不同發育期雞呼吸道相關淋巴組織中IL-2和IFN-γ mRNA表達水平的變化規律
2021-11-02 11:26:54楊樹寶張桂山張英楠吉林醫藥學院吉林吉林市3203吉林農業大學動物科技學院吉林長春308長春科技學院生命科學院吉林長春30600
中國獸醫學報 2021年9期
關鍵詞:水平
楊樹寶,張桂山,徐 晶,張英楠,3* (.吉林醫藥學院,吉林 吉林市 3203;2.吉林農業大學 動物科技學院,吉林 長春 308;3.長春科技學院 生命科學院,吉林 長春 30600)
呼吸道相關性淋巴組織(RALT)是黏膜相關性淋巴組織(MALT)的重要組成部分,MALT同時也包括腸道相關淋巴組織以及泌尿生殖道相關淋巴組織[1]。由于雞沒有外周淋巴結,所以MALT成為主要的次級免疫器官,對禽類的免疫保護起重要作用[2]。伴隨著呼吸道結構和功能的一系列轉變,RALT也發生著相應的變化。由于雞出殼后需要立即呼吸,從而呼吸道會接觸到與成雞接觸類型一致的混于空氣中的微生物群,且極有可能接觸到病原菌,而此時的雛雞呼吸道并未建立獲得性免疫的保護,雞在出殼后,母源抗體非常有限,因此,雞如何在出殼初期保護自身不受病原體侵擾,RALT發育及功能是否成熟,能否給呼吸道及機體提供免疫保護等問題值得研究。
IL-2是由激活的Th1細胞分泌產生的一類重要細胞因子之一,能抑制Th2型細胞發育,選擇性增強Th1型細胞分化增殖,亦能誘導T、B細胞增殖分化,促進自然殺傷細胞產生細胞因子,具有抗病毒、抗腫瘤的作用[3]。IFN-γ是由活化的T淋巴細胞和NK細胞等分泌的具有抗病毒、抗腫瘤和免疫調節等生物活性的重要細胞因子,能直接促進T、B淋巴細胞的分化,增強MHC-Ⅰ類和MHC-Ⅱ類分子的表達,是重要的免疫調節分子,常被用作感染機體的細胞介導免疫的指示器[4]。由于Th1細胞主要介導細胞免疫反應,其中起主要作用的是細胞因子IL-2和IFN-γ,因此本試驗利用實時熒光定量PCR方法檢測不同時期RALT中IL-2和IFN-γ mRNA的表達水平,在一定程度上反映不同時期雞RALT的細胞免疫功能變化情況,從而為研究雞出殼初期的免疫機制及雞呼吸道疾病(尤其是禽流感)的防治以及研制有效的鼻腔免疫疫苗提供理論依據。
1 材料與方法
1.1 實驗動物200枚海蘭褐蛋雞SPF種蛋,購自吉林卓越生物科技有限公司。
