攻胎飼糧中添加25(OH)D3對高胎齡母豬及其初生仔豬血清激素和抗氧化指標的影響
2021-08-10 07:27:08崔建鑫楊艷陽張卓煒鄧紀昌黃坤玄唐兆新
中國獸醫學報 2021年6期
崔建鑫,楊艷陽,喬 娜,張卓煒,鄧紀昌,黃坤玄,楊 帆,李 英*,唐兆新*
(1.華南農業大學 獸醫學院,廣東 廣州 510642;2.江西農業大學 動物群發性疾病監測與防控研究所,江西 南昌 330045)
國內現代瘦肉型母豬養殖使用年限是3~4年,平均年產2.2~2.5胎,年提供仔豬一般水平為20頭(國外可達到30頭左右)。增加母豬生產胎次和提高高胎齡母豬所產仔豬體質可以有效提高母豬的利用率。根據養豬業的發展現狀,提高母豬胎次,加強仔豬體質迫在眉睫。鈣是影響母豬繁殖的重要因素,在胚胎著床、發育中起著重要作用,同時高臍帶血鈣可能有助于胚胎的發育[1-3]。妊娠期間,母體需要更多的鈣,但隨著年齡的增加,高胎齡母豬整體機能減弱,鈣攝取能力降低,且經過多次生產后更易出現缺鈣現象[4-5]。維生素D是一種脂溶性維生素,可以增強小腸對鈣的吸收,被稱作補鈣劑。與口服維生素D相比,口服25(OH)D3是一種更加安全高效的補充維生素D的方式[6]。這種新型的維生素D在2007年被允許用到豬飼料中[7],我國農業部第2045號公告將25(OH)D3列入《飼料添加劑品種目錄(2013)》,準許25(OH)D3在豬生產上作為維生素及類維生素類飼料添加劑使用。近年來的研究認為在飼糧中添加25(OH)D3可以提高母豬的生產性能[8],但在高胎齡母豬及高胎齡缺鈣母豬攻胎飼糧中添加25(OH)D3對母豬及仔豬體內激素及抗氧化水平產生的影響尚不清楚。本試驗將40頭妊娠85 d 的母豬按照飼糧中鈣含量及是否添加25(OH)D3隨機分為4組,連續飼喂至分娩前1 d,探究攻胎飼糧中添加25(OH)D3對高胎齡母豬及初生仔豬血清激素及抗氧化水平的影響,為高胎齡母豬及缺鈣高胎齡母豬攻胎飼糧的配制提供科學依據。
1 材料與方法
1.1 試驗設計試驗選用健康狀況良好、遺傳背景相似、預產期均衡的7胎齡妊娠母豬40頭,隨機分為4組,每組10個重復。試驗日糧配比在符合NRC(2012)標準配比的基礎上,按照試驗要求有目的的對飼料鈣、磷和25(OH)D3的含量進行調整并將其分為對照組、低鈣組、25(OH)D3組和低鈣+25(OH)D3組。飼料組成及其營養水平見表1。試驗過程中,鈣、磷和25(OH)D3添加情況見表2。

表1 基礎飼糧組成及營養水平(風干基礎) %

表2 試驗飼料鈣、磷和25(OH)D3添加水平
