原提呈能力有著密切聯系。如圖4,5所示,HEP和EHEP均可以顯著促進RAW 264.7細胞表面分子CD40和CD86的表達(P<0.05),且呈現明顯的劑量效應。從圖4A,B中可以看出,在0.391~6.250 mg/L質量濃度范圍,HEP與EHEP的CD40表達率均顯著高于空白組(P<0.05),且EHEP組的CD40表達率顯著高于同等劑量下HEP組的表達率(P<0.05),以6.250 mg/L EHEP的CD40的表達率最高,為14.8%。從圖5A,B中可以看出,在0.391~6.250 mg/L質量濃度范圍,HEP與EHEP的CD86表達率均顯著高于空白組(P<0.05),以6.250 mg/L EHEP的CD86的表達率最高,為19.8%。結果表明,與HEP相比,EHEP更能增強RAW 264.7細胞表面分子CD40和CD86表達的能力。

圖4 HEP和EHEP對RAW 264.7細胞表面分子CD40表達的影響

圖5 HEP和EHEP對RAW 264.7細胞表面分子CD86表達的影響
2.4.3細胞吞噬活性 吞噬能力是衡量巨噬細胞活性的一個指標,活化后的巨噬細胞具有較強的吞噬能力,可通過吞噬侵入的病原體來提高機體的免疫調節作用。由圖6可知,HEP與EHEP在0.391~6.250 mg/L 的劑量范圍內均能不同程度增強RAW 264.7細胞的吞噬能力。與空白組相比,0.391,0.781,1.563,6.250 mg/L HEP的吞噬率均顯著升高(P<0.05);0.391~6.250 mg/L EHEP的吞噬率均顯著升高(P<0.05)。與HEP組相比,0.391 mg/L EHEP的吞噬率顯著高于HEP組中所有劑量下的吞噬率(P<0.05)。結果表明,EHEP在促進RAW 264.7細胞吞噬活性方面優于HEP。

圖6 HEP和EHEP對RAW 264.7細胞吞噬功能的影響
2.4.4NO含量 NO即是一種非特異性免疫的免疫分子,也是調節獲得性免疫的信使分子,在特異性免疫中發揮重要作用。NO是巨噬細胞發揮吞噬作用的基本條件,其分泌量的多少與巨噬細胞的吞噬能力呈正比。由圖7可知,HEP及EHEP在一定質量濃度范圍內均可促使RAW 264.7細胞釋放NO的量增加,且呈現明顯的劑量依賴性。隨著質量濃度的增加,HEP對細胞釋放NO的量逐漸減少,當HEP的質量濃度增加至3.125,6.250 mg/L時,NO的含量與空白組相比顯著降低(P<0.05),而EHEP隨著質量濃度的增加對細胞釋放NO的量逐漸增多,當EHEP質量濃度增加至6.250 mg/L時,NO的釋放量最高,為14.63 mg/L,且顯著高于HEP組中所有劑量下的NO含量(P<0.05)。