1.2 主要試劑及儀器RNAisoTMPlus、PrimeScript RT-PCR Kit、pMD?18-T 載體和SYBR?PrimeScript RT-PCR KitⅡ均購自大連寶生物;AxyPrepTM Plasmid Miniprep Kit 50-prep購自美國AXYGEN BIOSCIENCES;E.N.Z.A?Gel Extraction Kit 購自Omega公司。
依愛EIF/C.DME3456孵化機/出雛機(中國),NanoDrop 超微量高精度紫外/可見光光度計,Eppendorf 冷凍離心機(德國Eppendorf公司),Eppendorf AG,22331Hamburg梯度PCR儀,ABI Applied Biosystems StepOne實時熒光定量PCR儀(美國ABI公司)。
1.3 引物設計與合成根據GenBank中登錄的chIL-2、chIFN-γ和chβ-actin的序列,在保守區域分別設計1對特異性引物(表1)。引物由生工生物工程(上海)股份有限公司合成。

表1 IL-2、IFN-γ和β-actin的特異性引物
1.4 試驗設計利用孵化器對種蛋進行孵化,孵化溫度:37.5~38.2℃,濕度:60%~70%,每2 h自動翻蛋1次,出殼后,雛雞進行常規飼養。分別在15,18,20胚齡及1,4,7,14,21,35,56,90日齡,每個時期分別取5只雞胚或雛雞的氣管、喉和肺臟等組織,液氮速凍后-80℃保存備用。
1.5 總RNA提取與反轉錄按照RNAisoTMPlus (D9108A)說明書提取雞呼吸道各段組織的總RNA。參照反轉錄試劑盒(DRR037A)說明書進行RNA反轉錄,合成cDNA。
1.6 陽性標準品的制備以35日齡雛雞的肺臟cDNA為模板,對IL-2、IFN-γ和β-actin基因進行普通PCR擴增,目的基因經1%凝膠電泳后按照E.N.Z.A?Gel Extraction Kit說明書進行回收純化,然后將回收純化后的IL-2、IFN-γ和β-actin DNA與pMD 18-T載體進行連接、轉化至宿主感受態菌DH5α,將IL-2 、IFN-γ和β-actin陽性克隆進行序列鑒定,測序結果與在GenBank中登錄的雞IL-2 mRNA (AF000631),IFN-γ mRNA (X99774)及β-actin mRNA (X00182)的保守序列進行比對,運用DNAStar進行同源性分析。用核酸蛋白分光光度計測定IL-2、IFN-γ和β-actin DNA片段的陽性質粒D值,按照以下公式計算質粒拷貝數:
