儲存紅細胞中表達降低的circRNA以及其靶基因預測的研究*
2021-08-24 03:01:46張怡宇黃國清王強張勇剛苑召虎陳小潔黃建云李楠魏亞明
臨床輸血與檢驗 2021年4期
關鍵詞:研究
張怡宇 黃國清 王強 張勇剛 苑召虎 陳小潔 黃建云 李楠 魏亞明
在低溫儲存條件下,儲存紅細胞會發生一系列的變化,也稱為儲存損傷。氧化損傷和代謝障礙是儲存損傷的原因,而ATP減少、2,3-DPG變化、離子泵功能、RBC膜和細胞骨架的重組和破壞、血紅蛋白和氧化蛋白的結合、帶3蛋白的降解以及筏蛋白的變化是儲存損傷的具體表現[1]。在儲存過程中積累的部分生物反應調節劑(BRM)會充當輸血受血者的促炎劑,例如細胞因子、趨化因子、生物活性脂質和代謝產物[2]。如果將這些發生變化的血制品輸入到患者,有可能引起輸血不良反應。在低溫保存條件下,供體紅細胞的表現存在廣泛的遺傳變異性,所以在組學基礎上制定新途徑來減少儲存損傷也十分不易[3]。
在全血中,環狀RNA(circular RNA,circRNA)是轉錄組的天然成分[4]。circRNA呈閉合環形結構,在無核細胞中大量存在。它不易被核糖核酸酶R水解,而且比線性RNA更穩定,所以更適宜用作生物標記物[5,6]。除此外,circRNA是高效的miRNA海綿,通過不同的結合位點能吸附多個miRNA,通過解除miRNA對mRNA的抑制作用,來促進mRNA表達及蛋白翻譯,參與調節多種信號通路[7,8]。circRNA還可以起到存儲,分類或定位RBP(RNA結合蛋白)的作用,如Foxo3環狀RNA與特定蛋白質之間的相互作用顯示出可延緩細胞周期進程[4]。
在多數疾病中,circRNA上調和下調與具體的疾病機制密切相關。在無核細胞中,整個轉錄組呈降解趨勢。已有研究證明在儲存紅細胞中非編碼RNA之一的miRNA下調模式更為明顯[9],而本研究中下調的circRNA數量遠大于上調的circRNA數量,研究下調的circRNA更具代表意義。下調的circRNA其作為儲存損傷標志物的候選者更為合理。基于這個背景,本研究對儲存20 d的紅細胞和新鮮紅細胞采用高通量測序,觀察下降明顯的circRNA的表達變化及circRNA所對應的miRNA的分子功能,用生物信息學方法分析下降明顯的circRNA、其靶miRNA及miRNA關聯的mRNA并進行PCR驗證,預測其功能以及作為儲存損傷標志物的可能性。
材料和方法
1 一般資料 血液標本為5名25~30歲O型健康合格獻血者的標本200 mL(人)份,與CPDA-1保存液混合,過濾去除白細胞后制成懸浮紅細胞,分別收集新鮮組(0 d)、20 d收集標本進行試驗。收集方法如下:以(1 000 g,20 min,4℃)為條件進行輕離心,在生物安全柜內將血漿層(去除血小板和血漿)吸走,留取紅細胞層,用PBS洗滌三次。其中3名健康者標本用來circRNA測序,所有5名健康者標本用來circRNA的RT-qPCR驗證。所有試驗均分為0和20 d 2組。研究獲本院醫學倫理委員會批準(K-2018-027-01)。
