5-羥色胺對豬植入前胚胎發(fā)育的作用
2021-08-10 07:27:10段佳慧張學明李子義
中國獸醫(yī)學報 2021年6期
關鍵詞:水平
韓 宇,張 蒙,段佳慧,張學明,李子義,唐 博*
(1.吉林大學 動物醫(yī)學學院,吉林 長春 130062;2.吉林大學 第一醫(yī)院 轉化醫(yī)學研究院 人類疾病動物模型國家地方聯(lián)合工程實驗室,吉林 長春 130062)
5-羥色胺(5-HT),又稱血清素,是一種廣泛分布于中樞神經(jīng)系統(tǒng)和胃腸道的神經(jīng)遞質,也是一種激素和生長因子,在動物發(fā)育、生理和病理過程中發(fā)揮著至關重要的作用[1]。5-HT廣泛分布在雌性生殖組織如人類卵巢、大鼠子宮、輸卵管、卵巢和胎盤以及小鼠卵母細胞和顆粒細胞[2]中,并在整個胚胎發(fā)育的早期階段起著至關重要的作用[3]。外源性添加5-HT,可抑制小鼠囊胚的形成,增加凋亡率[4];植入后胚胎中添加5-HT會影響胎兒發(fā)育,導致胚胎畸形如顱面缺陷和先天性心臟畸形[5]。
在哺乳動物胚胎發(fā)育過程中,父方和母方的基因組必須經(jīng)歷表觀遺傳重構和轉錄模式的改變才能實現(xiàn)全能性[6]。表觀遺傳修飾是調節(jié)細胞命運、分化和衰老等關鍵過程的決定性因素,主要包括DNA的共價修飾和組蛋白的翻譯后修飾[7]。組蛋白修飾是組蛋白在相關酶的激活下進行甲基化、乙酰化、磷酸化、腺苷化等修飾過程,是一種廣泛存在于真核生物中的表觀遺傳修飾模式,在調節(jié)基因活性和染色質狀態(tài)中發(fā)揮著重要作用[8]。不同類型的組蛋白修飾具有不同的分布和功能[9],例如,H3K4me3可以標記基因啟動子和轉錄的起始[10], H3K27me3可作為發(fā)育基因啟動子的抑制標記[11]。此外,H3K27Ac是一種啟動子和增強子的活性標記物[12]。因此,在哺乳動物胚胎發(fā)育過程中,組蛋白修飾可以調控基因表達,影響轉錄調控。
動物在生存過程中受到刺激后會產(chǎn)生應激反應,在應激狀態(tài)下,下丘腦-垂體-腎上腺軸(HPA軸)失調,進而使5-HT的分泌紊亂[13]。在母豬飼養(yǎng)過程中,存在很多應激刺激,繼而造成母體5-HT的分泌變化,這是否會對胎兒發(fā)育產(chǎn)生影響?豬胚胎中5-HT的變化是否會影響胎兒的表觀遺傳?為了探索這些未知問題,本研究用5-HT處理豬孤雌胚胎,探究了5-HT與胚胎發(fā)育及表觀遺傳修飾三者之間的關系。本研究對5-HT在豬胚胎上的作用機制提供了新的見解,并揭示了其在胚胎發(fā)育過程中發(fā)揮作用的潛在途徑。
1 材料與方法
