MrgD通過抑制內質網應激調節肝癌細胞脂肪酸代謝
2021-10-10 06:38:22區婷婷曾楠杜弢
嶺南現代臨床外科 2021年4期
關鍵詞:肝癌
區婷婷,曾楠,杜弢
細胞內的脂肪代謝對于維持細胞正常的能量代謝和細胞內環境穩態發揮著不可忽視的作用,腫瘤細胞異常增殖所需能量供應除了增加葡萄糖攝入消耗供能外,也可通過增加脂肪代謝供能[1]。有研究發現減少脂肪積累或者減弱脂質代謝可影響前列腺細胞的癌變進程[2],另有研究指出脂肪代謝與腫瘤細胞耐藥和化療不敏感度有關[3,4],這些研究提示靶向脂肪酸合成可能成為腫瘤治療的新策略[5]。
最近的一項研究發現抑制脂肪酸和膽固醇的合成可以有效降低肝癌的發生[6]。內質網應激(endoplasmic reticulum stress,ERS)在肝細胞脂肪變性和肝癌進展中發揮重要作用,許多疾病如高脂血癥、炎癥、病毒等可干擾肝細胞ER的體內穩態,進而導致肝脂質代謝失調與肝細胞的炎性壞死、凋亡,促進肝臟疾病的發生發展[7]。腎素-血管緊張素系統(renin-angiotensin system,RAS)中ACE2/Ang-(1-7)/Mas軸對維持機體肝臟脂代謝的穩態非常重要[8,9]。其中Mas受體可緩解ERS誘導劑毒胡蘿卜素誘導的HepG2細胞的ERS,從而減少HepG2細胞內甘油三酯的積累[10]。Mas受體相關G蛋白偶聯受體D(MAS-related G-protein Coupled Receptor member D,MrgD),又稱為MrgprD或TGR7,與Mas受體具有一定比例的同源性。這兩個受體對機體的血管與心臟等發揮類似的保護作用,如舒張血管[11,12]、抗心肌纖維化、抗心室肥大[13,14]等。目前,關于MrgD對肝癌細胞脂肪代謝方面的作用尚不清楚,本研究將初步探討MrgD調控肝癌細胞脂肪代謝和ERS的作用。
1 材料與方法
1.1 細胞培養
HepG2及其穩定轉染細胞株在含10%FBS的低糖DMEM培養基,置于37℃、5%CO2培養箱中培養。
1.2 構建過表達與干擾MrgD的穩定HepG2細胞株