2 試劑與儀器 紅細胞保存液CPDA-1購自廣州費森尤斯卡比;0.9%生理鹽水購自佛山雙鶴藥業;PrimeScript RT Master reagent Kit with gDNA Eraser(perfect Real Time)、SYBR Premix Ex Taq Master MixⅡ、無酶水均購自日本TaKaRa公司;TRIzol-LS購自美國life公司;RNaseR購自美國Epicentre公司;一次性白細胞過濾輸血器購自南京雙威公司;自動紅細胞計數儀購自北京賽科希德公司;電熱恒溫培養箱購自上海精宏公司;低溫高速離心機購自美國Eppendorf公司;引物序列由上海捷瑞合成;使用qTOWER2.2購自德國analytikjena的PCR儀做qRTPCR檢測。
3 circRNA高通量測序檢測 3名健康者標本的新鮮紅細胞和4℃儲存20 d紅細胞分別提取總RNA儲存于-80℃,進行circRNA高通量測序。對0 d組、20 d組的6份標本(3份/組)做高通量測序。
4 circRNA的生物信息學方法與流程 對下調明顯的十個circRNA的類型、來源、新舊進行分析,最終完成其功能注釋。用circBase(http://circbase.org/)與mirTarBase(http://mirtarbase.mbc.nctu.edu.tw/php/index.php)完成數據庫注釋與靶向預測。然后對cirRNA作用miRNA進行靶標預測,所用軟件是mireap(http://mireap.sourceforge.net/),miranda(https://sourceforge.net/projects/mireap/),targetscan(http://www.targetscan.org/vert_72/)。并將miRNA可能作用的mRNA也進行了關聯分析,所用軟件是mirTarBase(mirtarbase.mbc.nctu.edu.tw/php/index.php)。
5 circRNA、miRNA、mRNA表達量的測定 采用qRT-PCR方法測定miRNA的相對表達量,具體方法參考文獻[10]。circRNA的引物如表1所示,miRNA及mRNA引物設計用軟件prime3(http://frodo.wi.mit.edu/primer3/)完成。

表1 qRT-PCR用到的引物
6 統計學處理 應用SPSS 16.0軟件和Graphpad Prism5軟件行配對t檢驗,數據以均數±標準差()表示,P<0.05表示差異有統計學意義。
結 果
