甘肅省鼠疫耶爾森菌多位點可變數目串聯重復序列基因分型及地區分布
2021-08-31 08:24:00郭麗民席進孝葛亞俊王宇萌苗克軍徐大琴
中國人獸共患病學報 2021年8期
郭麗民,席進孝,葛亞俊,王宇萌,苗克軍,吳 斌,徐大琴
鼠疫是一種流行在嚙齒動物間通過蚤傳播的自然疫源性疾病,病原體為鼠疫耶爾森菌。鼠疫菌對糖、醇的酵解能力是生化分型的重要指標之一。紀樹立等[1]根據鼠疫菌生物學特征,將我國不同類型鼠疫疫源地分離的鼠疫菌株分為17個生態型。段永明等[2]將甘肅省鼠疫疫源地分離的菌株分為阿爾金型、青藏型、祁連型和甘寧黃鼠型。不同生態型的鼠疫菌均占有一塊各自相對獨立的疫源地分布區,而且菌株在分布區內占有優勢。各疫源地動物間鼠疫流行強度的差異,使得菌株在適應環境的變化過程中遺傳特征發生變化,這種變異被保存下來,造成不同疫源地間菌株在基因上的差異。由于甘肅省動物間鼠疫流行猛烈,常波及人間鼠疫疫情流行,而且非法獵捕販運旱獺容易造成鼠疫遠距離傳播,為此需要利用簡單經濟的分子分型方法對鼠疫疫情進行溯源分析。VNTR在鼠疫菌基因組中分布廣泛,可以通過分析多個位點的變異信息,進行鼠疫菌基因分型和進化分析,即MLVA(Multiple locus variable number tandem repeat analysis)方法。本實驗室早期利用15個VNTR位點進行鼠疫菌株的基因分型,只將鼠疫菌株分為兩個群,不能區分不同生化型的鼠疫菌株[3]。本次研究采用MLVA(14+12)分級分型的方法對甘肅省鼠疫菌株進行基因分型[4],以期建立甘肅省鼠疫菌VNTR位點多態性數據庫,為鼠疫疫情的鑒定溯源提供理論支持。
1 材料與方法
1.1 菌株來源及DNA制備 選取甘肅省1962-2014年間198株鼠疫菌,其中甘寧黃鼠型菌株6株、青藏型菌株128株、祁連型菌株46株、阿爾金型菌株18株。菌株保藏于甘肅省疾病預防控制中心鼠疫防制科。鼠疫菌DNA提取參照文獻[5]進行,置-20 ℃保存。