將pcDNA3-MRGD-EGFP表達質粒質粒(Addgene)的人源MrgD全長序列克隆插入CMV-eGFPPGK-Puro表達載體質粒中的eGFP,獲得MrgD表達載體PGMLV-CMV-H_MRGD-eGFP-PGK-Puro。同時,設計針對人源MrgD序列干擾的MrgD-shRNA質粒,插入PGMLV-SC5載體,構建MrgD干擾載體H_MRGD-shRNA1(PGMLV-SC5)。上述質粒載體均帶有嘌呤霉素篩選抗性。用構建的慢病毒載體與包裝質粒共轉染293T細胞,收集病毒原液,超濾濃縮并測定病毒滴度。用包裝好的慢病毒以及慢病毒陰性對照感染HepG2細胞并用嘌呤霉素篩選,得到目的轉染穩定細胞株,qRT-PCR評估MrgD表達變化。

表1 MrgD shRNA oligo序列
1.3 HepG2細胞脂肪變性模型的構建與驗證
1.3.1 在1%BSA低糖DMEM培養基中,以油酸和棕櫚酸按一定比例配置200 μM游離脂肪酸(Free fatty acids,FFA),用于誘導細胞脂肪變性模型。
1.3.2 選用生長狀態良好的對數生長期的HepG2細胞,胰酶消化、離心、重懸后,計數按5.0×104/孔的密度鋪于24孔板(6孔板則按1.5×105/孔種板),隔夜培養細胞穩定貼壁后,細胞密度約70%,對照組每孔加入500 μL(6孔板按2 mL/孔)含1% BSA的低糖DMEM培養基,模型組每孔加入等體積的200 μM FFA高脂培養基(由1% BSA的培養基稀釋),繼續培養12 h。
1.3.3 油紅O染色觀察細胞脂質沉積:油紅O粉末(Sigma),以異丙醇溶解配置0.5%油紅O母液,與ddH2O按3∶2比例混勻后過濾配置工作液,按試劑標準步驟對4%多聚甲醛固定后的細胞進行染色觀察。
1.4 Western Blot法檢測相關蛋白表達
提取細胞蛋白進行蛋白濃度的測定及調配后,以10%SDS-PAGE凝膠進行電泳后轉膜。用一抗稀釋液按1∶1000分別稀釋相如下的抗體:并下調固醇調節元件結合蛋白2(Sterol regulatory element binding protein 2,SREBP2)(CST)、乙酰輔酶A羧化酶(Acetyl CoA carboxylase,ACC1)(CST)、免疫球蛋白重鏈結合蛋白(immunoglobulin heavy chain binding protein,Bip)(CST)、GAPDH(Santa),置4℃緩慢搖床中孵育過夜,用含5%脫脂牛奶的1×TBST按照1∶5000的比例稀釋二抗;ECL法顯影,用Image J軟件各目的條帶的灰度值進行分析。