1 測序結果分析 紅細胞在儲存20 d對比0 d中,下調明顯的10個circRNA有:hsa_circ_0005413、novel_circ_0002503、hsa_circ_0008937、hsa_circ_0040340、hsa_circ_0003126、hsa_circ_0007995、hsa_circ_0023937、novel_circ_0000402、hsa_circ_0049462、hsa_circ_0028281(圖1)。下調明顯的前10個circRNA有3條來自11號染色體,且這10個circRNA均來自外顯子(表2)。Novel命名的circRNA表示在紅細胞中發現的新circRNA,即目前未被circBase數據庫收錄的circRNA。
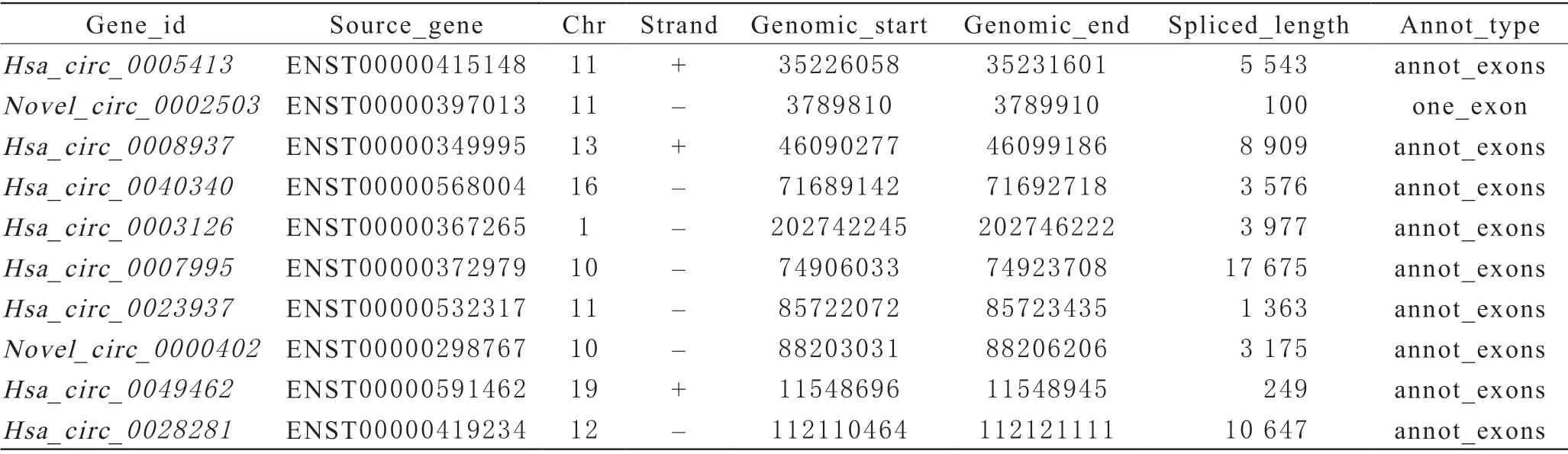
表2 circular RNA類型統計表
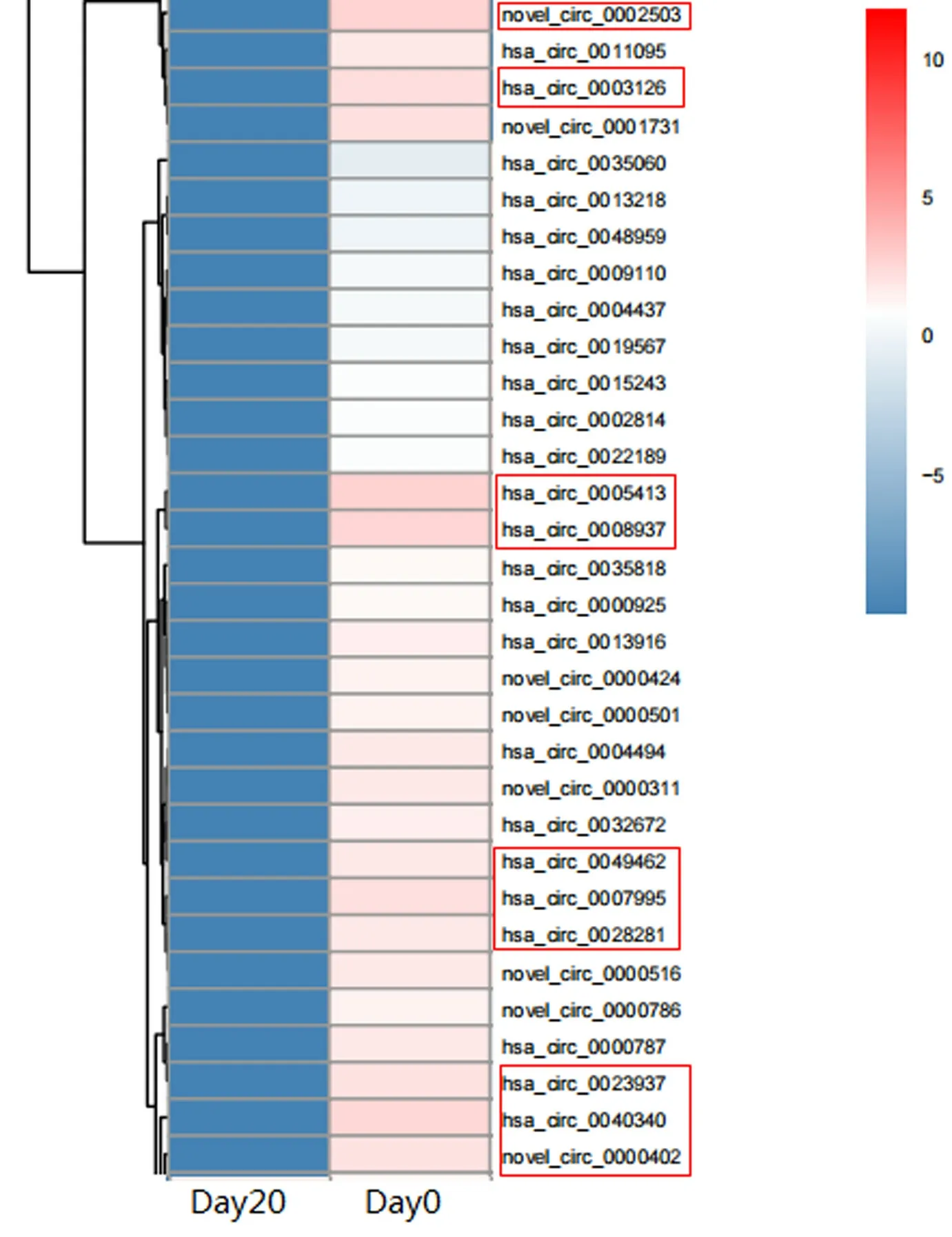
圖1 circRNA表達情況熱圖(n=3)
2 生物信息學預測結果 利用生物信息學軟件預測發現下降明顯的十個circRNA中,hsa_circ_0005413能海綿吸附hsa-miR-6748-3p,hsa-miR-6871-5p,hsamiR-7160-5p,其中hsa-miR-6871-5p能靶向作用于BACH1(basic leucine zipper transcription factor 1,
堿性亮氨酸拉鏈轉錄因子1)、SOD2(superoxide dismutase 2,超氧化物歧化酶2)等mRNA;hsamiR-7160-5p能靶向作用于EIF2AK2(eukaryotic translation initiation factor 2-alpha kinase 2,真核翻譯起始因子2-α激酶2)、CASP8(caspase 8,半胱天冬酶8)等mRNA。hsa_circ_0008937能海綿吸附hsa-miR-6778-3p,hsa-miR-6778-3p能靶向作用于SH3BP5(SH3-domain binding protein 5,SH3-結構域結合蛋白5)、AKAP2(A kinase(PRKA)anchor protein 2,激酶(PRKA)錨蛋白2)、PPP1R12C(protein phosphatase 1,蛋白磷酸酶1)等mRNA。hsa_circ_0040340能海綿吸附hsa-miR-122-5p、hsamiR-3173-5p、hsa-miR-5590-5p,其中hsa-miR-122-5p能靶向作用于MAPK1(mitogen-activated protein kinase 1,絲裂原激活的蛋白激酶1)、BCL2L1(BCL2-like 12(proline rich),BCL2樣12(富含脯氨酸))、CDK4(cyclin-dependent