馬身豬CD63基因CDS區(qū)的克隆、表達及生物信息學分析
2021-09-14 08:12:36張雪蓮晉大鵬吳怡琦李文霞秦本源蔡春波高鵬飛郭曉紅李步高曹果清
家畜生態(tài)學報 2021年8期
張雪蓮,晉大鵬,劉 敏,吳怡琦,李文霞,秦本源, 蔡春波,高鵬飛,郭曉紅,李步高,曹果清*
(1.山西農(nóng)業(yè)大學 動物科學學院,山西 太谷 030801;2. 中國農(nóng)業(yè)科學院 北京畜牧獸醫(yī)研究所,北京 100193)
抗原分化簇63(cluster of differentiation 63, CD63)作為最具有特征性的血小板活化分子標志物[1],被發(fā)現(xiàn)于激活的血小板表面,命名為血小板糖蛋白40(platelet glycoprotein 40, Plt Gp40)[2]。隨后研究發(fā)現(xiàn),CD63是存在于人早期黑色素瘤細胞中的黑色素瘤抗原491(ME491)[3],還與溶酶體相關的膜表面糖蛋白3(lysosome-associated membrane glycoprotein 3, LAMP3)是同一種分子[4]。CD63作為四跨膜蛋白超家族(transmembrane 4 superfamily, TM4SF)成員,包含4個跨膜域與2個胞外域,具有相似的結構和功能。
CD63與細胞活化、黏附、變異、炎癥反應、人免疫缺陷病毒(HIV)、生殖系統(tǒng)及腫瘤的侵襲、轉移等多種生命過程相關[5-6]。CD63作為嗜堿性粒細胞的活化標志,廣泛表達于活化的嗜堿性粒細胞膜表面;此外在其他活化細胞表面也有表達,包括血小板、內(nèi)皮細胞、淋巴細胞、單核細胞、中性粒細胞、樹突狀細胞、巨噬細胞和組織肥大細胞等[7]。Miyamoto等[8]發(fā)現(xiàn),在活化的小鼠巨噬細胞中,CD63 mRNA表達顯著增加。CD63作為活化血小板表面的抗體,可介導血小板的活化過程,內(nèi)皮細胞與中心粒細胞結合后,參與炎癥反應及腫瘤轉移等病理過程[9-10]。此外CD63在免疫應答方面也發(fā)揮了重要作用,在巨噬細胞(APC)和樹突狀細胞(DC)中,CD63可通過內(nèi)體途徑伴隨主要組織相容性復合體II(Major Histocompatibility Complex II, MHC-II)分子呈遞抗原并啟動免疫應答,即CD63可以與細胞表面的MHC-II相互作用,形成的CD63-MHC-II類復合物重新分布在細胞表面以呈遞給CD4陽性T細胞從而促進免疫應答的發(fā)生[11-12],并起到保護膜蛋白免于降解的作用[13-14];CD63通過協(xié)調(diào)核內(nèi)體和自噬過程以調(diào)節(jié)細胞內(nèi)潛伏期膜蛋白1(Latent membrane protein 1, LMP1)水平[15]。