圖7 HEP和EHEP對RAW 264.7細胞生成NO含量的影響
3 討論
HEP是猴頭菇中分離出的一種具有天然活性的物質[15],具有抗氧化、免疫調節、抗腫瘤等[16-18]作用,但其相對分子質量較高,不利于細胞等生物體的吸收[19]。有研究表明[10-11],酶解法降解后的多糖具有較強的生物活性。在本研究中,我們發現EHEP能促進RAW 264.7巨噬細胞分泌NO、CD40和CD86,同時還能增強細胞的吞噬功能,因此可將EHEP作為潛在的免疫增強劑。
眾所周知,多糖的結構特征與其生物活性具有密切關系,不同種類的多糖具有不同的結構,因此活性也存在較大的差異。相對分子質量是其具備生物活性的必要條件,多糖具有一定的相對分子質量范圍,一般認為相對分子質量在10~500 kDa之間能保持最大活性[20]。若相對分子質量太大,則不利于其跨越細胞膜進入生物體內發揮生物活性作用;若相對分子質量過低,則無法形成產生活性的高級聚合結構[19]。本研究結果顯示,采用內切-1,5-α-阿拉伯聚糖酶對HEP進行酶解修飾后,EHEP的相對分子質量為1.25×104kDa。在前期的研究中[4],HEP的相對分子質量為4.3×104kDa,由此可以得出酶解修飾降低了HEP的相對分子質量,這與REN等[11]的研究結果一致。我們推測可能是酶解斷裂了HEP的糖鏈,進一步改變了其空間結構,從而降低HEP的相對分子質量。與HEP的紅外光譜相比[4],EHEP在HEP的峰值處均有吸收峰,但在575.85 cm-1處不具有吸收峰,而在928.03 cm-1處具有吡喃環的特征吸收峰,表明酶解對HEP的官能團結構產生了影響。HEP的掃描電鏡圖顯示[4],其形貌棱角分明且表面光滑。在本試驗中,EHEP呈現出凹凸不平的褶皺形貌,且表面變得粗糙。HEP屬于天然高分子多糖,糖鏈間通過氫鍵交聯,經過酶解后,導致糖鏈間氫鍵交聯密度變小,疇結構變大,從而導致表面粗糙度增加,這與報道結果一致[21]。HEP的原子力顯微鏡圖顯示[4],HEP的分子鏈高度為-10.3~10.7 nm,而本研究中EHEP的高度低于HEP,為-10.1~9.5 nm,與秦利鴻等[22]研究結果一致。多糖的生物活性是各級結構之間以及各種理化性質之間相互制約而產生的綜合效應[23],AIPIRE等[24]研究表明,烏拉爾甘草多糖是含有α、β糖苷鍵的多糖,其相對分子質量為1.49×104Da,具有較強的免疫調節作用,這與EHEP的結構特征類似,表明EHEP活性的增強與其各級結構的改變以及相互交聯有關。
MTT法可用于檢測細胞活性的強弱,細胞的增殖活性與吸光度成正比[25]。因此,確定細胞的安全質量濃度對于后續的研究具有重要作用。在本試驗中,0.391~3.125 mg/L HEP和0.391~25.000 mg/L EHEP均可以顯著促進RAW 264.7細胞的增殖(P<0.05),為保證試驗在同一水平上比較,取0.391,0.781,1.563,3.125,6.250 mg/L 5個劑量考察HEP和EHEP對RAW 264.7細胞的免疫活性作用。
巨噬細胞是機體重要的免疫細胞,主要通過吞噬作用清除侵入體內的病原微生物,在防御感染、自身穩定和免疫監視中都起著重要的作用[26]。研究發現[27],多糖具有增強免疫如活化巨噬細胞的功能,活化的巨噬細胞可分泌NO、CD40和CD86等從而調節免疫作用。NO即是免疫分子又是信使分子,在非特異性免疫和特異性免疫應答中發揮著重要作用。本試驗研究結果顯示,HEP和EHEP均能促進RAW 264.7細胞NO的釋放。與HEP相比,0.391~6.250 mg/L EHEP的NO釋放量均顯著高于同等劑量下HEP的NO釋放量(P<0.05),表明EHEP比HEP更能促進RAW 264.7細胞的活化,這與REN等[28]的研究結果一致。細胞表面共刺激因子在細胞免疫應答中起著重要的作用,而CD分子的活性可以反映出免疫細胞的活化程度,如CD40和CD86。CD40是細胞表面的糖蛋白,主要參與B細胞的生長及分化[29],CD86是CD80的配體,能夠和CD80一起調節T細胞的活化,在免疫反應的誘導和調節中起著重要作用[30]。在本試驗中,與空白組相比,HEP和EHEP均能促進RAW 264.7細胞表面因子CD40和CD86的表達。6.250 mg/L EHEP的CD40、CD86表達率最高,且顯著高于除LPS之外的其他各組(P<0.05)。由此可以得出,EHEP對巨噬細胞的免疫調節作用更強。
吞噬作用是巨噬細胞最重要的功能,可通過吞噬作用將病原微生物清除并維持機體內環境的穩態,其吞噬作用的強弱反映了免疫活性的高低、也可以反映其活化的程度[31]。多糖通過活化巨噬細胞,使其吞噬功能增強,從而調節免疫反應。本研究結果顯示,HEP和EHEP均能增強巨噬細胞的吞噬功能,但EHEP表現出更強的吞噬作用,這與CHENG等[32]的研究報道一致。
綜上所述,本研究采用內切-1,5-α-阿拉伯聚糖酶對HEP進行酶解修飾,HEP與EHEP作用于巨噬細胞后,EHEP表現出更強的免疫活性,可顯著增強巨噬細胞的增殖率,提高細胞表面分子CD40、CD86的表達及吞噬作用,由此可以得出酶解修飾能夠顯著提高HEP的體外免疫活性,這可能與相對分子質量的降低以及空間結構的變化存在一定的關系[33-34],但其作用機制還有待于深入研究。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