kinase 4,細胞周期蛋白依賴性激酶4)等mRNA;Hsa-miR-3173-5p能靶向作用于CPEB4(cytoplasmic polyadenylation element binding protein 4,胞質聚腺苷酸化元件結合蛋白4)、SCAMP2(secretory carrier membrane protein 2,分泌載體膜蛋白2)、CAMKV(CaM kinase-like vesicle-associated,CaM激酶樣囊泡相關)、ARHGEF2(Rho guanine nucleotide exchange factor(GEF)2,Rho鳥嘌呤核苷酸交換因子(GEF)2)。hsa_circ_0007995能海綿吸附hsa-miR-1254、hsa-miR-4534、hsa-miR-6751-5p、hsa-miR-99a-3p,其中Hsa-miR-4534能靶向作用于TMEM214(transmembrane protein 214,跨膜蛋白214)、EMC3(ER membrane protein complex subunit 3,ER膜蛋白復合物亞基3)、THAP1(THAP domain containing 1,THAP域包含1)、PER2(period circadian clock 2,周期生物鐘2)、SEMA7A(semaphorin 7A,GPI膜錨)、CAPZA1(capping protein(actin filament)muscle Z-line,alpha 1,旋蓋蛋白(肌動蛋白絲)肌肉Z線,alpha 1)、MAP1LC3B(microtubuleassociated protein 1 light chain 3 beta,微管相關蛋白1輕鏈3 beta)、ABCA6(ATP-binding cassette 6,ATP結合盒成員6)、PGM2L1(phosphoglucomutase 2-like 1,磷酸葡萄糖突變酶2樣1)、TMOD3(tropomodulin 3,原肌鈣蛋白3)、PPP2R1B(protein phosphatase 2,regulatory subunit A,beta;蛋白磷酸酶2,調節亞基A,β)、TMEM214(transmembrane protein 214,跨膜蛋白214)。novel_circ_0000402能海綿吸附hsa-miR-1273g-3p、hsa-miR-1285-3p、hsamiR-4304,其中hsa-miR-1285-3p能靶向作用于PRSS21(proteaseserine21,蛋白酶絲氨酸21)、MRI1(methylthioribose-1-phosphate isomerase 1,甲基硫代核糖-1-磷酸異構酶1)、UFM1(ubiquitin-fold modifier 1,泛素倍數調節劑1)。