而當細胞受到病原菌刺激活化后,CD63從反面高爾基網(wǎng)(trans Golginetwork, TGN)轉運至細胞表面,或者直接通過細胞內(nèi)途徑轉運至內(nèi)體和溶酶體,參與多種生理過程。劉艷杰等[16]研究表明,CD63在免疫應答和病原菌侵襲中發(fā)揮重要作用,這為進一步研究豬免疫應答分子調(diào)控網(wǎng)絡提供了一定的理論基礎。
馬身豬是山西省地方豬種,具有產(chǎn)仔多、耐粗飼、繁殖力高、抗逆性強、肌內(nèi)脂肪含量高、肉質(zhì)品質(zhì)好等優(yōu)點。近幾年關于馬身豬的研究多集中在肉質(zhì)[17-18]和脂肪沉積[19]方面,而對馬身豬免疫方面的研究相對較少。此外CD63基因的研究主要集中在人、小鼠等試驗動物上,有關豬CD63基因的研究未見報道。因此,本試驗對馬身豬CD63進行克隆及生物信息學分析,以期為進一步研究豬CD63生物學功能提供理論依據(jù)。
1 材料與方法
1.1 試驗動物和樣品采集
本研究所用試驗動物為90日齡山西省地方豬種馬身豬3頭,均為公豬,28日齡斷奶時去勢,由山西省大同市種豬場提供。達目標日齡當天屠宰,屠宰后,分別取馬身豬的心臟、肝臟、脾臟、腎臟、腰大肌、背部皮下脂肪等樣品,放入已標記的凍存管中,迅速置于液氮中保存。
1.2 主要試劑與儀器
1.2.1 主要試劑 Green Taq Mix(Vazyme,南京);DL1 000 DNA Marker、RNAiso Plus regent、Prime Script RT reagent Kit with gDNA Eraser、SYBR Premix Ex Taq II等購于大連TaKaRa。
1.2.2 主要儀器 AB-9902 PCR儀(AB,美國);Universal Hood II 核酸蛋白成像儀(Bio-Rad,美國);ND-1000核酸蛋白測定儀(Nanodrop,美國);DYY-3電泳儀(六一儀器廠,北京);ABI 7500實時熒光定量PCR儀(ABI,美國)。
1.3 試驗方法
1.3.1 總RNA提取和cDNA合成 采用TaKaRa RNAiso Plus試劑盒提取各組織的總RNA,提取方法參照試劑盒說明書。將提取的總RNA用核酸蛋白測定儀測定其純度及濃度,OD260/OD280在1.8~2.0范圍內(nèi)的總RNA用于后續(xù)研究。采用PrimeScript RT reagent Kit with gDNA Eraser試劑盒將RNA反轉錄成cDNA。合成的cDNA置于-20 ℃冰箱保存?zhèn)溆谩?/p>