1.2 儀器及試劑 PCR基因擴增儀(美國Bio-Rad公司),凝膠成像分析系統(美國Bio-Rad公司),3730(美國ABI公司),5415D小型高速離心機(德國Eppendorf公司),水平電泳儀(北京六一儀器廠)。PCR擴增試劑購自北京全式金生物技術有限公司,DL1000(寶生物有限公司),瓊脂糖(西班牙Biowest公司),低分子量Markers和500liz、1200liz內標marker(北京賽百盛基因技術有限公司)。
1.3 方 法
1.3.1 引物 采用14+12對VNTR引物[4,6],引物信息見表1。

表1 14+12對MLVA引物序列信息
1.3.2 PCR反應體系及條件 反應體系:Premix Ex Taq 12.5 μL,10 μmol/L 上下游引物各1.0 μL,模板DNA 2 μL,補水至25 μL。PCR反應條件:95 ℃預變性5 min,95 ℃變性1 min,最適退火溫度1 min,72 ℃延伸1 min,30個循環,72 ℃ 延伸5 min。引物為M58、MS56、MS38、M21、M15、M61、M25、M23 時,退火溫度為60 ℃,引物為MS09、N2486、MS73、MS41、M34、M33、M22、M43、M28、M29時,退火溫度為55 ℃,引物為N3779、N2896、N0865、N2976、N1606、N2577、N3773、N2117體系的退火溫度為52 ℃ 1 min。
1.3.3 水平瓊脂糖凝膠電泳 取5 μL PCR產物,與1 μL 6×loading buffer混合后,采用1.5%瓊脂糖凝膠電泳后,凝膠成像分析有無產物和非特異性擴增、核對擴增產物大小。
1.3.4 PCR產物拷貝數的確定 將PCR產物送專業技術服務公司(北京睿博興科生物技術有限公司)進行毛細管電泳,根據分子量大小確定各個位點的拷貝數。
1.3.5 聚類分析 統計菌株各個位點的拷貝數,采用BioNumerics5.10軟件,采用效用均等的分類資料分析方法對各對引物的拷貝數進行聚類分析。為加強MLVA分型方法中14個低變異度指標在鼠疫種群中的區分能力,減弱高變異度的12個指標對聚類關系的影響,將第一級分型14個指標的權重設為2,用于種內溯源分析的12個指標權重設為1[7]。甘肅省1∶25萬矢量化鎮(鄉)地圖由中國疾病預防控制中心提供。采用ArcGIS10.3軟件進行鼠疫菌MLVA類群地區分布特征分析。