3 circRNA的表達量 熒光定量PCR顯示相對于新鮮紅細胞組,儲存20 d組的hsa_circ_0005413、hsa_circ_0008937、hsa_circ_0040340、hsa_circ_0003126、hsa_circ_0007995、hsa_circ_0023937、novel_circ_0000402、hsa_circ_0028281表達量顯著下降(P<0.05,表3)。其中novel_circ_0002503和hsa_circ_0049462未被RTPCR所驗證,可能因為其片段比較短。
表3 circRNA的qRT-PCR表達結果(,n=5)
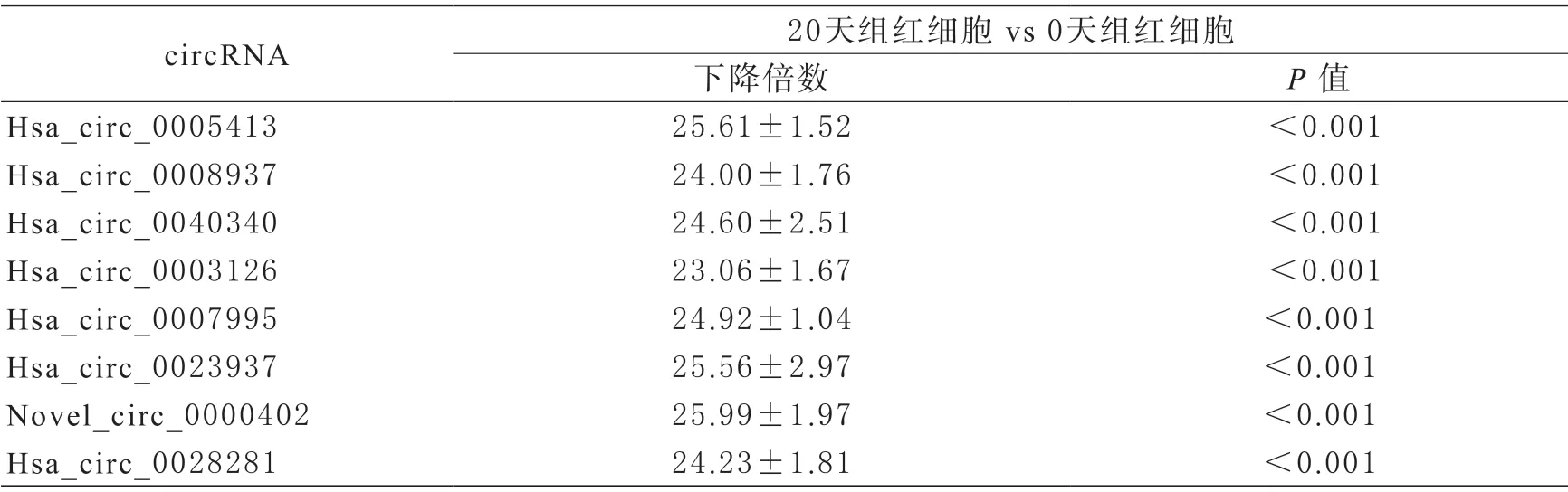
表3 circRNA的qRT-PCR表達結果(,n=5)
4 miRNA的表達量 由于circRNA能海綿吸附miRNA,而miRNA能靶向作用于mRNA的性質,許多研究證實了競爭性內源RNA-(competing endogenous RNAs,ceRNA)機制的存在。根據ceRNA機制原理,circRNA和mRNA的表達量變化應是同一方向,而circRNA和miRNA的表達量變化應是相反方向。之前的芯片研究揭示了紅細胞儲存中miRNA的表達量變化[9]。本研究在生物信息學預測到的miRNA中,選取了在芯片中表達量升高的miRNA做qRT-PCR驗證。熒光定量PCR顯示相對于新鮮紅細胞組,儲存20天組的hsa-miR-7160-5p、hsamiR-122-5p、hsa-miR-4534的表達量顯著上升(P<0.05,表4)。
表4 miRNA的qRT-PCR表達結果(,n=5)
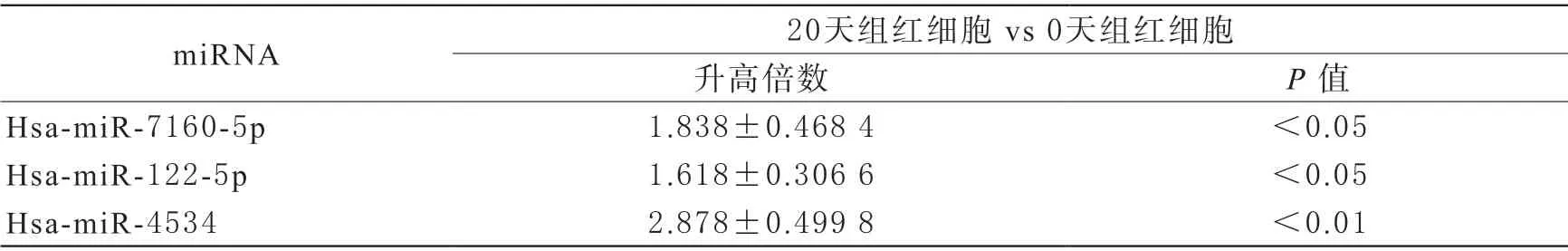
表4 miRNA的qRT-PCR表達結果(,n=5)
5 mRNA的表達量 在生物信息學預測結果基礎上,對預測得到的靶mRNA查閱大量文獻。篩選出在儲存損傷中發揮重大作用的3條mRNA進行RT-qPCR驗證。熒光定量PCR顯示相對于新鮮紅細胞組,儲存20 d組的CASP8、CDK4、TMOD3顯著下降(P<0.05,表5)。
表5 mRNA的qRT-PCR表達結果(,n=5)
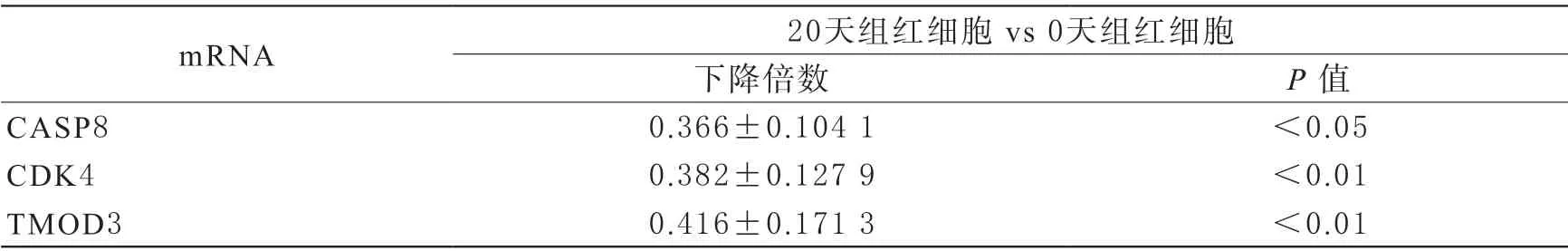
表5 mRNA的qRT-PCR表達結果(,n=5)
6 與儲存損傷有關的circRNA-miRNA-mRNA通路分析 經過qRT-PCR驗證及文獻查閱,初步發現hsa_circ_0005413-hsa-miR-7160-5p-CASP8、hsa_circ_0040340-hsa-miR-122-5p-CDK4、hsa_circ_0007995-hsa-miR-4534-TMOD3軸,參與了儲存損傷的生物學過程。