1.3.2 引物的設計與合成 根據(jù)NCBI GenBank數(shù)據(jù)庫中預測的豬CD63基因mRNA序列(登錄號:XM_005663878.2),用NCBI Primer-BLAST和Oligo7軟件分段設計擴增CD63基因CDS區(qū)的引物P1及P2,并設計熒光定量引物P3及內(nèi)參基因18S rRNA的引物(表1),送上海生工股份有限公司合成。

表1 引物信息Table 1 Primer information
1.3.3CD63基因CDS區(qū)的擴增及克隆 以馬身豬所有組織的混合cDNA作為模版,擴增豬CD63基因CDS區(qū)。PCR反應總體積為20 μL:Green Taq Mix 10 μL,cDNA 2 μL,P1或P2上下游引物(10 μmol·L-1)各0.5 μL,ddH2O補至20 μL。反應程序:95 ℃預變性5 min;95 ℃變性15 s,59 ℃退火15 s,72 ℃延伸30 s,35個循環(huán);72 ℃延伸7 min。反應產(chǎn)物用1%的瓊脂糖凝膠電泳進行檢測,選擇擴增效果好的PCR產(chǎn)物送華大基因公司測序。
1.3.4 不同組織CD63基因表達特性分析 以18S rRNA為內(nèi)參基因,采用qRT-PCR技術對馬身豬各組織CD63的表達譜進行分析。qRT-PCR反應總體積為20 μL:上、下游引物(10 μmol·L-1)各0.3 μL,cDNA 2 μL,2×SYBR Premix Ex Taq II 10 μL,RNAase Free ddH2O補至20 μL。反應程序參照SYBR Premix Ex Taq II說明書:95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火30 s,45個循環(huán);95 ℃ 15 s,60 ℃ 1 min,95 ℃ 30 s制作熔解曲線。每個樣本技術重復3次。
1.3.5 CD63的生物信息學分析 測序結果采用DNAMAN軟件進行序列比對及拼接;運用NCBI在線軟件(https://www.ncbi.nlm.nih.gov/orffinder/)和(https://www. ncbi.nlm.nih.gov/Structure/cdd/ wrpsb.cgi)分析CD63基因的開放閱讀框并預測CD63蛋白保守結構域;應用PortScale(https://web.expasy.org/protscale/)和ProtParam(http://web.expasy.org/protparam/)軟件對豬CD63的理化性質(zhì)進行預測;采用NetPhos 