1.5 qRT-PCR檢測相關mRNA表達
Trizol法提取細胞總RNA,參照TaKaRa RR036A逆轉錄試盒說明書,將細胞RNA逆轉錄為cDNA。qRT-PCR參照TaKaRa RR820A(TB Green Premix Ex TaqⅡ)說明書,使用Roche LightCycler?480Ⅱ進行qRT-PCR程序。通過內參基因的Ct值計算出目的基因的相對表達量。以Folds=2-ΔΔCt公式比較目的基因的表達差異,ΔΔCt=(Ct目的基因-Ct內參基因)處理組-(Ct目的基因-Ct內參基因)對照組。qRT-PCR引物序列:H_MRGPRD-F:GCTGTTCGTGGTGGTCCTG;H_MRGPRD-R:GGAGAGGCGTGACAAGCTG;H_GAPDH-F:GTCTCCTCTGACTTCAACAGCG;H_GAPDH-R:ACCACCCTGTTGCTGTAGCCAA。
1.6 細胞免疫熒光
對數生長期的細胞消化重懸后計數,按8×104/孔于24孔板中均勻種板,細胞貼壁后當細胞達到約40%匯合度進行免疫熒光觀察。MrgD一抗和二抗Cy3均按1∶500比例稀釋;按標準步驟進行抗體孵育后,在熒光顯微鏡下,選擇抗體對應的激發光源,觀察、采集圖像。
1.7 統計學分析
采用SPSS 2.0統計軟件進行分析,實驗數據采用均數±標準誤(±s),采用GraphPad Prism 9.0軟件對數據進行統計、作圖,多組間比較采用單因素方差分析。以P<0.05為差異具有統計學意義。
2 結果
2.1 HepG2的MrgD過表達與干擾穩定株的構建與鑒定
對構建的人源MrgD表達載體PGMLV-CMVH_MRGD-eGFP-PGK-Puro和含有MrgD序列干擾的MrgD-shRNA質粒測序鑒定正確后,構建的慢病毒載體轉染HepG2細胞,得到穩定表達細胞株,通過qRT-PCR與Western Blot分別檢測細胞株MrgD的mRNA與蛋白水平。qRT-PCR(圖1A)結果顯示過表達細胞MrgD的mRNA水平較對照細胞顯著上升(P<0.001),4組shRNA干擾組細胞MrgD的mRNA水平較其對照組明顯降低(P<0.001),其中shRNA3最低,后續試驗均選用感染shRNA3序列細胞株;免疫熒光顯示的MrgD蛋白表達有同樣的趨勢(圖1B)。