討 論
本研究從circRNA-miRNA-mRNA網絡角度為紅細胞儲存損傷機理和預防提供了新見解。hsa_circ_0005413-hsa-miR-7160-5p-CASP8、hsa_circ_0040340-hsa-miR-122-5p-CDK4、hsa_circ_0007995-hsa-miR-4534-TMOD3軸是本文全新發現。這3條軸的miRNA分子中,暫未有hsa-miR-7160-5p的研究,miR-122在肝脂肪變性中上調,與衰老相關,并在多種人體組織中過表達[11-13],hsa-miR-4534在健康人和2型糖尿病血清樣本中的表達有顯著差異[14]。這3條軸中的mRNA分子中,caspase-3和-8在成熟紅細胞大量存在,由于存在這些蛋白酶,缺少線粒體凋亡級聯反應的必需成分如caspase-9、Apaf-1和細胞色素c的紅細胞可以進入凋亡途徑[15]。細胞周期蛋白依賴性激酶4(cyclindependent kinase 4,CDK4)在人紅細胞中存在,CDK4抑制劑能減弱氧化應激下的PS暴露,降低紅細胞凋亡率[16]。(Tropomodulin3,TMOD3)是一種結合并覆蓋在紅系和非紅系細胞的肌動蛋白絲尖端的蛋白質,TMOD3的破壞會使紅細胞生成受損,它介導的肌動蛋白重塑對于成紅細胞-巨噬細胞粘附,協調細胞周期與分化以及F-肌動蛋白組裝和成紅細胞去核過程中的重塑中起著重要作用[17]。另外TMOD3對肌動蛋白絲的覆蓋不足可能會導致尖端的肌動蛋白動力學更大,絲長重新分布導致血影蛋白-肌動蛋白網狀結構連接不規則和減弱,導致紅細胞膜不穩定[18]。可以初步推測,hsa_circ_0007995可通過吸附hsa-miR-4534下調TMOD3的表達導致紅細胞膜不穩定性增強。hsa_circ_0005413吸附hsa-miR-7160-5p,下調CASP8的表達阻止紅細胞凋亡通路,hsa_circ_0040340能吸附hsa-miR-122-5p下調CDK4的表達,降低紅細胞凋亡率。hsa_circ_0005413和hsa_circ_0040340可能是潛在的延緩儲存損傷的有益基因。
根據本研究的生物信息學預測結果,未經過RT-qPCR驗證的hsa_circ_0005413-hsa-miR-6871-5p-BACH1、hsa_circ_0005413-hsa-miR-6871-5p-SOD2、hsa_circ_0040340-hsa-miR-122-5p-MAPK1、hsa_circ_0040340-hsa-miR-122-5p-BCL2L1、hsa_circ_0007995—Hsa-miR-4534-PER2、hsa_circ_0007995—Hsa-miR-4534-SEMA7A途徑有可能在紅細胞生理過程發揮了重要作用。