1.2 飼養管理試驗在廣州韶關翁源縣豐澤畜牧養殖有限公司豬場進行。在母豬妊娠85~114 d按照試驗分組飼喂母豬,85~90 d每頭母豬每日飼喂2 kg,91 d至分娩,每頭母豬每天飼喂3 kg,懷孕110 d,轉入產房。母豬分娩后,改用母豬哺乳料。產房溫度控制在20℃,試驗期間按豬場日常管理程序給試驗豬喂料、飲水和免疫。
1.3 母豬血樣采集分別在母豬妊娠85,114 d 耳緣靜脈采血,新鮮血液于4℃放置30 min,3 000 r/min 離心15 min,收集上清,分裝于0.5 mL離心管,液氮保存備用。
仔豬血樣:仔豬初生2 h內,前腔靜脈采集血液。血清收集和儲存方法與母豬血樣相同。
1.4 試驗材料及儀器雌二醇(E2)、豬促紅細胞生成素(EPO)、豬甲狀旁腺素(PTH)、豬皮質醇(Cor)和豬甲狀腺激素(T4)酶聯免疫分析試劑盒均購自上海酶聯公司。過氧化氫酶(CAT)、超氧化物歧化酶(SOD)、谷胱甘肽過氧化物酶(GSH-Px)、血清丙二醛(MDA)和活性氧(ROS)試劑盒購自南京建成。Elx 808型酶標儀,購自Bio Teck公司(美國);Nanodrop2000型,微量紫外分光光度計,購自Thermo Fisher科技公司(美國)。
1.5 母豬及仔豬血清激素水平檢測母豬及仔豬血清冰上融化后,離心,使用酶聯免疫試劑盒檢測母豬血清中E2、EPO、PTH、Cor和T4激素水平和仔豬血清中EPO、PTH、Cor和T4激素水平。檢測過程嚴格按照試劑盒說明書進行。
1.6 母豬及仔豬血清抗氧化水平檢測母豬及仔豬血清冰上融化后,離心,采用酶標儀法測定血清中MDA含量、ROS水平以及SOD的活力,其中ROS水平用熒光強度值(FIV)表示。微孔法檢測血清中CAT、GSH-Px的活力。嚴格按照試劑盒說明書操作。

2 結果
2.1 攻胎飼糧添加25(OH)D3對高胎齡母豬血清激素的影響檢測結果如表3所示,母豬妊娠85 d血清內E2、EPO、PTH、T4、Cor激素在各組之間均無顯著性差異(P>0.05)。表4中的結果表明,與對照組相比,母豬妊娠114 d時的25(OH)D3組E2激素含量顯著降低(P<0.05),但EPO激素含量顯著上升(P<0.05)。妊娠114 d,母豬血液內T4激素含量均有升高,但各組之間均無顯著性差異(P>0.05);低鈣組母豬血清中PTH含量顯著高于25(OH)D3組和低鈣+25(OH)D3組(P<0.05),但與對照組相比沒有顯著性差異(P>0.05)。Cor激素在低鈣+25(OH)D3組含量最高,且顯著高于25(OH)D3組(P<0.05),除此之外,其他各組之間均差異不顯著(P>0.05)。主效應分析表明,飼糧中添加25(OH)D3可顯著提高母豬EPO的激素水平(P<0.05),低鈣的情況下可造成Cor含量上升。

表3 各組妊娠85 d母豬血清中激素表達水平