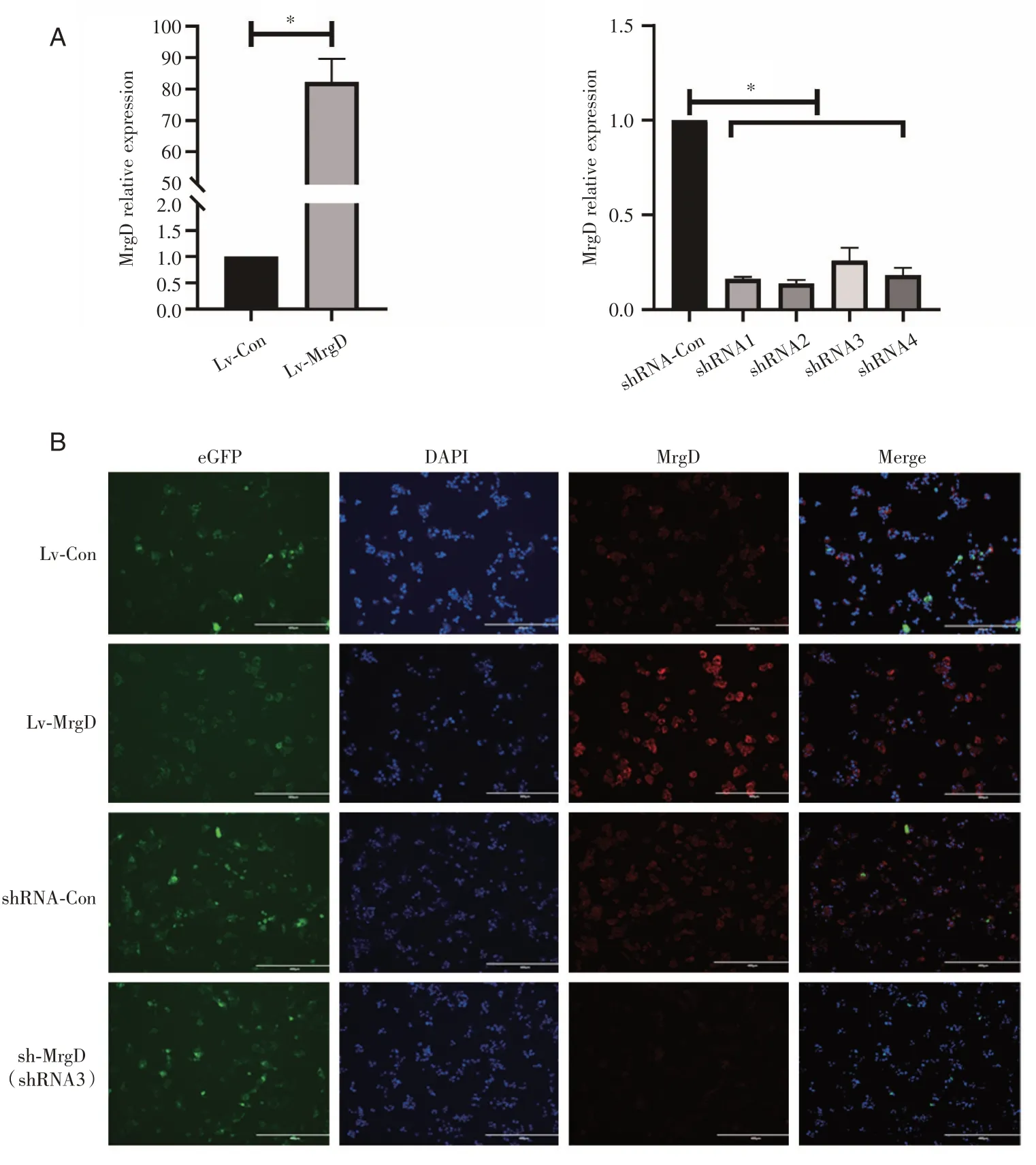
圖1 過表達和低表達MrgD的HepG2穩定株的建立與鑒定A:qRT-PCR檢測細胞MrgD mRNA表達水平,以GAPDH為內參;B:免疫熒光觀察細胞MrgD蛋白表達。Lv-Con:過表達對照組;Lv-MrgD:MrgD過表達組;Sh-Con:干擾對照組;Sh-MrgD:MrgD干擾組;*P<0.001。
2.2 體外脂肪變性模型的構建
采用含200 μM的由棕櫚酸(palmitic acid,PA)和油酸(oleic acid,OA)混合的FFA脂性培養基刺激HepG2細胞12 h。設置4個分組:對照組(1%BSA處理)以及OA∶PA比例分別為1∶2、1∶1、2∶1的FFA處理組,之后用MTS測定各組細胞的活性。結果顯示200 μM的FFA(OA∶PA=2∶1)處理細胞12 h不影響細胞活性(圖2A),故選用該方案誘導細胞脂肪變性并進行油紅O染色。結果顯示,與1%BSA培養基處理的對照組對比,FFA處理的HepG2細胞內的橘紅色脂滴顯著增加(圖2B)。
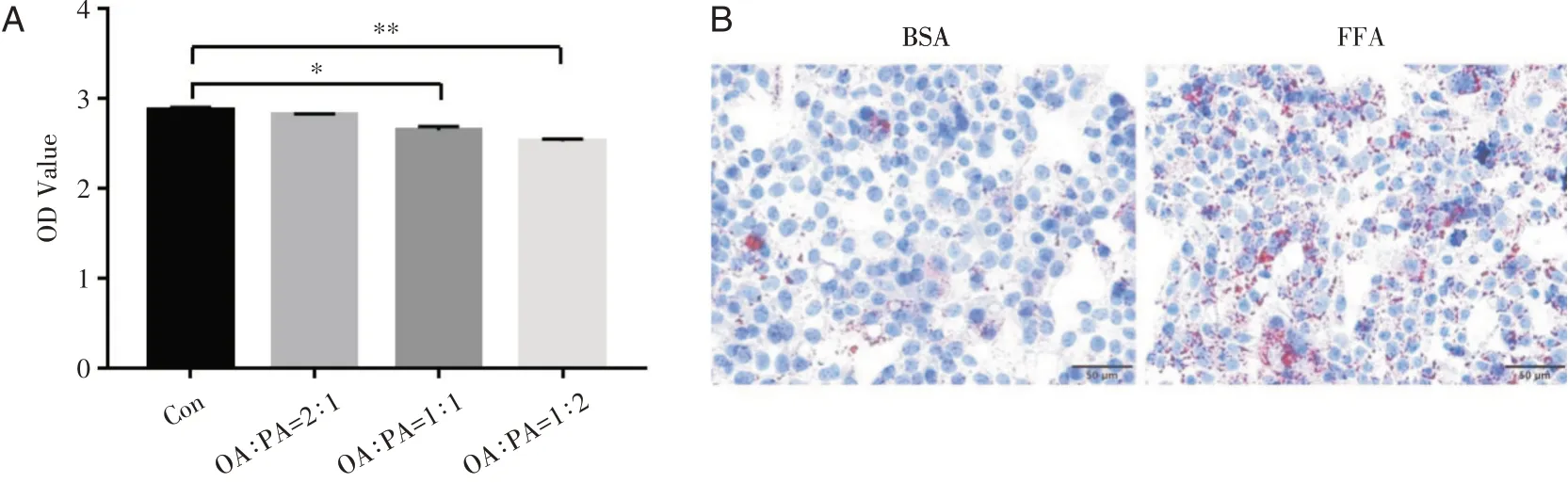
圖2 FFA誘導HepG2細胞脂滴堆積分別用含1%BSA(模型對照組)與含200 μM FFA的低糖DMEM培養基(模型組)培養HepG2細胞12 h A:MTS檢測200 μM的FFA(OA∶PA=1∶2、1∶1、2∶1)的處理HepG2細胞12 h的活性。*P<0.05,**P<0.01。B:200 μM的FFA(OA∶PA=2∶1)誘導細胞12 h的油紅O染色。
2.3 MrgD改善FFA誘導的肝細胞脂肪變性與內質網應激
為探究MrgD是否對肝臟脂肪代謝有影響,通過油紅O染色觀察MrgD過表達對FFA誘導后細胞內脂滴沉積含量的影響,Western blot檢測脂肪合成相關蛋白(ACC1)以及ERS標記蛋白(BiP、ATF4)水平。結果顯示,在HepG2脂肪變性模型中,MrgD過表達明顯抑制了FFA誘導的顯著增加的脂滴蓄積(圖3A);FFA誘導后,對照組細胞脂肪合成標記蛋白SREBP2、ACC1和ERS標記蛋白BiP水平上調,而過表達MrgD下調了上述蛋白的表達(圖3B)。
