siRNA干擾馬賽病毒潛在翻譯起始因子ORF314顯著抑制病毒復制
2021-08-29 07:53:28王為運夏宇程鐘江
微生物與感染 2021年6期
王為運,夏宇程,鐘江
復旦大學生命科學學院微生物學與免疫學系,上海 200438
21世紀初,首個感染阿米巴原蟲的巨型病毒——擬菌病毒的發現[1]使人們重新思考病毒的起源和本質。這些巨型病毒基因組可超過1 Mbp,編碼近 1 000 種蛋白質,遠超普通病毒。它們還編碼一些其他病毒中罕見的功能基因,特別是與蛋白質合成相關的基因。例如,擬菌病毒科的Tupanvirus和Klosneuvirus的基因組均編碼全套20種氨酰tRNA合成酶和數十種翻譯因子[2]。
馬賽病毒是繼擬菌病毒之后被發現的第2類阿米巴巨型病毒。2009年,第一株馬賽病毒在法國馬賽市一處冷卻塔的水樣中通過阿米巴共培養被分離[3],之后人們又陸續在突尼斯、塞內加爾、澳大利亞、日本、馬來西亞、印度和巴西等地分離出相似病毒[4-11]。2018年,國內首次報道了馬賽病毒上海株(Mar-SH2016)[12]。對Mar-SH2016基因組進行注釋發現,它和其他馬賽病毒一樣,編碼4種潛在翻譯因子,分別是類翻譯延伸因子G結構蛋白(ORF110)、真核生物多肽鏈釋放因子eRF1(ORF139)、翻譯延伸因子EF1a(ORF173)以及真核翻譯起始因子eIF2/eIF5(ORF314),但這些因子在馬賽病毒復制過程中的功能尚未得到深入研究。
真核生物蛋白質翻譯過程是一個受細胞精密調控的過程,包括起始、延伸和終止3個階段,其中起始階段至關重要,需要細胞內一系列物質的參與。真核翻譯起始因子通過與核糖體、mRNA以及起始tRNA之間的相互作用控制蛋白翻譯的起始,目前已發現了至少12種。Mar-SH2016編碼的ORF314基因全長453 bp,編碼151個氨基酸(amino acid,aa),蛋白相對分子質量為17.3×103,與宿主卡氏棘阿米巴(Acanthamoebacastellanii)編碼的eIF2/eIF5氨基酸序列同源性達60%。
本研究以ORF314基因為研究對象,通過RNA干擾技術降低其在Mar-SH2016感染的卡氏棘阿米巴細胞中的mRNA表達水平,分析其對病毒復制的影響,以期為揭示這些病毒編碼的相關潛在翻譯因子的功能提供理論基礎。
1 材料和方法
1.1 材料
1.1.1 細胞及病毒卡氏棘阿米巴Neff株(ATCC 30010)購自美國菌種保藏中心,用PYG(peptone-yeast extract with glucose)培養基于 25 ℃ 靜置培養[13]。馬賽病毒上海株Mar-SH2016(GenBank登錄號:MG827395)由本實驗室分離保存。
1.1.2 引物及抗體根據Mar-SH2016編碼的eIF2/eIF5(ORF314)基因序列設計實時熒光定量聚合酶鏈反應(polymerase chain reaction,PCR)引物,由蘇州金唯智生物科技有限公司合成,上游引物為5’-CTTGGTACGAGCTCCGGTTT-3’,下游引物為5’-CCACAACCTCTGCAGGACAT-3’。抗ORF314抗體用本實驗室免疫小鼠(上海斯萊克實驗動物有限責任公司)制備,辣根過氧化物酶(horseradish peroxidase,HRP)標記兔抗小鼠IgG購自生工生物工程(上海)股份有限公司,基礎型化學發光檢測試劑購自上海雅酶生物醫藥科技有限公司。
1.2 方法
1.2.1 靶向ORF314基因的小干擾RNA(small interfering RNA,siRNA)制備根據RNA干擾設計原則,針對Mar-SH2016編碼的eIF2/eIF5(ORF314)基因設計3對siRNA,另設計1對無義siRNA為陰性對照(negative control,NC),序列如表1所示。針對靶基因的siRNA采用5’CY3修飾,由上海吉瑪制藥技術有限公司合成。