ds DNA拷貝數=
每一種陽性質粒分別進行10倍滴度稀釋,每個稀釋度設3個平行。
1.7 標準曲線的繪制采用SYBR GreenⅠ熒光染料法,以IL-2、IFN-γ和β-actin陽性標準品進行10倍梯度稀釋(1×10-1,1×10-2,1×10-3,1×10-4,1×10-5)濃度為模板,進行熒光定量PCR反應。不斷對模板濃度、引物濃度和退火溫度等進行優化,以出現最小Ct值和最高熒光強度為標準,最后計算Ct值,取5個點繪制3個基因的標準曲線。
1.8 樣品中目的基因的實時熒光定量檢測將方法1.5中得到的不同日齡呼吸道各段組織的cDNA進行熒光定量PCR,以檢測IL-2和IFN-γ mRNA的含量。各基因的反應體系均為20 μL,反應條件:95℃ 10 s;95℃ 5 s,60℃ 30 s,35個循環。
1.9 結果分析以20胚齡雞胚呼吸道RALT中目的基因mRNA相對表達水平為1,根據各時期IFN-γ和IL-2基因mRNA相對表達水平與20胚齡目的基因mRNA 相對表達水平的差異倍數進行統計分析,從而闡明不同發育階段雞RALT中IFN-γ和IL-2基因的動態表達情況。
2 結果
2.1 IL-2基因mRNA表達水平的動態變化由圖1A可見,胚胎時期肺臟中IL-2基因mRNA表達水平較低,且無明顯變化,出殼后1日齡時其表達水平明顯增加,約為胚胎時期的2倍。7~21日齡,IL-2基因mRNA表達水平持續顯著增加(P<0.05),到21日齡時其表達水平是胚胎時期的60倍左右,達到高峰。35日齡時雖有所下降,但56,90日齡時又上升至接近21日齡時的表達水平。
由圖1B可見,喉RALT中IL-2的mRNA表達水平在胚胎時期和出殼后1日齡時均較低,4日齡時增加至胚胎20日齡表達水平的3倍,14日齡時,其表達水平較之前日齡顯著增加(P<0.05),達到峰值。隨后的21,35日齡較14日齡的表達水平下降,56,90日齡時雖有所增加,但是仍然低于14日齡。
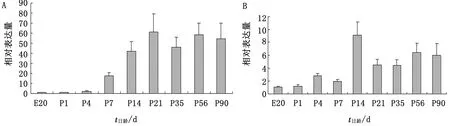
A.肺臟;B.喉圖1 不同發育階段雞RALT中IL-2基因mRNA表達水平的動態變化