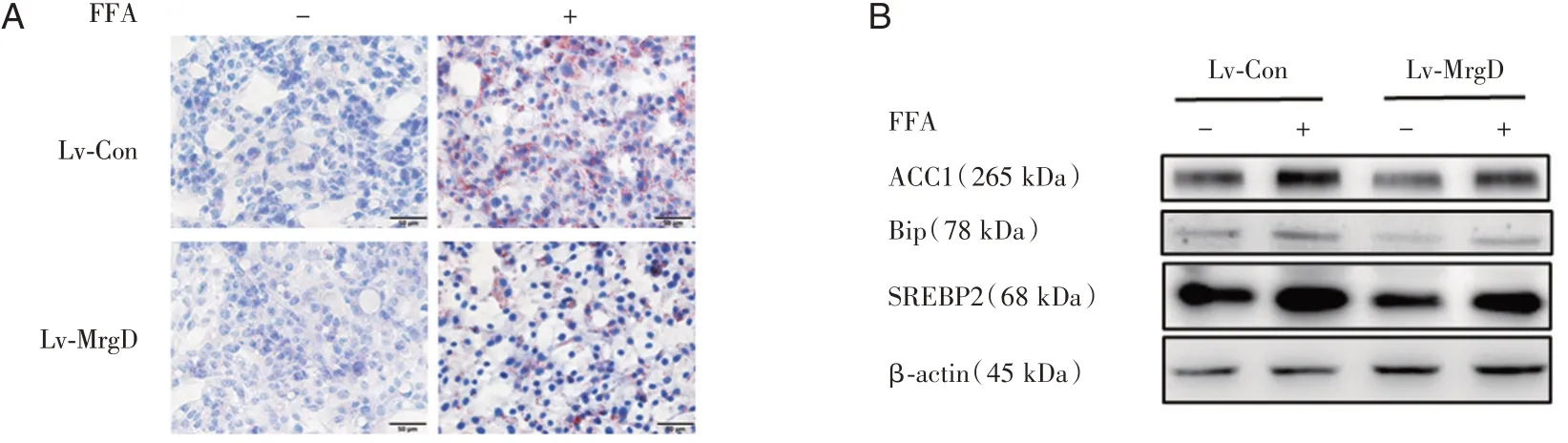
圖3 MrgD對肝細胞脂肪變性的影響用200 μM的FFA誘導MrgD過表達組及其對照12 h,進行后續實驗的驗證A:油紅O染色觀察各組脂滴沉積情況;B:Western Blot檢測過表達組的脂肪合成相關蛋白(ACC1、SREBP2)與ERS標記蛋白(BiP)水平。
2.4 干擾MrgD促進FFA誘導的肝細胞脂肪變性與內質網應激
同時,MrgD干擾HepG2細胞脂肪變性后,油紅O染色發現,相對于對照組,MrgD干擾后FFA誘導的脂滴合成顯著增加(圖5A);相應的脂肪合成標記蛋白SREBP2、ACC1表達水平顯著上調,同時ERS標記蛋白BiP表達水平增高(圖5B)。
3 討論
脂質代謝失調和炎癥是癌細胞的常見表型[15],細胞內脂肪酸作為膜脂、信號分子和修飾基團的生物合成前體為腫瘤生長提供能量儲存[16]。脂質合成、攝取和儲存通常在肝癌發生中上調,靶向脂質代謝可能是一種有前途的癌癥治療方法[17]。本研究發現在肝臟腫瘤細胞株HepG2中上調MrgD表達可通過抑制脂肪合成、改善ERS途徑顯著改善肝癌細胞脂肪變性,研究提示MrgD相關通路可能成為研究肝癌防治策略的新方向。
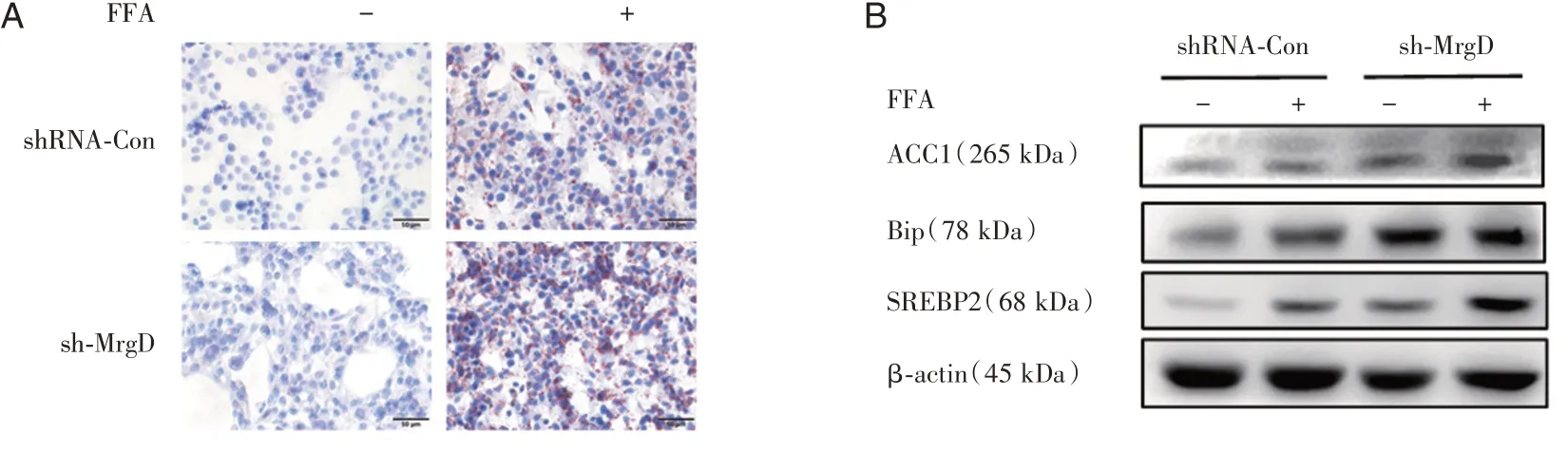
圖4 干擾MrgD對肝細胞脂肪變性的影響A:經FFA誘導后各組的油紅O染色;B:Western Blot檢測FFA誘導后各組的脂肪合成相關蛋白(ACC1、SREBP2)與ERS標記蛋白(BiP)。
FFA常用于誘導建立脂肪變性細胞模型[18,19],油酸(OA)和棕櫚酸(PA)是體內常見的FFA,可合成甘油三酯并儲存于肝細胞胞質中。體內甘油三酯含量過高會導致肝細胞脂肪變性,FFA能激活內質網應激的3條信號通路[20]。ERS的激活可導致肝脂質代謝失調與肝細胞的炎性壞死、凋亡,在肝細胞脂肪變性和肝癌進展中發揮重要作用[7]。本研究應用200 μM的FFA(OA∶PA=2∶1)誘導HepG2細胞脂肪變性,造模后肝細胞ERS及脂肪合成相關基因的mRNA和蛋白水平顯著升高。