1.1 試劑RNA提取試劑盒(RNeasy Mini Kit),購自Qiagen公司(德國);反轉錄試劑盒(TransScript All-in-One First-Strand cDNA Synthesis SuperMix),購自全式金公司(中國);細胞凋亡試劑盒(InSituCell Death Detection Kit)、熒光定量試劑盒,購自Roche公司(德國);H3K4me2,H3K4me3一抗,購自Abcam 公司(英國);5mC、5hmC一抗,購自Active Motif公司(美國);熒光二抗購自武漢博士德公司;其他所使用的化學試劑除特殊指出之外,均購自Sigma公司(美國)。卵母細胞成熟及胚胎培養(yǎng)所用培養(yǎng)液配方參照文獻[14]配制。
1.2 卵母細胞的收集和體外成熟(IVM)豬卵巢取自長春當?shù)赝涝讏觯糜?5.0~38.5℃的無菌生理鹽水中并在4 h內將其運送到實驗室。用10 mL注射器抽取3~6 mm直徑的卵泡,在體式顯微鏡下挑選包裹著均勻顆粒細胞的卵丘-卵母細胞復合物(COCs)。在添加3%牛血清白蛋白(BSA)的TCM-199中洗滌3遍后,將COCs轉移至體外成熟培養(yǎng)液中,在38.5℃、5% CO2及100%濕度的培養(yǎng)箱中培養(yǎng)42~44 h。
1.3 孤雌激活(PA)卵母細胞體外成熟后,利用透明質酸酶去除卵丘細胞,并挑選出卵周隙明顯、卵黃膜完整且排出第一極體的成熟卵母細胞。將卵母細胞置于融合液(甘露醇0.3 mmol/L,CaCl21.0 mmol/L,MgCl20.1 mmol/L,Hepes 0.5 mmol/L)平衡1 min,轉入鋪滿融合液的融合槽中,利用BTX2001細胞融合儀以1.2 kV/cm,30 μs,2次直流脈沖的條件進行激活。之后對照組轉入胚胎培養(yǎng)液PZM-3中進行培養(yǎng),試驗組則置于添加了不同濃度的5-HT胚胎培養(yǎng)液PZM-3中,于38.5℃、5% CO2和100%濕度的條件下進行培養(yǎng),48 h 后觀察卵裂情況,7 d后觀察并統(tǒng)計囊胚發(fā)育情況。
1.4 免疫熒光染色(IF)取出胚胎置于酸性臺式液中反復吹打以去除透明帶,在含0.1% PVA的PBS中清洗3次后轉入4%多聚甲醛中固定30 min。在PBS-PVA中洗滌3次后,將胚胎轉入0.1%Triton X-100中透化30 min(若一抗為5mC和5hmC,透化之后應先置于4 mol/L HCl中處理30 min后轉入Tris-HCl中透化20 min);在PBS-PVA中洗滌3次后轉入含有2%BSA的PBS中室溫封閉1 h或4℃過夜,隨后轉入一抗中室溫孵育1.5 h(一抗用含有2%BSA的PBS稀釋,稀釋比例見表1); 在PBS-PVA中洗滌3次后,將胚胎轉移至Alexa Fluor 488山羊抗小鼠(1∶500稀釋液,A-11001,Invitrogen,MA,美國)或Alexa Fluor 594山羊抗兔(1∶500 稀釋液,A-11037,Invitrogen,MA,美國)中避光室溫孵育1 h;在PBS-PVA中洗滌3次,置于DAPI中染色10~15 min; 最后將胚胎轉移至載玻片上,并用抗熒光淬滅劑處理后在熒光顯微鏡下觀察并拍照。