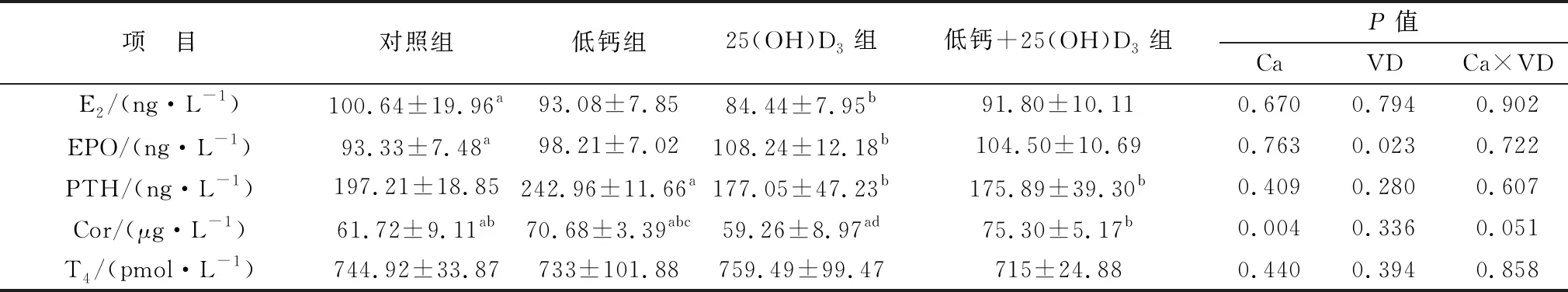
表4 各組妊娠114 d母豬血清中激素表達水平
2.2 攻胎飼糧中添加25(OH)D3對仔豬血清激素的影響初生仔豬血清激素檢測結果如表5所示,25(OH)D3組仔豬血清中EPO含量顯著高于對照組(P<0.05),低鈣+25(OH)D3組EPO含量顯著低于低鈣組和25(OH)D3組(P<0.05)。仔豬血清T4含量在各組之間均無顯著性差異(P>0.05)。與對照組相比,低鈣組、25(OH)D3組和低鈣+25(OH)D3組血清PTH激素含量均顯著升高(P<0.05)。Cor含量在25(OH)D3組顯著低于其他3個試驗組(P<0.05)。主效應分析表明,鈣和25(OH)D3聯合能顯著降低EPO激素水平的表達(P<0.05),鈣能顯著影響Cor激素水平。

表5 各組初生仔豬血清中激素表達水平
2.3 添加25(OH)D3對高胎齡母豬血清抗氧化水平的影響如表6所示,母豬妊娠85 d,抗氧化各指標在各組之間均無顯著性差異。表7所示,相比于對照組,妊娠母豬114 d時,25(OH)D3組與低鈣組的ROS水平顯著降低(P<0.05),但低鈣+25(OH)D3組ROS含量高于兩者。妊娠114 d母豬血清中,25(OH)D3組和低鈣+25(OH)D3組血清中SOD、CAT、GSH-Px的含量顯著高于對照組和低鈣組(P<0.05),與此同時,低鈣組中CAT含量與對照組相比顯著降低(P<0.05)。MDA水平在各組之間均沒有顯著變化(P>0.05)。主效應分析表明,在飼料中添加25(OH)D3可顯著提升SOD、CAT、GSH-Px的表達量(P<0.05)。
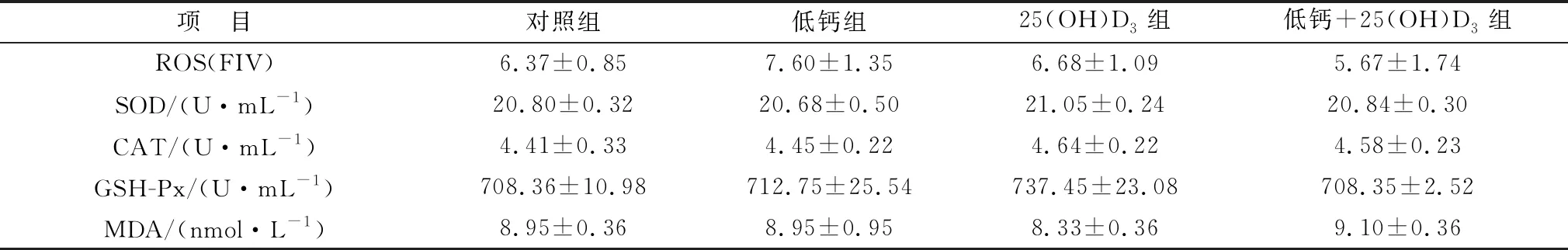
表6 各組妊娠85 d母豬血清中抗氧化指標水平
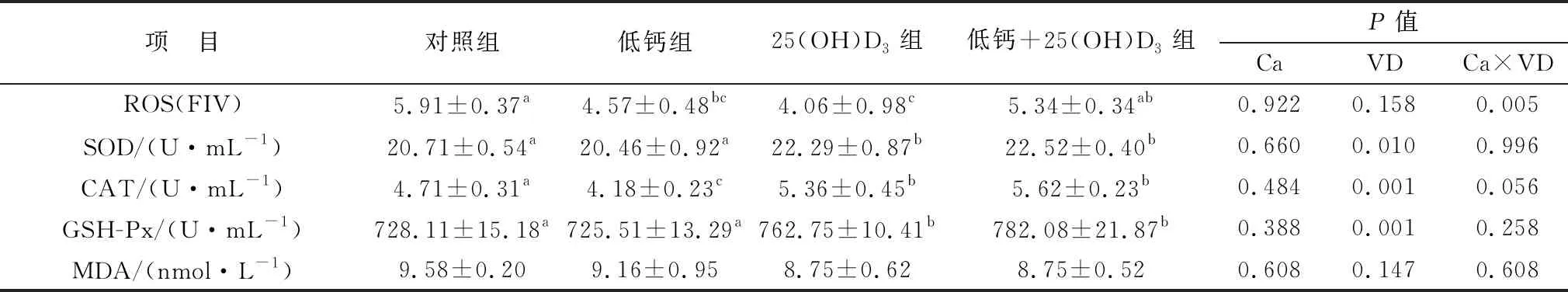
表7 各組妊娠114 d母豬血清中抗氧化指標水平
2.4 母豬飼糧中添加25(OH)D3對仔豬血清抗氧化水平的影響如表8所示,初生仔豬低鈣組和低鈣+25(OH)D3組血清中ROS水平顯著低于對照組(P<0.05)。與母豬血清激素抗氧化水平檢測結果相同,初生仔豬25(OH)D3組和低鈣+25(OH)D3組血清中SOD、CAT的含量顯著高于對照組和低鈣組(P<0.05)。GSH-Px和MDA水平在各組之間均沒有顯著變化(P>0.05)。主效應分析結果表明,25(OH)D3可以顯著提高仔豬的抗氧化水平。
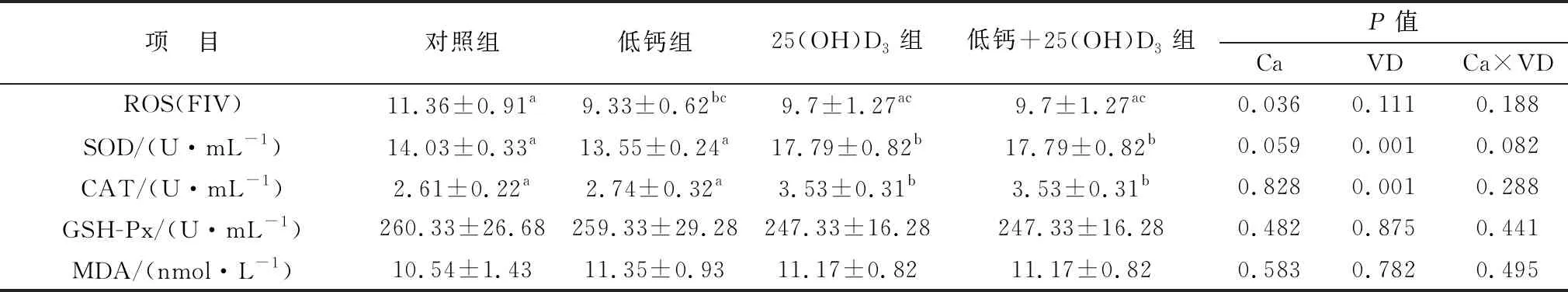
表8 各組初生仔豬血清中抗氧化指標水平