RAS對維持機體肝臟脂代謝的穩態非常重要,Mas受體是其中關鍵的一員,Mas受體敲除小鼠表現出肝臟脂質蓄積增強和脂肪合成相關蛋白上調,而過表達Mas受體則會扭轉上述變化趨勢[21]。MrgD受體是與Mas受體有一定比例同源性,與Mas受體發揮許多相似的生理作用,但其MrgD在肝細胞脂代謝中的作用尚不清楚。本研究通過構建FFA誘導的MrgD受體過表達與干擾的HepG2細胞脂肪變性模型,發現高表達MrgD明顯降低FFA誘導的胞質內脂質含量,并下調固醇調節元件結合蛋白2(SREBP2)和乙酰輔酶A羧化酶1(ACC1)。SREBP2是參與脂質合成的調控關鍵轉錄因子。已有研究表明[22,23],SREBP2表達受AdipoR1/AMPK/SIRT1信號通路調控的,AMPK激活后也可直接作用于SREBP2,使其磷酸化,進而下調SREBP2的靶基因,如參與膽固醇和脂肪酸生物合成等相關基因的轉錄。ACC1同樣在脂肪酸合成中起主要作用。ACC是一類生物素羧化酶,其催化乙酰輔酶A和碳酸酯的ATP依賴性縮合以形成丙二酰輔酶A,后者是從頭脂肪生成的必需和限速底物,也是肝臟組織細胞中長鏈脂肪酸β-氧化的關鍵調節酶CPT-1(Carnitine palmitoyltransferase 1,肉毒堿-棕櫚酰轉移酶1)的變構抑制劑[24]。
此外,本研究發現高表達MrgD可抑制免疫球蛋白重鏈結合蛋白(Bip),同時在低表達MrgD細胞中觀察到相反的結果。BiP是一種高度保守的分子伴侶,它充當Ca2+的調節劑協助蛋白質轉運到內質網,并通過其兩個結構域核苷酸結合域和底物結合域協助內質網中蛋白質的折疊過程。BiP作為維持內質網功能穩態的重要調節劑,可激活涉及未折疊蛋白反應的基因的轉錄,也是ERS的主要標記物[25]。已有的研究表明,FFA作用肝細胞后,脂肪合成基因SREBP-1c和BiP的mRNA和蛋白水平以及脂肪酸合成酶mRNA顯著增加,參與肝細胞脂肪變性過程[26]。這些研究結果提示MrgD可能通過調控ERS參與調節肝癌細胞脂肪合成。
綜上,本研究在體外初步揭示了MrgD通過調控內質網應激抑制肝癌細胞脂肪合成。調節肝細胞的脂質代謝異常及緩解ERS的機制,從而阻止或延緩肝臟脂肪變性和肝癌的發生發展,有望成為肝癌防治的一個有效方法,本研究有助于深入研究MrgD通路在肝癌細胞脂肪變性中的作用和機制,探討其在肝癌防治中的潛在價值。
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