本試驗在所有日齡組雞的氣管中均未檢測到IL-2 mRNA的表達。
2.2 IFN-γ基因mRNA表達水平的動態變化由圖2A可見,胚胎時期肺臟中IFN-γ mRNA的表達水平較低,1,7日齡時較胚胎時期增加10~20倍,14日齡時又比1日齡高出近35倍,但21日齡時又比14日齡顯著下降(P<0.05),此后日齡的表達水平又迅速增加,尤其是56日齡比14日齡高出3倍左右。
由圖2B可見,喉中IFN-γ基因mRNA表達水平在胚胎期和出殼后1日齡時均較低,4日齡時比胚胎20日齡顯著增加了13倍(P<0.05),7日齡時明顯下降,但14日齡又增加至與4日齡接近的表達水平,此后至35日齡,表達水平無明顯變化。56日齡時,其表達水平突然增加至4日齡時的20倍左右,而90日齡時又顯著下降(P<0.05)。
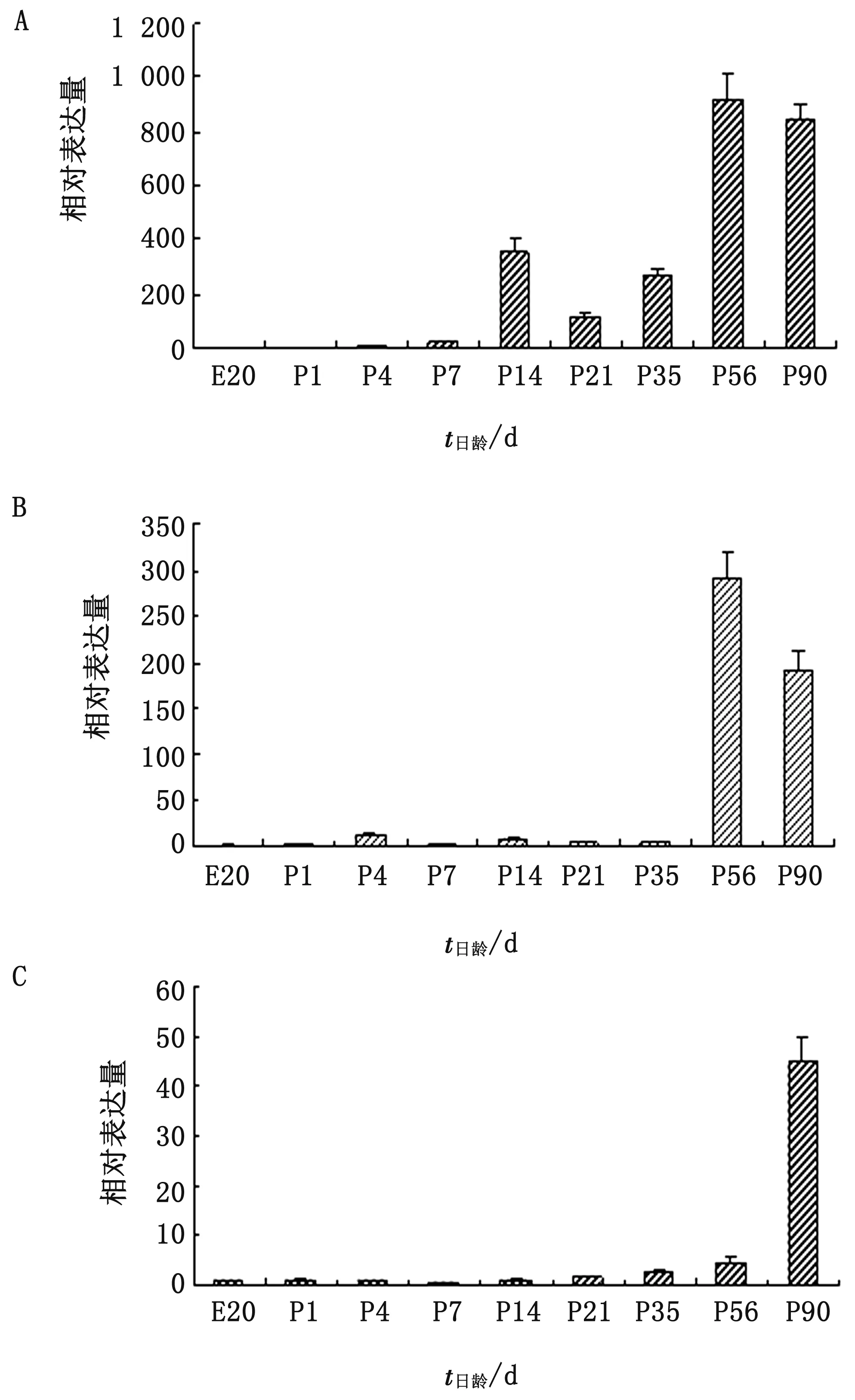
A.肺臟;B.喉;C.氣管圖2 不同發育階段雞RALT中IFN-γ基因mRNA表達水平的動態變化
由2C可見,胚胎時期一直到出殼后14日齡,氣管中IFN-γ的表達水平均比較接近,且表達量都很低,21~56日齡時,表達水平逐漸增加,但是變化幅度較小,56日齡時,表達水平大概是胚胎20日齡時的4倍左右,但90日齡時,表達水平迅速增長至43倍左右。
2.3 不同器官中IL-2和IFN-γ mRNA表達水平變化規律的比較由圖3A可見,鼻、喉和肺臟中IL-2 mRNA表達水平隨著日齡增長,整體變化趨勢相似,均隨著日齡增長表達水平逐漸增加,并在21日齡時基本穩定。其中,各日齡鼻和喉中IL-2 mRNA表達水平比較接近,且表達水平均較低,隨日齡增長,變化幅度較小,而肺臟中IL-2 mRNA表達水平從7日齡開始就顯著高于鼻和喉,21日齡時表達水平比鼻和喉高10倍左右。
由圖3B可見,各日齡鼻、喉、氣管和肺臟中IFN-γ mRNA表達水平變化趨勢與IL-2相似,隨著日齡增長表達水平逐漸增加,且4種器官中以肺臟表達水平最高,而氣管最低,鼻和喉接近,并在56,90日齡時達到較高表達水平,但仍顯著低于肺臟。
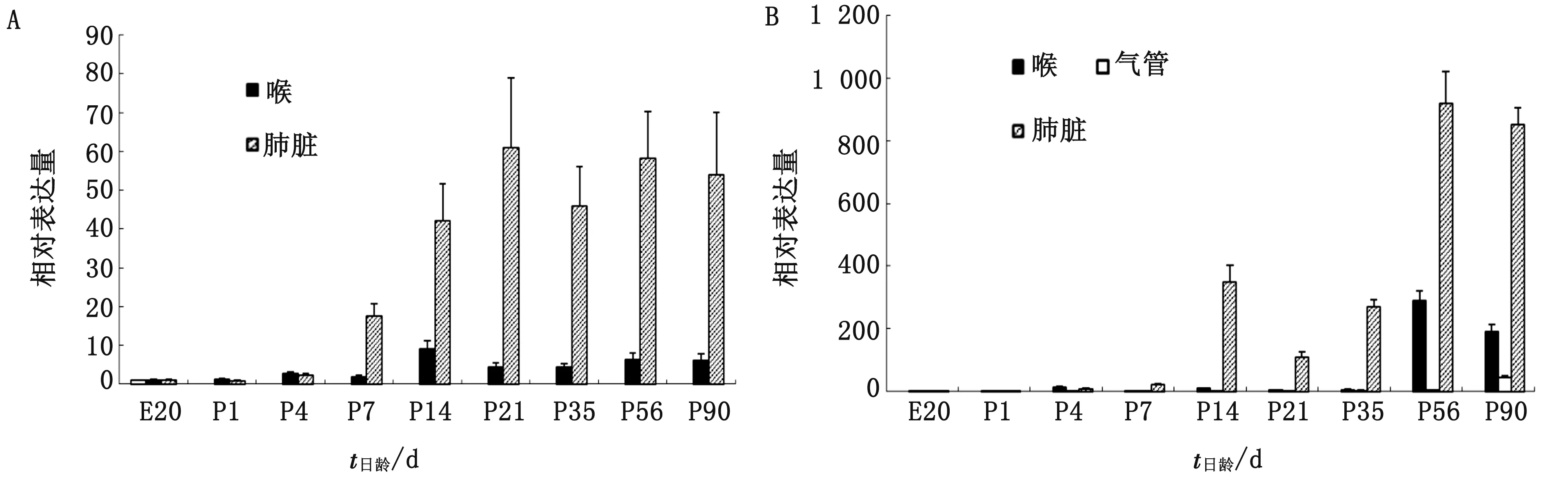
A.IL-2;B.IFN-γ圖3 不同器官中IL-2和IFN-γ mRNA表達水平的變化規律