例如,轉錄因子Bach1可能參與成紅細胞對缺鐵狀態的反應,它通過與血紅素的直接結合而失活[19]。血紅素通過轉錄因子Bach1在紅系細胞中正調控基因座控制區域的β-珠蛋白表達[20]。超氧化物歧化酶2(super oxide dismutases,SOD2)缺乏將導致小鼠紅細胞變形能力降低和血紅素降解增加[21]。紅細胞祖細胞中SOD2的丟失會導致蛋白質氧化損傷增強,膜變形改變以及紅細胞存活率降低[22]。絲裂原相關蛋白激酶1(mitogen-associated protein kinase 1,MAPK1),成紅細胞巨噬細胞蛋白(Emp)在紅細胞和巨噬細胞中均表達,Emp的下調會影響絲裂原相關蛋白激酶1(MAPK1)和胸腺瘤病毒原癌基因(AKT-1)的表達,從而導致細胞運動異常[23]。Bcl-2樣蛋白1(BCL2L1)屬于抗凋亡因子,有間接證據表明bcl-x可能在紅系細胞的凋亡中起作用[24]。有研究表明,將c-MYC和BCL-XL轉導至多能造血祖細胞中過表達,使糖蛋白A(+)成紅細胞能夠持續自我復制[25]。周期生物鐘基因Per2(Period circadian clock genes 2,Per2)具有非晝夜節律功能,Per2耗竭導致紅細胞代謝譜發生重大變化,包括乳酸增加和ATP水平降低,所以它的丟失會大大減少紅細胞的壽命[26]。JMH血型系統由6種位于Sema7A蛋白上的抗原組成,(Semaphorin 7A,Sema7A)是攜帶JMH血型抗原的蛋白質,參與免疫反應,在軸突生長和引導中起重要作用[27]。
除此以外,有些靶mRNA還能誘導circRNA環化,即促進circRNA的生成。如真核生物起始因子4A3(Eukaryotic initiation factor 4A3,EIF4A3)可誘導circMMP9環化,并增加了多形性膠質母細胞瘤細胞中circMMP9的表達[28]。這種調控機制是否在紅細胞儲存損傷中發揮作用還需要進一步研究。
在本文中,選取了高通量測序對circRNA分子進行研究。首先,選取下降明顯的十個circRNA做PCR驗證,其次用circBase和mirTarBase完成數據庫注釋與靶向預測,推測了circRNA海綿吸附miRNA的潛在功能。第三,將miRNA可能作用的mRNA也進行了關聯分析,構建了circRNA-miRNA-mRNA通路。本研究的一個局限是,并未對所有差異表達的circRNA進行PCR驗證和靶基因預測分析。但是我們的數據為紅細胞儲存損傷中的circRNA分子機制研究提供了新基礎,并發現某些circRNA可能與儲存損傷密切相關。利益沖突 所有作者均聲明不存在利益沖突
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