3.1(http://www.cbs.dtu.dk/services/NetPhos/)在線預測磷酸化位點;應用NetNGlyc 1.0在線軟件(http://www.cbs.dtu.dk/services/NetNGlyc/和http://www.cbs.dtu.dk/services/DictyOGlyc)分別預測其N-糖基化位點和O-糖基化位點;運用德泰生物在線軟件(http://www.detaibio.com/tools/signal-peptide.html和http://www.detaibio. com/tools/transmembrane.html)對CD63的信號肽和跨膜結構域進行預測;同時應用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)和PredictProtein(http://www.predictprotein.org/)對豬CD63蛋白質(zhì)的二級結構進行預測;應用Phyre 2.0軟件(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)對豬CD63蛋白的三級結構進行預測;用BLAST(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE _TYPE=BlastSearch&LINK_LOC=blasthome)軟件比較豬、人、牛、摩弗倫羊、羊駝、野駱駝、家貓、抹香鯨、北大西洋小須鯨、長江江豚等12個物種CD63蛋白氨基酸序列的相似性;采用MEGA X軟件構建上述物種基于CD63氨基酸序列的進化樹。
1.4 數(shù)據(jù)分析
應用2-△△CT法分析qRT-PCR檢測結果,結果以平均值±標準誤表示。采用SPSS version 22.0軟件的單因素方差分析方法比較CD63基因在不同組織間的表達差異,用Duncan's法進行多重比較,P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結果與分析
2.1 豬CD63基因CDS區(qū)的克隆及序列分析
NCBI數(shù)據(jù)庫中豬CD63基因序列為預測序列,本試驗中未能一次性成功擴增出該基因的完整CDS區(qū)序列,所以采用分段擴增,再進行拼接并結合與預測序列比對的方法,共獲得長度為838 bp的片段,該序列已上傳至NCBI數(shù)據(jù)庫,GenBank登錄號為MN082631。圖1中各片段產(chǎn)物的大小與預期結果一致。運用NCBI上的開放閱讀框查找器分析本研究獲得的豬CD63基因序列,發(fā)現(xiàn)其含有一個長714 bp的ORF,共編碼237個氨基酸(圖2B)。