2.4 不同發育階段雞同一RALT中IL-2和IFN-γ基因mRNA表達水平的比較由圖4A可見,胚胎20胚齡和出殼后1~7日齡時肺臟中IL-2和IFN-γ基因mRNA表達水平均比較接近,但從14~90日齡,各時期IFN-γ基因mRNA表達水平均遠高于IL-2,特別是14,56,90日齡IFN-γ基因mRNA表達水平分別高于IL-2 10,30倍左右。
由圖4B可見,除4日齡時IFN-γ基因mRNA表達水平顯著高于IL-2 4倍左右外,其他各個時期喉RALT中的IL-2和IFN-γ基因mRNA表達水平均比較接近。此外,由于56日齡時IFN-γ的增加倍數太高,如果與其他日齡列在同一圖中,會影響IL-2和IFN-γ的直觀比較,故56日齡并沒有反映到圖4B中,但實際56,90日齡時IFN-γ基因mRNA表達水平高于IL-2近48倍。
由于本試驗在所有日齡組雞的氣管中均未檢測到IL-2 mRNA的表達,所以無法與IFN-γ基因mRNA表達水平進行比較。
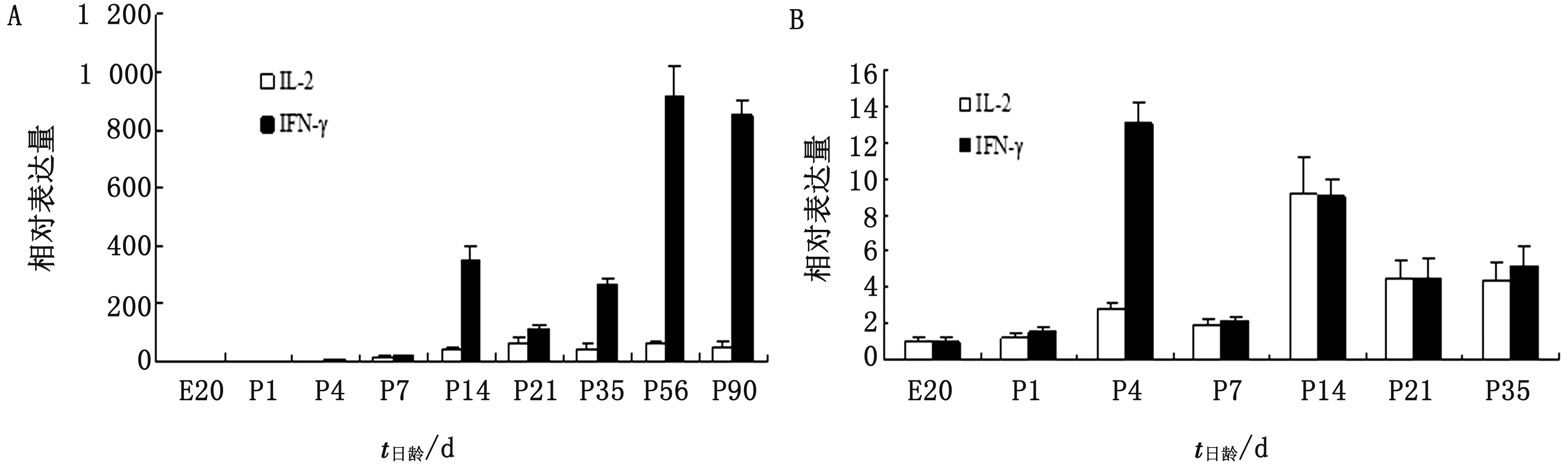
A.肺臟;B.喉圖4 不同發育階段雞RALT中IL-2和IFN-γ基因mRNA表達水平的比較
3 討論
目前,大多數關于呼吸道免疫系統的認識都是基于形態學的研究,對其免疫功能的研究在很大程度上也僅限于闡明抗原特異性抗體的反應,而對于正常雞呼吸道淋巴組織中細胞因子的分泌和表達研究較少。因此,本研究采用實時熒光定量PCR技術,檢測不同發育時期雞呼吸道淋巴組織中Th1細胞因子IL-2和IFN-γ的mRNA表達情況,從分子水平闡述雞呼吸道淋巴組織免疫功能的建立,從而了解不同時期雞呼吸道的免疫狀態。
IL-2是最重要的細胞因子之一,具有廣譜的免疫增強活性,主要由Th1細胞產生,可誘導T、B淋巴細胞的增殖分化,并可促進NK細胞的功能,能抑制Th2型細胞發育,選擇性增強Th1型細胞分化增殖,具有顯著的抗腫瘤、抗病毒的作用。IFN-γ是由活化的T淋巴細胞和自然殺傷細胞等分泌的一種低分子可溶性糖蛋白[5]。IFN-γ是由活化的NK細胞和T淋巴細胞等分泌的一種低分子可溶性糖蛋白,可以激活血管內皮細胞、NK 細胞和巨噬細胞,能直接作用于T、B淋巴細胞,促進分化。此外,IFN-γ是一種免疫調節的多效性因子,哺乳動物和家禽IFN-γ被用作感染機體的細胞介導免疫的指示器[6]。