表1 免疫熒光染色所用抗體
1.5 RNA提取、反轉錄和RT-qPCR收集對照組及5-HT處理組中不同發(fā)育時期(2-細胞、 4-細胞和囊胚)的胚胎,利用RNeasy Mini Kit(Qiagen,Hilden,德國)及反轉錄試劑盒(TransScript All-in-One First-Strand cDNA Synthesis SuperMix),按照說明書進行微量RNA提取及反轉錄。將所得cDNA作為模板,進行熒光定量PCR。表2列出了熒光定量PCR所用的引物。熒光定量PCR反應體系為10 μL SYBR Green Master Mix,1 μL上游引物和1 μL下游引物,1 μL cDNA和7 μL RNase-free water。反應條件:95℃預變性1 min,之后40個循環(huán)擴增程序(95℃ 15 s,60℃ 30 s,72℃ 30 s),然后進行溶解曲線分析(95℃ 60 s,60℃ 60 s,95℃ 1 s),最后冷卻至4℃。結果使用2-△△Ct方法進行計算。

表2 熒光定量PCR所用引物信息
1.6 TUNEL染色收集對照組及5-HT處理組的囊胚,置于酸性臺式液中反復吹打以去除透明帶,在含0.1%PVA的PBS中清洗3次后轉入4%多聚甲醛中固定30 min。在PBS-PVA中洗滌3次后,利用TUNEL凋亡染色試劑盒(InSituCell Death Detection Kit)進行染色,之后在PBS-PVA中洗滌3次,將囊胚轉移到DAPI染色液中孵育10 min。將胚胎轉移至載玻片上,并用抗熒光淬滅劑處理后在熒光顯微鏡下觀察并拍照。
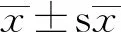
2 結果
2.1 5-HT對豬孤雌胚胎發(fā)育能力的影響與對照組相比,500,1 000 μmol/L 5-HT處理組的卵裂率和囊胚率極顯著降低(P<0.01)(圖1)。由于經(jīng)過1 000 μmol/L 5-HT處理的胚胎無法發(fā)育到囊胚階段,則選擇500 μmol/L作為后續(xù)研究的最佳濃度。
A.對照組及處理組的卵裂率變化;B.對照組及處理組的囊胚率變化;C.24,48 h及6 d時胚胎的發(fā)育圖像(40×)。*P<0.05;**P<0.01。下同
為了評估5-HT處理對胚胎質量的影響,收集對照組及處理組的囊胚利用TUNEL試劑盒進行凋亡分析,測量總細胞數(shù)和凋亡率。TUNEL染色結果如圖2所示,5-HT處理組的凋亡細胞數(shù)明顯高于對照組,而囊胚總細胞數(shù)顯著降低(P<0.01)。