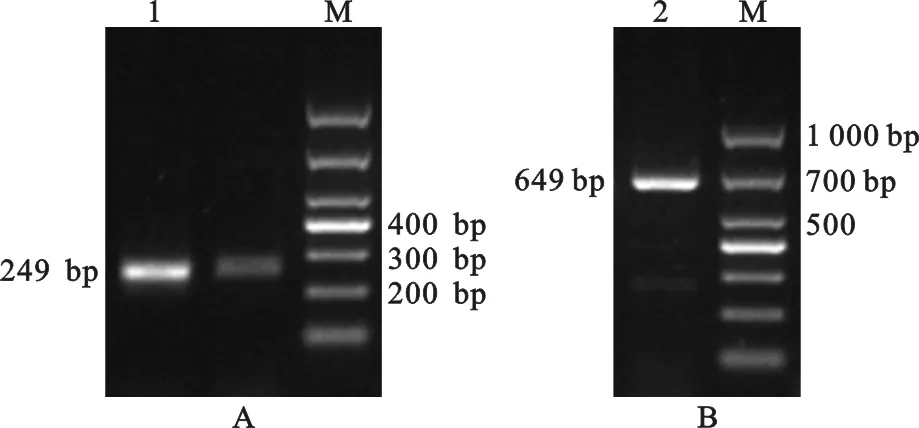
圖1 引物P1(A)及P2(B)的擴增結果 1,2. PCR產(chǎn)物;M. DL1 000 DNA MarkerFig. 1 Amplification results of primer P1 (A) and P2 (B) 1,2. PCR products;M. DL 1 000 DNA Marker

圖2 序列拼接結果(A)及預測的蛋白質(zhì)序列(B)Fig. 2 Sequence assembly result (A) and the predicted protein sequence (B)
2.2 豬CD63基因在不同組織中的表達譜
由圖3可以看出,CD63在各組織中均有表達,且在不同組織間表達量具有差異,在脂肪組織中表達量最高,其次是肝臟、肌肉、脾臟和心臟等組織,在腎臟中表達量最低。
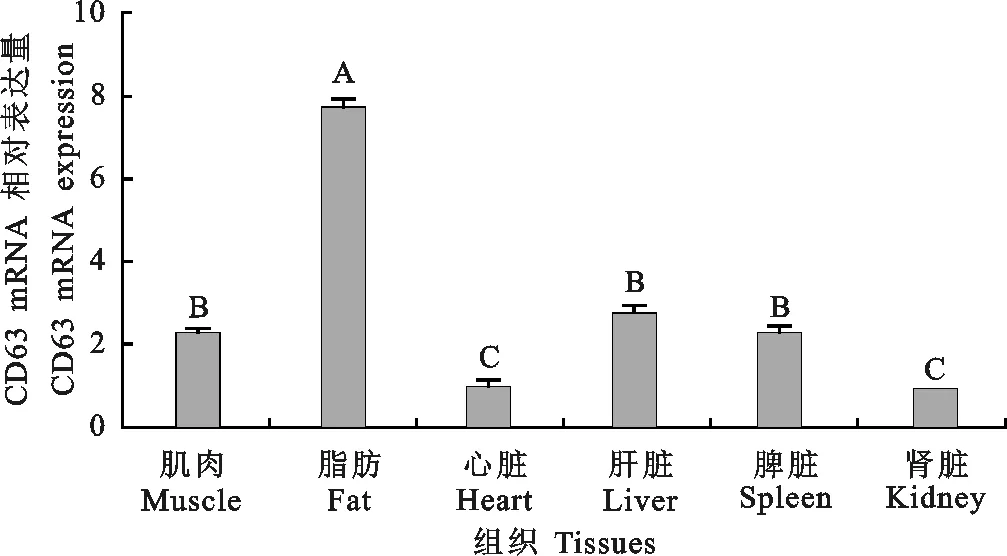
圖3 馬身豬不同組織CD63基因的表達量 不同大寫字母表示差異極顯著(P<0.01)Fig. 3 Expression patterns of CD63 in different tissues of Mashen pig Different uppercase letters indicate highly significant difference(P<0.01)
2.3 豬CD63的生物信息學分析
2.3.1 CD63理化性質(zhì)分析 用ProtParam軟件對CD63的理化性質(zhì)進行了預測,結果表明CD63由237個氨基酸組成,其分子式為C1171H1855N297O315S22,相對分子質(zhì)量為25 839.73,理論等電點(PI)為7.40,說明豬CD63由中性氨基酸組成;其中Val(12.7%)、Ala(10.5%)、Leu(10.5%)、Gly(7.2%)、Cys(6.3%)和Phe(5.9%)為主要氨基酸;CD63蛋白帶負電荷的殘基數(shù)(Asp+Glu)為17,帶正電荷的殘基數(shù)(Arg+Lys)為18;其消光系數(shù)(mol·L-1·cm-1·γ=280 nm)為26 315,不穩(wěn)定系數(shù)為24.07,并預測其在哺乳動物紅細胞中的半衰期為30 h,說明CD63蛋白較穩(wěn)定;脂肪族氨基酸指數(shù)為113.08,平均親水性為0.716。疏水性分析結果顯示,CD63蛋白的最小疏水性指數(shù)為-2.744(Position:128),最大疏水性指數(shù)為3.267(Position:56),為疏水性蛋白(圖4)。