本研究通過對不同發育時期執行并調節細胞免疫功能的關鍵細胞因子IL-2和IFN-γ表達水平的檢測,發現胚胎時期呼吸道淋巴組織中細胞因子的表達水平極低,即可以認為不表達,但在出殼后隨著生長發育,其表達水平逐漸增加,并且在7,14日齡時迅速增加至較高水平,并在21日齡時基本達到峰值,且各時期IFN-γ的表達水平都遠高于IL-2,這說明,隨著生長發育呼吸道淋巴組織的細胞免疫功能逐漸增強,在出殼初期的1,2周內增加尤為顯著,并且在21日齡時基本達到成熟水平,另外IFN-γ將在呼吸道淋巴組織執行細胞免疫功能的過程中發揮主要作用。
CD4+輔助性T細胞主要包括Th1和Th2 2個亞群,Th1細胞對樹突狀細胞(DC)呈遞的抗原和B細胞通過共刺激分子呈遞的抗原應答性最好,DC通過分泌IL-12和IL-18刺激Th1細胞分泌IL-2、IFN-γ、TNF-α和TNF-β[7]。活化的輔助性T細胞通過免疫突觸高度定向地分泌IL-2和IFN-γ。Th1型細胞分泌的細胞因子包括IFN-γ和IL-2等,與CD8+細胞毒性T細胞( CTL) 的增殖、分化和成熟有關,能促進細胞免疫應答,介導遲發型超敏反應,抵抗胞內病原菌和病毒感染[8-10]。本研究對喉、氣管和肺臟3種器官中IL-2和IFN-γ mRNA表達水平的比較發現,肺臟的表達水平遠高于其他器官。有研究表明肺臟在胚胎期就出現較多抗原呈遞細胞-MHC-Ⅱ+細胞[11],由于表達MHC-Ⅱ+分子的DC細胞能夠促進IL-2和IFN-γ的分泌,所以肺臟中有較高水平的IL-2和IFN-γ mRNA表達水平。另外課題組前期研究發現肺臟在35日齡之前T淋巴細胞以CD4+細胞為主,而之后以CD8+細胞為主[12],這也與本研究所觀察到的IL-2和IFN-γ mRNA表達水平逐漸增加相關,即CD4+細胞數量的增加能夠分泌更多的IL-2和IFN-γ,而IL-2和IFN-γ又能不斷促進CD8+細胞的增殖和分化,因此CD8+細胞逐漸增多,并在35日齡時數目超過CD4+細胞。
綜上所述,本研究結果表明隨著生長發育RALT的細胞免疫功能逐漸增強,且在21日齡時基本達到成熟水平。另外,各時期IFN-γ的表達水平均遠高于IL-2,說明IFN-γ將在呼吸道淋巴組織執行細胞免疫功能的過程中發揮主要作用。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