3 討論
3.1 攻胎飼糧中添加25(OH)D3對母豬及仔豬血清激素的影響膽骨化醇是補充維生素D常見的渠道,腸黏膜吸收膽骨化醇之后轉變為25(OH)D3,繼而轉變為1,25-二羥基維生素D3(1,25(OH)2D3)。激素對機體的生長、發育、繁殖、代謝等起著重要的調節作用,適當的調節激素水平對機體生長發育等有促進作用[9-10]。鈣穩態是通過腸道吸收、腎臟排泄和骨代謝來調節的,這些過程受PTH、性激素和維生素D等調控[11]。PTH作為與機體生長發育至關重要的激素之一,其含量與機體內鈣和25(OH)D3的含量密不可分。機體可通過調節PTH的含量來控制血清內25(OH)D3與鈣的平衡,當機體缺乏維生素D時,PTH分泌增多,且血清中的PTH與25(OH)D3含量成反比[12-13]。同時有研究表明[14],PTH能動員鈣從骨組織中釋放到血清中,低鈣會在一定程度上提高PTH的表達量。在本研究中,低鈣組母豬和仔豬血清內PTH含量最高,且低鈣組母豬血清內PTH含量顯著高于25(OH)D3組和低鈣+25(OH)D3組,同時低鈣組仔豬血清PTH含量顯著高于其他3個試驗組,本試驗結果對PTH、鈣、維生素D之間的相互影響做了印證。4個試驗組中,25(OH)D3組的E2激素表達量最低并顯著低于對照組,低鈣+25(OH)D3組表達量也低于對照組,但差異不顯著。母體妊娠8周之后,E2主要由胎兒-胎盤分泌,且妊娠后期母體E2水平上升會導致胎兒獲得的營養物質減少,引起胎兒T4水平下降,生長速度下降[15,16]。而本試驗中,各試驗組間母豬和仔豬血清T4含量均沒有顯著性差異。且母豬25(OH)D3組血清內T4含量最高,低鈣組及低鈣+25(OH)D3組依次低于對照組;仔豬25(OH)D3組血清內T4含量最低,低鈣+25(OH)D3組及低鈣組依次高于對照組。陳勝等[15]在對兔的研究中發現,母兔TH水平上升對初生仔兔TH水平無顯著影響。但BUZZARD等[17]認為,由于種間成熟期以及胎盤屏障的通透性的差異性,其結果會有一定的差異。根據試驗結果推測機體內鈣和25(OH)D3的含量對T4水平沒有顯著的影響。
促紅細胞生成素(EPO)是由腎皮質腎小管周圍間質細胞和肝臟細胞分泌的一種激素類物質,參與紅細胞前體細胞的分化與成熟。KIM等[18]發現EPO在體內可通過激活JAK/STAT信號通路促進骨形態蛋白(BMP2)的產生和骨形成,同時EPO在體外可以直接激活間充質細胞分化為成骨細胞[18]。用25(OH)D3喂養高胎次母豬可減少骨代謝,改善骨狀態,同時也可增強初生仔豬的生長性能及骨狀態[19]。本試驗結果表明,25(OH)D3組母豬及仔豬血清內EPO激素水平顯著高于對照組,但是低鈣組母豬及仔豬血清中EPO含量均高于對照組,但差異不顯著。