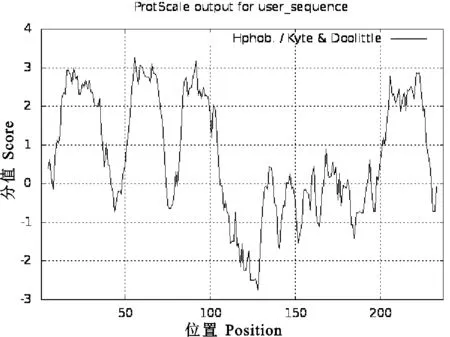
圖4 豬CD63蛋白疏水性分析Fig. 4 Hydrophobicity analysis of pig CD63 protein
信號肽和跨膜區(qū)域(圖5)及保守結構域(圖6)預測結果表明,CD63氨基酸序列不含信號肽,但是含有4次跨膜結構(13~35,50~72,85~107,205~227),N-和C-末端都位于膜的細胞內(nèi)側,其功能域預測與結構域預測相一致。CD63糖基化位點預測結果顯示(圖7),CD63有3個可能的N-糖基化位點,發(fā)生在“Asn-X-Ser/Thr”(X是除脯氨酸外的任一氨基酸,Asn為天冬酰胺,Ser為絲氨酸,Thr為蘇氨酸)的Asn殘基上,其中2個位點位于第3個與第4個跨膜結構域之間的胞外區(qū)內(nèi),另一個在第1個與第2個跨膜結構域之間的胞外區(qū)內(nèi),但沒有發(fā)現(xiàn)O-連糖基化位點。磷酸化預測結果顯示(圖8),Ser磷酸化位點有6個,分別位于第51、91、160、167、196和232位;Thr磷酸化位點有4個,分別位于第44、48、75和152位;Tyr磷酸化位點有1個,位于第80位。
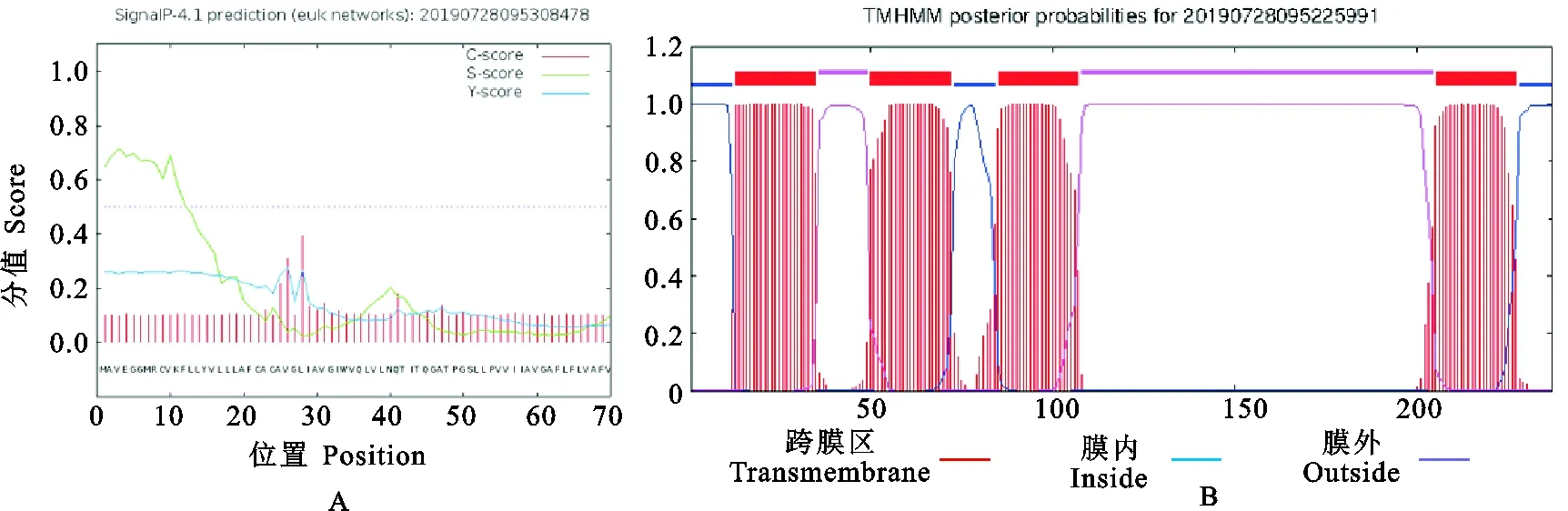
圖5 豬CD63蛋白的信號肽(A)和跨膜結構域(B)預測Fig. 5 Prediction of signal peptide (A) and transmembrane domain (B) of pig CD63 protein
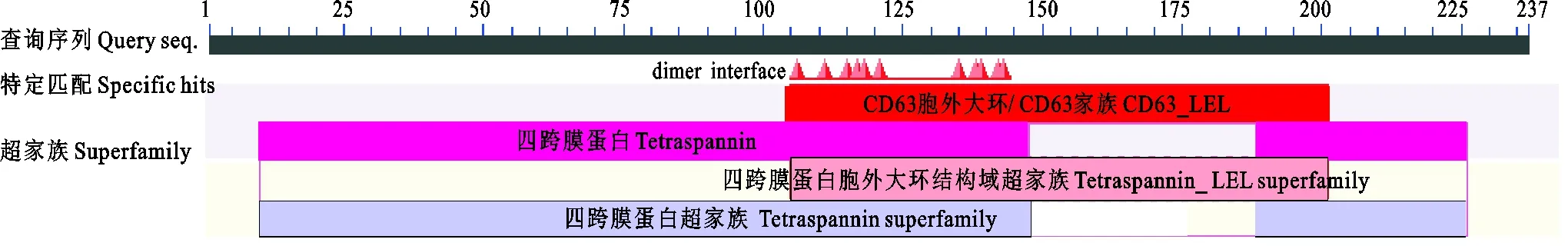
圖6 豬CD63蛋白保守結構域分析Fig. 6 Conservative domain analysis of pig CD63 protein
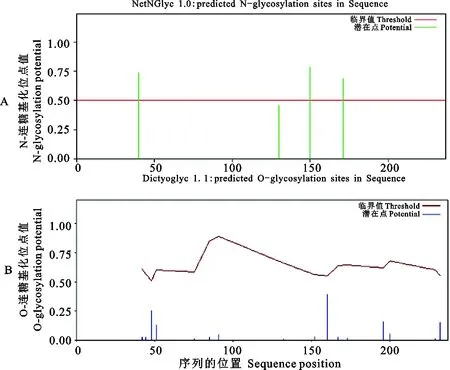
圖7 豬CD63蛋白N-連糖基化位點和O-連糖基化位點預測Fig. 7 Prediction of N-linked glycosylation site and O-linked glycosylation site of pig CD63 protein
2.3.2 豬CD63蛋白高級結構預測 同時應用PredictProtein和SOPMA軟件對豬CD63蛋白質(zhì)的二級結構進行預測,結果如圖9所示。CD63的二級結構主要由α-螺旋(h)、延伸(e)、β-折疊(t)和無規(guī)則卷曲(c)組成,分別占58.23%、11.39%、5.49%和24.89%;采用Phyre2.0軟件預測CD63蛋白三級結構(圖10),發(fā)現(xiàn)豬CD63均存在α-螺旋、β-折疊和disordred區(qū)域。