2 結 果
2.1 甘肅省鼠疫菌VNTR拷貝數結果 “14+12”個VNTR位點在甘肅省鼠疫菌基因組中拷貝數的結果見表2。與已完成全基因組測序的東方型鼠疫菌株CO92的VNTR位點拷貝數比較,26個VNTR位點均表現出不同于CO92的拷貝數。引物M34拷貝數變化類型最多,為17種,而最少的拷貝數變化類型為2種,說明26對引物的分辨能力高,能將甘肅省鼠疫菌株分為不同的基因型。
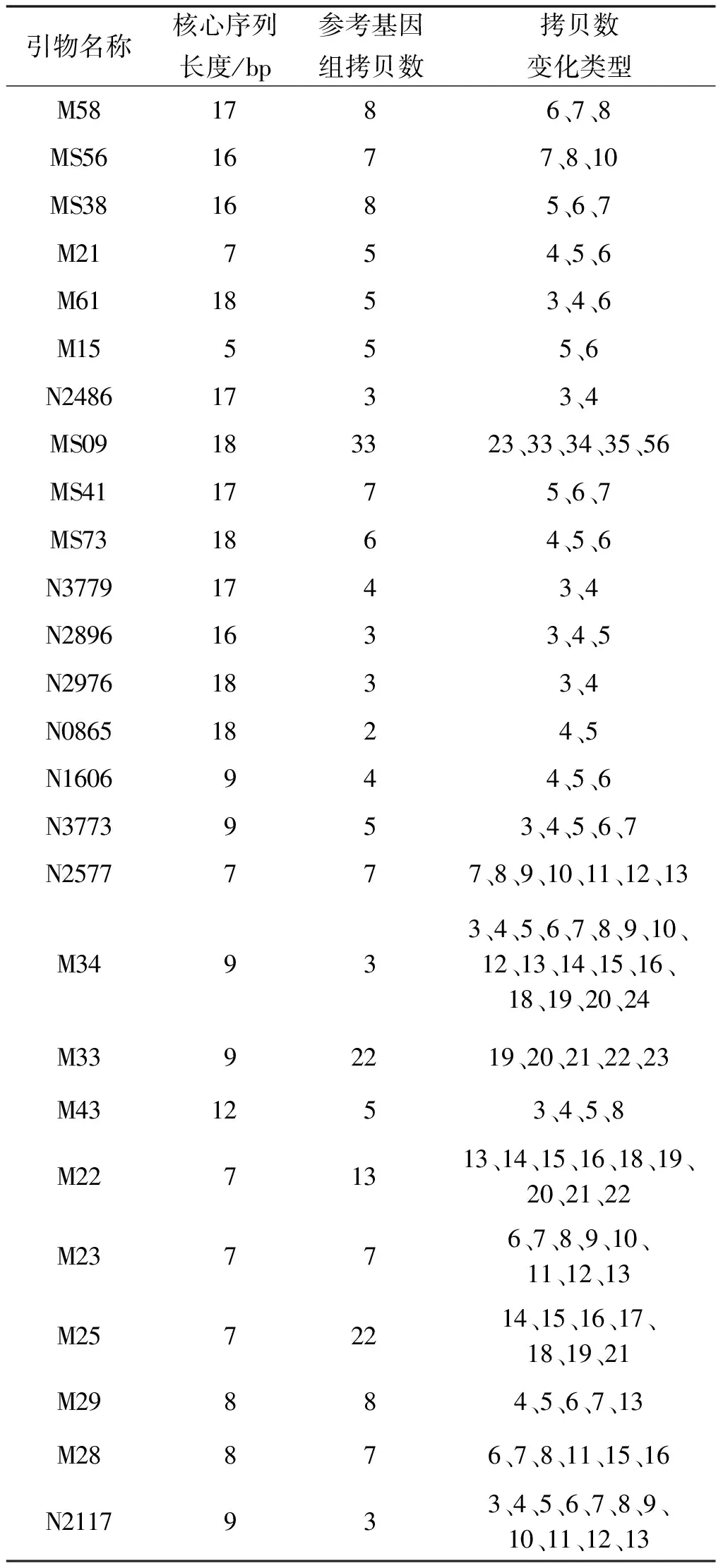
表2 14+12個VNTR位點在甘肅省鼠疫菌株基因組中的拷貝數變化類型
2.2 甘肅省鼠疫菌MLVA基因型及地區分布 “14+12”個VNTR位點將198株鼠疫菌分成1-127型,基因分型比較復雜。阿拉善黃鼠疫源地(會寧縣和平川區)MLVA基因型有122、124-127型;甘南高原疫源地(夏河縣)MLVA基因型有39-43型;阿爾金山鼠疫源地(阿克塞縣)MLVA基因型有89-115型;大雪山鼠疫疫源地(肅北縣和玉門市)有44-68型、69-88型、116-119型,祁連山北麓鼠疫源地(肅南縣)MLVA基因型有1-38型。見表3。
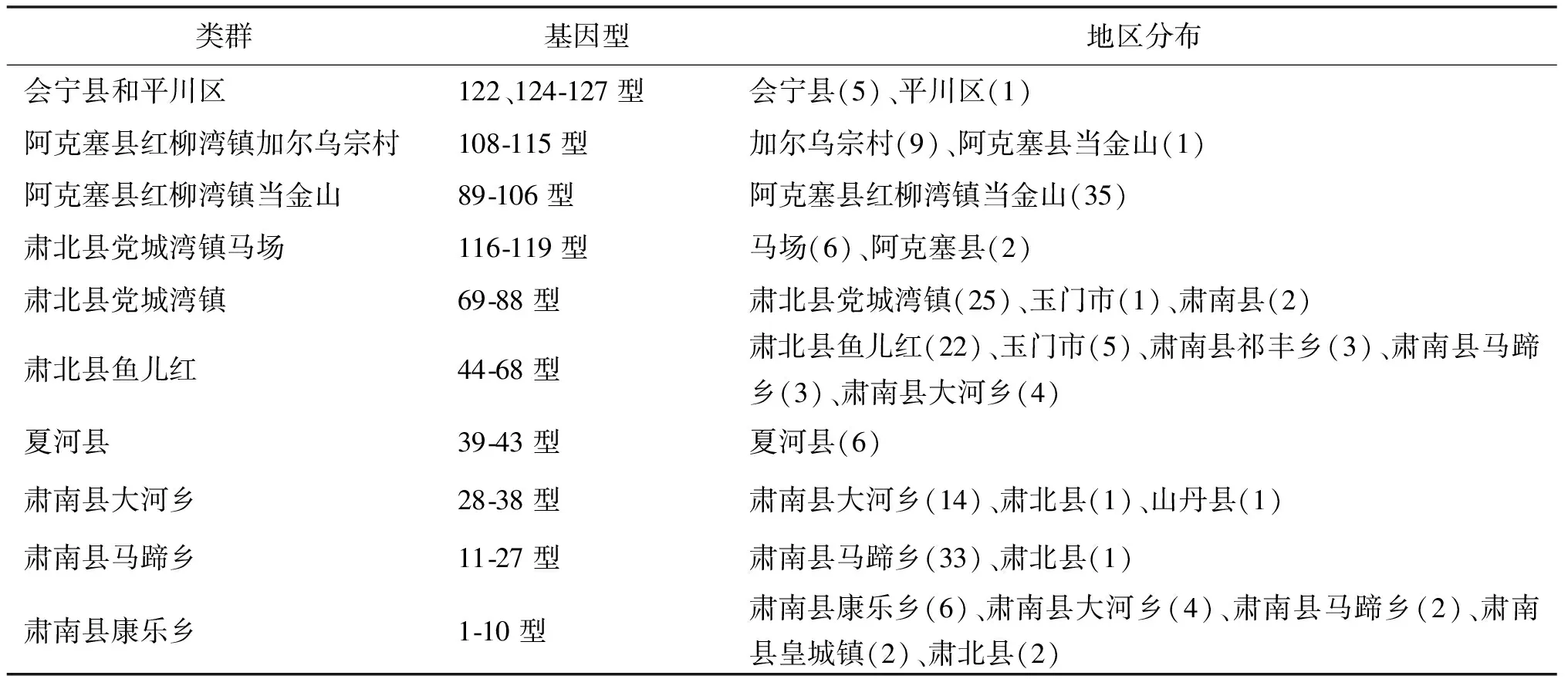
表3 甘肅省鼠疫菌MLVA類群及地區分布
2.3 甘肅省鼠疫菌VNTR聚類圖 采用“14+12”個VNTR位點分析中,將14個低變異度指標權重設為2,另外12個指標權重設為1。聚類結果顯示,198株鼠疫菌分為10個群,對比菌株的分離地點發現,大部分菌株根據分離地點聚集成為一群,每個群中菌株可繼續分為5-28個不同分支。通過聚類圖可以看出,阿拉善黃鼠疫源地菌株全部聚集為一群。阿克塞縣鼠疫菌株主要分為2個群,分別集中在阿克塞縣紅柳灣鎮當金山群(35/47)和加爾烏宗村群(10/47)。肅北縣鼠疫菌主要分為3個群,分別為黨城灣鎮馬場群(6/57)、黨城灣鎮群(25/58)和魚兒紅群(21/57)。玉門市菌株(5/6)主要聚集在肅北縣魚兒紅群內。夏河縣菌株單獨成為一群。肅南縣菌株主要分為3個群,分別為大河鄉群(14/73)、馬蹄鄉群(33/73)和康樂鄉群(6/73)。見圖1和圖2。