A.囊胚凋亡細胞染色(200×);Bright.明場;Merge.合并;B.囊胚總細胞數(shù)及凋亡細胞數(shù)的變化
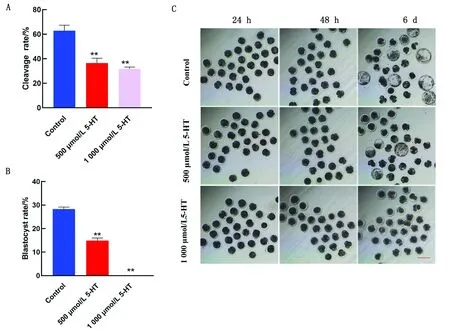
2.2 5-HT對豬孤雌胚胎組蛋白甲基化的影響免疫熒光染色結果表明,與對照組相比,5-HT組的H3K4me2及H3K4me3水平在2-細胞期顯著降低(P<0.01),在4-細胞期H3K4me2的水平有所提高,在囊胚期逐漸減弱(P<0.05)(圖3,4)。 熒光定量PCR結果顯示,在添加500 μmol/L 5-HT處理的2-細胞期胚胎中,去甲基酶KDM1A和KDM5B的含量明顯增加,而在4-細胞期顯著下降,最后在囊胚期顯著上升(圖5)。

A.通過免疫熒光染色檢測H3K4me2水平(200×);Bright.明場;Merge.合并;Blastocyst.囊胚;B.H3K4me2熒光強度分析

A.通過免疫熒光染色檢測H3K4me3水平(200×);B.H3K4me3熒光強度分析
A.KDM1A的mRNA水平;B.KDM5B的mRNA水平
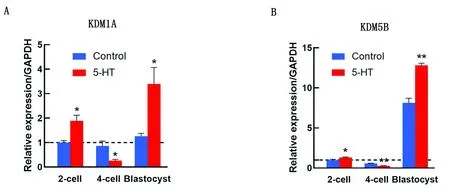
2.3 5-HT對豬孤雌胚胎DNA甲基化水平的影響根據(jù)免疫熒光染色和相對熒光強度的結果,對照組和5-HT組在2-細胞、4-細胞和囊胚期的5mC和5hmC信號水平均未觀察到顯著性差異(圖6)。
A.通過免疫熒光染色檢測DNA甲基化水平(200×);5mC.5甲基胞嘧啶;5hmC.5羥甲基胞嘧啶;B.5mC熒光強度分析;C.5hmC熒光強度分析
2.4 5-HT對多能性基因及凋亡基因的影響熒光定量PCR結果如圖7所示,5-HT處理組多能性基因SOX2的mRNA水平在2-細胞、4-細胞及囊胚期顯著上升,NANOG的mRNA水平在2-細胞、4-細胞及囊胚期顯著下降,促凋亡基因BAX在4-細胞及囊胚期顯著上升(P<0.01),而抑凋亡基因BCL2則無顯著性差異。
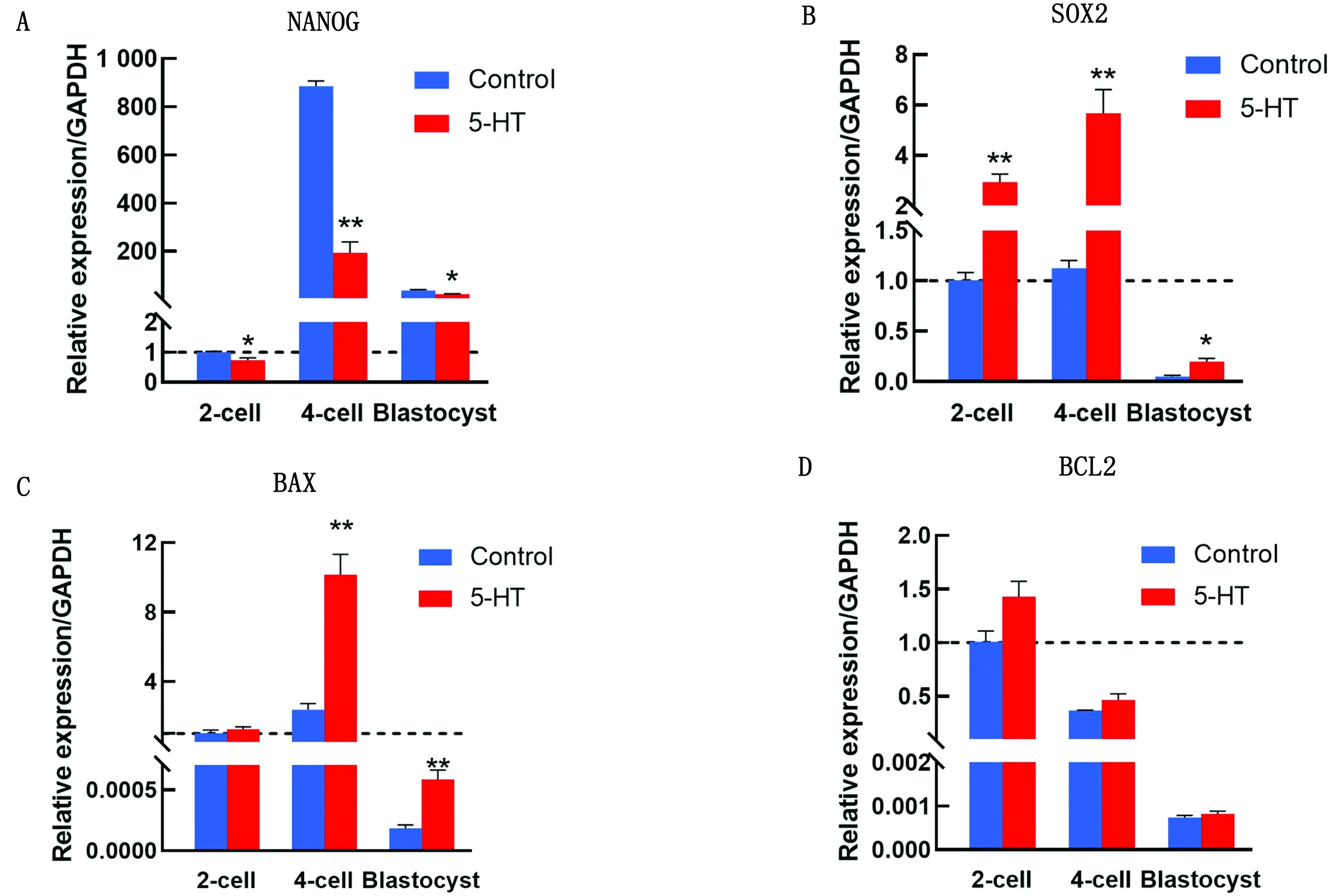
A.多能性基因NANOG的mRNA水平;B.多能性基因SOX2的mRNA水平;C.促凋亡基因BAX的mRNA水平;D.抑凋亡基因BCL2的mRNA水平
3 討論
5-HT在不同的發(fā)育過程中起重要作用,包括細胞增殖、分化,卵母細胞成熟及胚胎發(fā)育。LEONOV等[15]首次發(fā)現(xiàn)1 g/L的5-HT會抑制胚胎的發(fā)育。SAKHAROVA等[16]發(fā)現(xiàn)利用5 μmol/L 5-HT預處理后在4-細胞階段冷凍保存的胚胎活力明顯增強。IL′KOV等[17]觀察到1 μmol/L 5-HT顯著抑制小鼠植入前胚胎發(fā)育,增加凋亡率。本試驗采用500,1 000 μmol/L 5-HT對豬植入前胚胎進行處理,發(fā)現(xiàn)其顯著降低卵裂率及囊胚率,增加凋亡細胞數(shù),這與5-HT對小鼠胚胎的抑制作用是一致的。此外,5-HT處理增加了促凋亡基因BAX的轉錄水平,進一步證實5-HT對豬植入前胚胎發(fā)育具有抑制作用。