1.2.2 siRNA轉染將卡氏棘阿米巴于12孔細胞培養板培養至密度為105cell/mL,移除PYG培養基,用PAS鹽溶液(按ATCC Meidum 1323標準配方配制)完全浸沒細胞,輕輕漂洗2~3次后加入適量新鮮PYG培養基。參照Lipofectamin RNAiMAX(Invitrogen)說明書進行細胞轉染,siRNA終濃度為100 nmol/L。轉染后于25 ℃靜置培養3 h,滴加Mar-SH2016病毒懸液,感染復數(multiplicity of infection,MOI)約為10,1 h后用PAS鹽溶液輕輕漂洗細胞,除去未吸附的病毒,重新加入適量PYG培養基,于25 ℃靜置培養,定時收集細胞。
1.2.3 轉染效率熒光檢測將5’CY3修飾的siRNA轉染卡氏棘阿米巴細胞4 h后,用CKX41熒光倒置顯微鏡(Olympus)紅色激發光對細胞進行觀察,攝影記錄轉染細胞的比例。
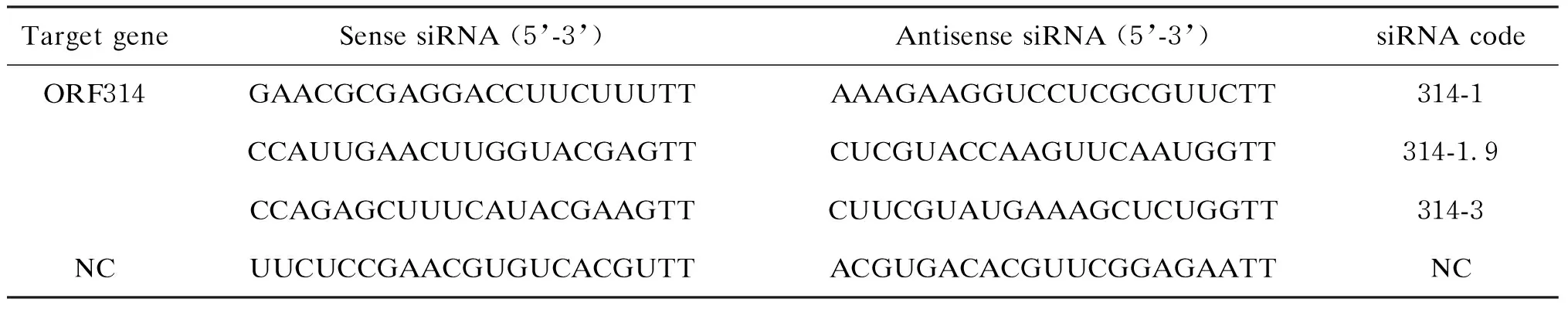
表1 靶向ORF314基因的siRNA序列
1.2.4 實時定量PCR(real-time quantitative PCR,RT-qPCR)檢測ORF314基因mRNA表達水平將卡氏棘阿米巴于12孔細胞培養板用PYG培養基培養(25 ℃)至細胞匯合度為70%~90%(密度約為105cell/mL),加入Mar-SH2016病毒懸液(MOI約為10),感染1 h后小心移去PYG培養基,用磷酸鹽緩沖液(phosphate buffered saline,PBS)輕輕漂洗3次,加入適量新鮮培養基,于25 ℃繼續培養。收集不同感染時間的細胞,用RNAiso Plus(TaKaRa)試劑盒抽提細胞總RNA,用PrimeScript RT reagent kit with gDNA eraser(TaKaRa)試劑盒對其進行反轉錄得到cDNA。以此cDNA為模板,以細胞線粒體細胞色素B基因為內參,用BlasTaqTM2X qPCR MasterMix(ABM)試劑盒進行RT-qPCR檢測。
1.2.5 ORF314基因干擾效果鑒定于4 ℃ 500 g離心5 min,收集siRNA干擾后的卡氏棘阿米巴細胞沉淀,用RNAiso Plus試劑盒抽提細胞總RNA,利用RT-qPCR檢測干擾后的ORF314基因mRNA表達水平。用棘阿米巴細胞裂解液[20 mmol/L Tris-HCl,pH 7.6;300 mmol/L NaCl;20 mmol/L KCl;6 mmol/L MgCl2;1% Triton X-100;1 mmol/L苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)]裂解細胞沉淀,提取蛋白,用十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)和蛋白免疫印跡法檢測干擾后的ORF314基因蛋白表達水平。
1.2.6 病毒毒力測定Mar-SH2016感染后6 h收集siRNA干擾后的細胞懸液,反復凍融3次,以釋放病毒顆粒,離心(500 g,5 min)收集含有病毒的上清液。向96孔板中每孔加入100 μL細胞懸液,密度約為4×104cell/mL,于25 ℃細胞培養箱中培養30 min至細胞貼壁,將含有病毒的上清液用PBS進行10倍系列稀釋至10-11。吸取100 μL病毒稀釋液,緩慢滴加至含有細胞的孔板中,于25 ℃細胞培養箱內培養5 d,顯微鏡下觀察,以出現明顯的滋養體細胞裂解死亡為陽性,用Reed-Muench法計算病毒毒力(TCID50)[14]。
1.3 統計學分析
2 結果
2.1 ORF314基因在Mar-SH2016感染細胞中的表達動態