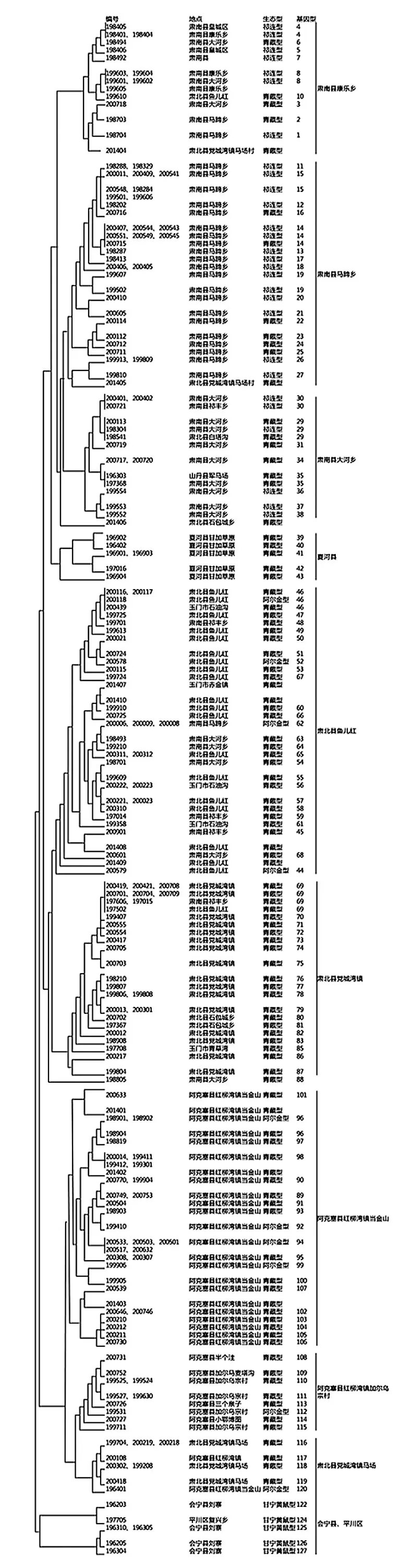
圖1 198株甘肅省鼠疫菌VNTR位點拷貝數聚類圖

注:甘肅省1∶25萬矢量化鎮(鄉)地圖由中國CDC提供。由于地圖中無法標注加爾烏宗村和馬場村,分別在相應的城鎮地圖中按照村的具體位置選取一定區域進行標注。
3 討 論
MLVA方法廣泛應用于鼠疫菌基因分型,但是VNTR位點選擇的不同對分子分型和聚類的結果存在一定差別。Pourcel等[8]人選用25個位點將180株鼠疫菌分成61個基因組型,而且3個生物型分別位于3個主要分支上,并發現中世紀型菌株存在一定的多態性。Klevytska等[9]采用46個位點對94株鼠疫菌進行分型,正確反映了古典型、中世紀型、東方型和田鼠型菌株之間的進化關系。Li等[4]從88個鼠疫菌VNTR位點篩選出“14+12”個位點用于快速溯源分析,提高了基因分型的分辨率,縮短實驗檢測的時間和成本。本實驗采用的26個位點能夠將生態型不同的鼠疫菌區分開,并呈現明顯的區域聚集性特征。
甘肅省鼠疫菌MLVA基因分型與生態型能夠很好的吻合,并且基因分型能夠繼續細分,精確到每個菌株的分離地點。會寧縣和平川區阿拉善黃鼠疫源地為甘寧黃土高原阿拉善黃鼠疫源地的西南部分,景觀為低山丘陵干草原,菌株為甘寧黃鼠型,菌株獨自成為一群。甘南高原疫源地的夏河縣和祁連山北麓東段區的肅南縣均為旱獺疫源地,景觀均為森林高山草甸草原。夏河縣菌株為青藏型,肅南縣菌株為祁連型。夏河縣菌株聚集成為一群后,與肅南縣菌株聚集在一起,肅南縣菌株根據分離地點繼續分為3個群,每群間菌株沒有明顯的地理屏障,菌株VNTR位點重復數有細微差別,每個群內菌株繼續分化,表明MLVA分型方法分辨率極高,可用于觀察菌株的微遺傳進化。阿爾金山疫源地的阿克塞縣和大雪山疫源地的肅北縣為旱獺疫源地,景觀類型為高山草原。