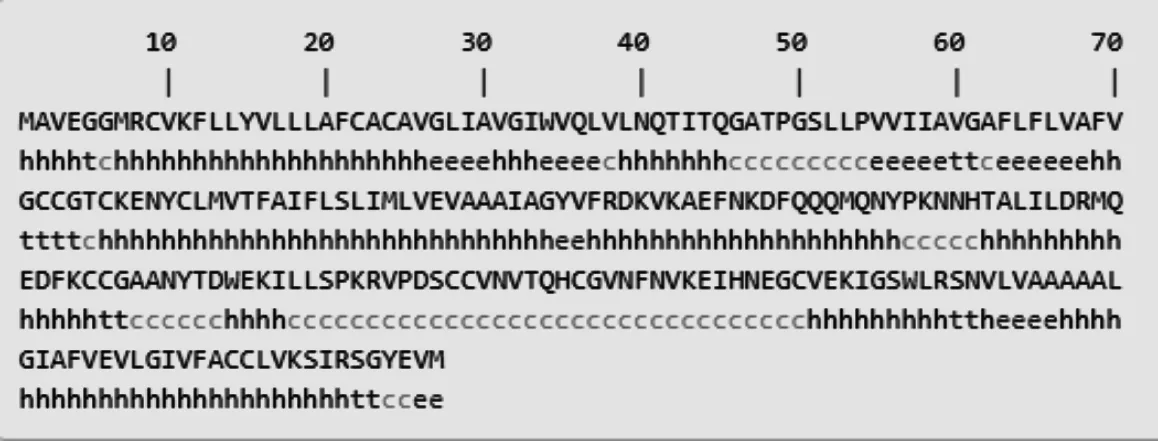
圖9 豬CD63蛋白二級結構預測Fig. 9 Prediction of secondary structure of porcine CD63 protein sequences
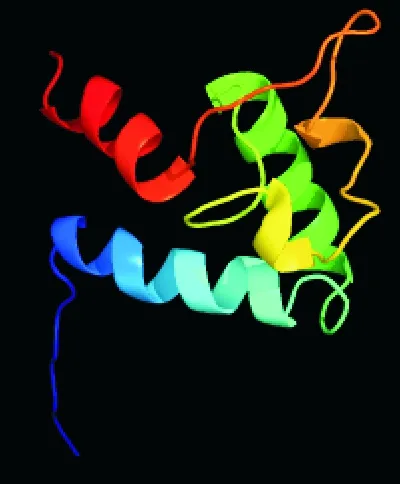
圖10 豬CD63蛋白三級結構預測Fig. 10 Prediction of tertiary structure of porcine CD63 protein
2.3.3 不同物種CD63氨基酸序列進化樹的構建 本研究獲得的馬身豬CD63氨基酸與野豬序列一致,用NCBI BLAST比較包括野豬、人、牛、長江江豚、摩弗倫羊等在內(nèi)的12個物種CD63氨基酸序列的相似性,結果表明豬的CD63與野駱駝的相似性最高,達90.72%;其次是羊駝、長江江豚、抹香鯨,相似性均達到90.30%。采用MAGE X軟件的鄰接法構建這12個物種的進化樹,如圖11所示,豬CD63與駱駝科的野駱駝和羊駝聚為一個分支,與鯨類和長江江豚較近,與其他5個物種的親緣關系比較遠,說明豬CD63可能與駱駝科間具有相似的功能。
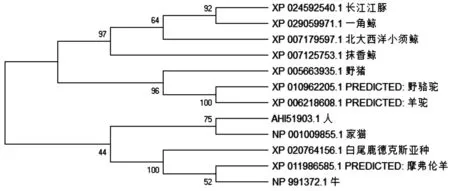
圖11 12個物種CD63蛋白系統(tǒng)進化樹Fig. 11 CD63 protein system phylogenetic tree of 12 species
3 討 論
CD63作為四跨膜蛋白家族的表面膜蛋白之一,可通過4個跨膜結構域穿過質(zhì)膜,表明這些結構域可能參與某些細胞類型的信號轉導[20]。CD63已被定性為多種細胞類型的激活或分化標記,并且已知在細胞膜中與β1整合素、主要組織相容性復合抗原和其他四跨膜蛋白緊密結合[21-23]。本研究中對豬CD63理化性質(zhì)和結構預測,與前人結果一致[16]。馬身豬的CD63是一種比較穩(wěn)定的典型的4次跨膜蛋白,兩個胞外域分別與對應的配體結合,其N末端和C末端均位于細胞內(nèi),與骨架蛋白及胞內(nèi)信號分子結合參與信號轉導。本研究中CD63蛋白消光系數(shù)為26 315,理論等電點(PI)為7.40,不穩(wěn)定系數(shù)為24.07,最小疏水性指數(shù)為-2.744(Position:128),最大疏水性指數(shù)為3.267(Position:56),屬于中性穩(wěn)定疏水蛋白,與在七鰓鰻中的研究結果相同[24]。