對各組EPO的表達量進行分析后我們發現,25(OH)D3在母豬血清EPO的變化中起主效應;而在仔豬血清中鈣和25(OH)D3聯合作用起主效應,但對于仔豬低鈣組EPO表達量的升高,推斷缺鈣初期機體會代償性產生較多的EPO,但其作用機制還需要進一步深入研究。Cor是一種內源性的糖皮質激素,參與哺乳動物各種復雜的生物效應,如生長、免疫反應和新陳代謝,在機體受到壓力時會升高。試驗結果顯示,25(OH)D3組母豬與仔豬血清中Cor含量最低,由此推斷25(OH)D3可以減輕母豬孕期壓力。對各組母豬血清內Cor含量進行分析發現,鈣在Cor的表達中起主效應。
3.2 攻胎飼糧中添加25(OH)D3對母豬及仔豬血清抗氧化水平的影響ROS被認為是生成ATP過程中的氧化代謝產物,ROS的積累超過細胞抗氧化能力水平時會損傷細胞,有研究表明Ca2+與ROS有一定的關聯[20-21]。母豬缺鈣會使線粒體呼吸會受到一定程度的抑制,導致ROS含量降低[22]。本試驗結果顯示妊娠母豬低鈣組血清中的ROS含量顯著低于對照組,同時妊娠114 d的母豬25(OH)D3組血清內的ROS含量低于低鈣組,推測其可能是因為Ca2+參與呼吸鏈中部分酶的合成,誘導ROS的產生。機體抗氧化水平與Ca2+的攝取量呈正相關,同時也受到1,25(OH)2D3激素的調控[23]。目前發現1,25(OH)2D3能增加抗氧化酶、解耦連蛋白2(CUP2)、CUP3的表達[24]。UCP可抑制電子傳遞鏈中電子泄露,降低能量代謝中產生的自由基[25]。
SOD是一種需氧生物中以超氧陰離子為底物的酶,也是一類重要的氧自由基清除劑,妊娠后逐漸增加。CAT是一種機體內廣泛存在的一類末端氧化酶,能催化H2O2分解,減少羥自由基(·OH)的產生,清除體內ROS,減輕其對細胞的損害[26]。GSH-Px是機體內存在的含硒清除自由基和抑制自由基反應的系統,可以清除ROS和酯質過氧化物。本試驗結果顯示,25(OH)D3組在母豬及仔豬血清中SOD、CAT的活力顯著高于對照組與低鈣組,而低鈣+25(OH)D3組SOD、CAT的活力處于25(OH)D3組和低鈣組之間,且25(OH)D3對SOD、CAT的活力影響起主效應,同時母豬血清中GSH-Px有著相同的結果;表明25(OH)D3可以有效改善高胎齡母豬及初生仔豬的抗氧化能力。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
學苑創造·A版(2015年11期)2016-01-14 09:03:27
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