阿克塞縣菌株主要為阿爾金型,肅北縣菌株主要為青藏型。阿克塞縣菌株分成2個群,是當金山群和祁連山西段分離的菌株聚集成的加爾烏宗村群,兩群菌株之間地理上接壤,沒有明顯的地理屏障。加爾烏宗村群與肅北縣黨城灣鎮馬場群的菌株聚集成為大群,兩地地理上接壤,沒有明顯的地理屏障,屬于祁連山西段,菌株之間交流頻繁,適應相應的地理環境成為優勢菌株。肅北縣的鼠疫菌除馬場群外,還存在黨城灣鎮群和魚兒紅群。黨城灣鎮群和魚兒紅群菌株VNTR位點不同,黨城灣鎮和魚兒紅之間存在大雪山地理屏障,使兩地菌株成為兩個群。玉門市鼠疫疫源地及毗鄰的肅南縣祁豐鄉生境特征與肅北縣魚兒紅相同,且地理位置接壤,屬于同塊疫源地,故菌株都聚集為一群。
甘肅省鼠疫菌遺傳特征復雜,采用MLVA分型方法發現肅南縣康樂鄉、馬蹄鄉和大河鄉群內均含有肅北縣菌株,而肅北縣黨城灣鎮、魚兒紅內含有肅南縣菌株,肅北縣黨城灣鎮馬場群內含有阿克塞縣菌株。這種群內菌株交叉存在的結果與DFR、CRISPR分型方法結果一致[10-12]。不同群內菌株的交叉存在,表明阿爾金山-祁連山北麓東段疫源地鼠疫菌基因組之間存在交流,究其原因有待進一步研究。鼠疫菌VNTR位點拷貝數不斷變化,菌株在自然選擇壓力作用下適應不同生態景觀成為某地區的主要基因類群,從而肅北縣和肅南縣疫源地菌株存在主要基因類群和次要基因類群。不同基因類群鼠疫菌的存在表明鼠疫在動物間持續流行。相對應處于肅北縣與阿克塞縣接壤地區的馬場,肅北縣和肅南縣接壤地區的祁豐鄉出現主次基因類群和不同生態型菌株交叉共存,出現菌株移行重疊現象,更例證了研究中發現群內菌株交叉存在的現象。同種疫源地出現不同類型的鼠疫菌,使得菌株溯源復雜多樣,尤其是在人間鼠疫疫情追蹤溯源時要予以重視。
MLVA 作為一種分子分型方法,其具有高分辨率,可操作性強,費用較低,適合推廣應用。本次采用MLVA(14+12)方法對198株甘肅省鼠疫菌進行基因分型,獲得準確的鼠疫菌VNTR位點拷貝數本底資料,為今后鼠疫菌株的追蹤溯源提供技術資料,為研究甘肅省鼠疫疫源地鼠疫菌遺傳進化規律奠定基礎,也為今后開展鼠疫菌遺傳突變的監測提供依據。
利益沖突:無