Mar-SH2016感染卡氏棘阿米巴(MOI約為10)后一定時間收取阿米巴細胞,采用RT-qPCR分析ORF314基因的轉錄動態,以細胞線粒體細胞色素B基因為內參。結果顯示,ORF314基因在Mar-SH2016感染的宿主細胞中得到表達(見圖1)。感染后1.5 hORF314基因已有所表達,隨后表達水平持續升高,感染后8 h達到峰值,且一直持續至感染后16 h。
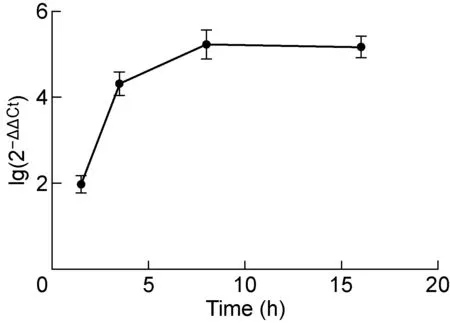
圖1 Mar-SH2016潛在翻譯因子ORF314的轉錄動態
2.2 siRNA轉染卡氏棘阿米巴的效果
用5’CY3修飾的siRNA轉染卡氏棘阿米巴細胞,4 h后在熒光顯微鏡紅色激發光下可以觀察到細胞質內分布著一些紅色熒光顆粒,細胞外也有一些游離的帶有熒光的siRNA復合物(見圖2)。結果顯示,1/3以上的細胞成功轉染siRNA。
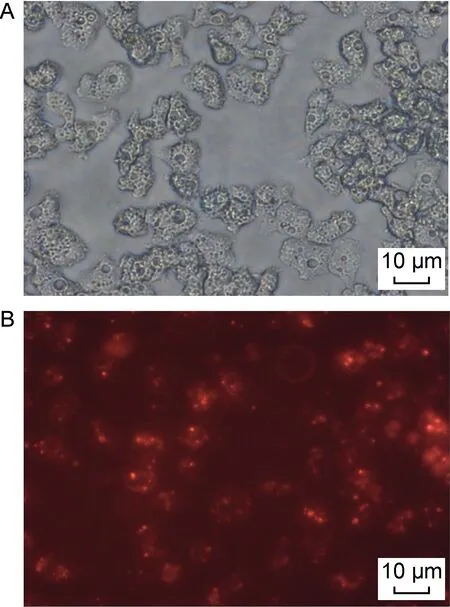
A: Bright field microscopy. B: Florescent microscopy with red excitation light.
2.3 siRNA干擾后ORF314基因表達下調
根據ORF314基因序列設計3對siRNA和1對無義siRNA,轉染卡氏棘阿米巴3 h后用Mar-SH2016感染(MOI約為10)。采用RT-qPCR和蛋白免疫印跡法,測定ORF314基因表達水平,結果如圖3所示。與轉染無關siRNA的陰性對照相比,靶向ORF314基因的3對siRNA中有2對能顯著降低其mRNA表達水平(P<0.01),編號分別是314-1和314-1.9(見圖3A)。蛋白免疫印跡結果顯示,與陰性對照相比,同樣是編號為314-1和314-1.9的2對siRNA干擾后的ORF314基因蛋白表達量降低(見圖3B)。
2.4 siRNA干擾ORF314基因表達后Mar-SH2016毒力降低
將靶向ORF314基因的3對siRNA轉染卡氏棘阿米巴,3 h后用Mar-SH2016感染(MOI約為10)。感染后6 h以卡氏棘阿米巴作為宿主細胞測定病毒毒力,結果如圖4所示。與轉染無關siRNA的陰性對照相比, 3對siRNA中有2對能顯著降低Mar-SH2016的毒力(P<0.01),編號分別是314-1和314-1.9,與干擾ORF314基因轉錄和蛋白合成的結果一致。Mar-SH2016毒力下降約30%,表明ORF314基因表達被干擾后Mar-SH2016的正常復制受到一定影響。

A: Transcriptional level of ORF314 in Mar-SH2016-infected cells after siRNA interference by RT-qPCR (**P<0.01 compared with NC). B: Protein expression of ORF314 in Mar-SH2016-infected cells after siRNA interference by Western blotting. ORF314: the purified ORF314 protein expressed in prokaryotic cells; A.c.-ORF314: cell lysates of Acanthamoeba castellanii after virus infection; Si-314-1, Si-314-1.9, Si-314-3 and Si-NC: cell lysates of Acanthamoeba castellanii transfected with three pairs of siRNA against ORF314 and control siRNA, respectively.