已有研究發(fā)現(xiàn)5-HT對哺乳動物植入前胚胎發(fā)育具有抑制作用,但其對胚胎表觀遺傳修飾的作用鮮有研究。本試驗中5-HT處理后H3K4me2和H3K4me3水平在2-細胞階段顯著降低,然后在4-細胞階段升高,而在囊胚階段逐漸恢復。H3K4me2和H3K4me3的動態(tài)變化主要受組蛋白去甲基化酶KDM1A調控,而組蛋白去甲基化酶KDM5B可以調控早期胚胎中H3K4me3水平[18-19]。5-HT可以通過轉谷氨酰胺酶2(TGM2)與胞質蛋白共價連接,進而附著到組蛋白上(組蛋白5-羥色胺化),而這一過程會破壞DNA與組蛋白的結合,并調控基因表達,這表明5-HT與組蛋白修飾之間存在密切的聯(lián)系[20-21]。組蛋白甲基化在早期胚胎發(fā)育和細胞命運中起著至關重要的作用[22-23]。在真核細胞的染色質結構中,組蛋白H3賴氨酸4的甲基化與基因轉錄激活和轉錄沉默密切相關, 例如,H3K4me2可以募集HDAc以抑制轉錄起始,而H3K4me3主要吸引轉錄調節(jié)因子,標記活性基因的轉錄起始位點,并通過調節(jié)染色質來促進基因轉錄[24]。本試驗結果顯示KDM1A和KDM5B的表達變化與H3K4me2和H3K4me3的水平保持一致,表明5-HT處理可通過影響組蛋白甲基化來抑制胚胎發(fā)育。本研究發(fā)現(xiàn)5-HT處理可影響豬孤雌胚胎組蛋白甲基化水平,為豬雌配子在胚胎發(fā)育的表觀遺傳調控提供了新的見解。
在哺乳動物胚胎發(fā)育的早期,表觀遺傳的動態(tài)變化在全基因組轉錄中起著至關重要的作用,從而導致了對諸多基因如多潛能相關基因的研究[25]。本研究發(fā)現(xiàn),5-HT處理顯著提高了SOX2 mRNA表達水平,但降低了NANOG mRNA的表達水平。因此,可推測多能性基因啟動子處組蛋白修飾的動態(tài)變化可能調節(jié)自身及其下游基因的表達,從而影響豬的早期胚胎發(fā)育。然而,在胚胎發(fā)育過程中父源基因組對5-HT的影響還有待于進一步的研究,需進一步探討5-HT對豬體外受精胚胎發(fā)育的影響及組蛋白修飾的調控。
綜上所述,5-HT對豬的植入前胚胎發(fā)育起著至關重要的作用,5-HT可能通過改變組蛋白修飾從而調節(jié)基因表達,進而抑制豬孤雌胚胎的發(fā)育。本研究為哺乳動物體內5-HT、組蛋白修飾及胚胎發(fā)育三者之間的關系提供了新的見解。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