通過相似性比對發(fā)現(xiàn),豬CD63與野駱駝的相似性最高,達90.72%,其次是羊駝、長江江豚、抹香鯨等,相似性均達到90.30%。系統(tǒng)發(fā)育樹表明,相較于哺乳動物來說,豬的CD63與駱駝科的羊駝和野駱駝親緣關系最近,其次是與鯨類和長江江豚的遺傳距離較近,符合物種進化規(guī)律。
CD63在免疫應答中發(fā)揮重要作用,除了CD63-MHC-II類復合物,在白細胞募集、吞噬及過敏等免疫反應中均具有重要地位。內(nèi)皮細胞表面的CD63能與P型選擇素相互作用調(diào)控白細胞的募集過程[25]。作為攻擊入侵病原體的主要細胞介導的防御機制,吞噬作用在哺乳動物的先天免疫反應中起重要作用。CD63在細胞內(nèi)吞和吞噬作用中起關鍵作用,有報道稱在人類樹突狀細胞中,CD63內(nèi)化的同時伴隨著釀酒酵母的吞噬[26];Yu等[27]揭示了CD63在促進血細胞介導的吞噬作用中的確切作用。在過敏反應中,CD63可在特異性免疫球蛋白IgE介導的肥大細胞脫粒中起重要作用。Kraft等[28]研究表明,可將CD63作為過敏反應的重要組分,發(fā)現(xiàn)敲除CD63基因可顯著降低肥大細胞脫粒,從而減少體內(nèi)的急性過敏反應。本研究中,CD63基因在馬身豬的脂肪、肝臟、肌肉和脾臟的表達量較高,在心臟和腎臟中表達量較低,該結果與鱸魚和斑點叉尾鮰的表達結果類似[16,29],均在肝臟和肌肉中表達較高。脂肪組織除了可以儲存脂肪外,還具有免疫調(diào)節(jié)功能。作為免疫器官[30],脂肪組織中的T細胞是免疫代謝的關鍵因素[31],CD63基因在脂肪組織中高表達,是因為脂肪組織中已鑒定出多種免疫細胞,包括B細胞,T細胞,巨噬細胞和嗜中性粒細胞,而CD63在這些細胞表面均表達。馬身豬的肌內(nèi)脂肪含量較高[32],導致CD63在肌肉中的表達量也較高。此外肝臟可以選擇性富集自然殺傷細胞和殺傷性T細胞[33],因此推測CD63基因在免疫器官脂肪組織、脾臟和肝臟中高表達,可能參與相關免疫調(diào)控,其作用機制有待進一步研究。
4 結 論
本研究成功獲得了馬身豬CD63基因的CDS區(qū)序列(MN082631),全長714 bp,編碼237個氨基酸。生物信息學分析表明,CD63與駱駝科的羊駝和野駱駝同源性較高,為中性、親水、穩(wěn)定型四跨膜蛋白,二、三級結構預測顯示,該蛋白主要以α-螺旋和無規(guī)卷曲為主。該基因在所有組織中均有表達,在脂肪組織中的表達量最高,其次是肝臟、肌肉和脾臟。本研究結果為深入探討CD63基因的結構和功能提供了參考。