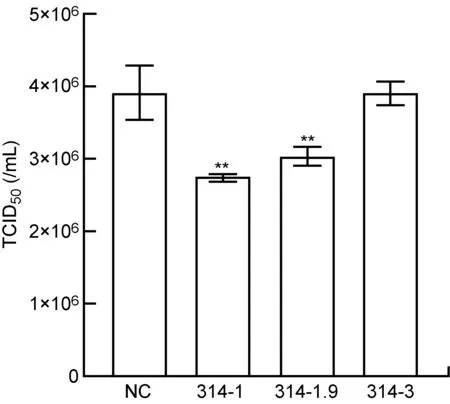
NC: cells treated with control siRNA; 314-1, 314-1.9, 314-3: cells transfected with three pairs of siRNA against ORF314. **P<0.01 compared with NC.
3 討論
大部分已知的巨型病毒基因組中超過一半的假定基因尚沒有明確的功能注釋[15],只有小部分擬菌病毒蛋白的功能得到了研究,包括一些氨基酰-tRNA合成酶[16-17]和參與糖生物合成的蛋白[18]。Sobhy等[19]首次利用RNA干擾技術降低擬菌病毒相關基因的表達,發現擬菌病毒纖毛的形成可能與功能未知的L725蛋白和L829蛋白、注釋功能為氧化還原酶的R135蛋白以及屬于TPR蛋白超家族[20-22](介導蛋白質與蛋白質、病毒與宿主之間相互作用以及調節病毒復制的一類蛋白質)的R856蛋白有關,敲低這4個基因對擬菌病毒的纖毛形成有影響。Zinoviev等[23]對擬菌病毒科和馬賽病毒科病毒編碼的潛在翻譯延伸因子1家族的GTPase(trGTPase)進行研究發現,前者的trGTPase與GTP結合蛋白1(GTP binding protein 1,GTPBP1)的關系密切,在翻譯延伸過程中能將同源氨基酰-tRNA傳遞到核糖體A位點,而后者的trGTPase與真核生物翻譯延伸因子eEF1A、多肽鏈釋放因子eRF3和mRNA檢測因子Hbs1的關系更為密切,具有類似于eRF3的功能,可終止翻譯進程并促進多肽鏈的釋放。
本研究對馬賽病毒上海株Mar-SH2016的真核翻譯起始因子eIF2/eIF5(ORF314)進行RNA干擾,用RT-qPCR檢測其轉錄動態。結果顯示,在Mar-SH2016感染宿主卡氏棘阿米巴的過程中,ORF314基因呈現動態表達,于約1.5 h內開始表達,感染后8 h表達量達到峰值且持續至感染后16 h。將siRNA轉入宿主細胞,用Mar-SH2016感染6 h后ORF314基因表達水平顯著下降,且病毒毒力顯著降低(P<0.01),下降近30%,表明病毒的復制受到影響。將siRNA轉染卡氏棘阿米巴細胞并延長病毒感染時間至9 h,ORF314基因的表達不再被siRNA顯著影響(結果未顯示)。這可能是由于siRNA的干擾效果持續時間有限,須進一步加大siRNA用量并設法提高轉染效率。
此外,siRNA引起Mar-SH2016毒力降低的幅度遠低于ORF314基因轉錄水平下降的幅度。這可能是因為馬賽病毒ORF314基因與宿主棘阿米巴基因組中的氨基酸序列同源性達60%,可以在一定程度上彌補ORF314水平降低對病毒復制的影響。盡管如此,本研究發現siRNA干擾ORF314基因表達能顯著抑制Mar-SH2016的早期復制,提示Mar-SH2016在感染早期可能需要ORF314基因的協助來進行自身蛋白質合成,表明從嚴格意義上來說馬賽病毒編碼的這些與翻譯相關的基因并不是冗余的。
巨型病毒的發現改變了以往人們對病毒的認知,對馬賽病毒的一些重要問題開展深入研究有助于了解此類巨型病毒的復制機制,加深對其本質和起源的認識,同時有助于認清其對人類及其他生物的潛在影響,為進一步探究其在生態系統中的地位和作用提供了理論依據。
