江蘇東臺開莊遺址動物遺存研究報告
2021-07-17 07:21:38乙海琳甘恢元許晶晶
東南文化 2021年3期
乙海琳 甘恢元 許晶晶
(1.杜倫大學 英國杜倫;2、3.南京博物院 江蘇南京 210016)
內容提要:開莊遺址2018-2019年的發掘工作中收獲了豐富的動物遺存,包含龜科、大雁、狗、熊、豬、梅花鹿、水鹿、麂、獐、麋鹿、水牛等,尤其以麋鹿等野生動物種類占優。對H13性質的分析及麋鹿各部位出現頻率的討論顯示出開莊遺址和其他遺址之間可能存在搬運行為或其他聯系。生業經濟方面,開莊遺址良渚晚期是以飼養家豬和狩獵野生動物相結合的模式,而到了龍山文化時期,則迅速轉變為大量采集狩獵的資源獲取模式。
一、遺址基本情況介紹
開莊遺址位于江蘇省東臺市溱東鎮開莊村。1995年,當地磚瓦廠取土首次發現該遺址[1];2018年底,由南京博物院聯合東臺市博物館對開莊遺址進行考古勘探并發掘。此次發掘,共發現各類遺跡51個(組),出土各類遺物200余件[2]。
本文將對開莊遺址出土的動物遺存進行介紹和分析,近現代地層的動物遺存只提供部分數據(表一)。
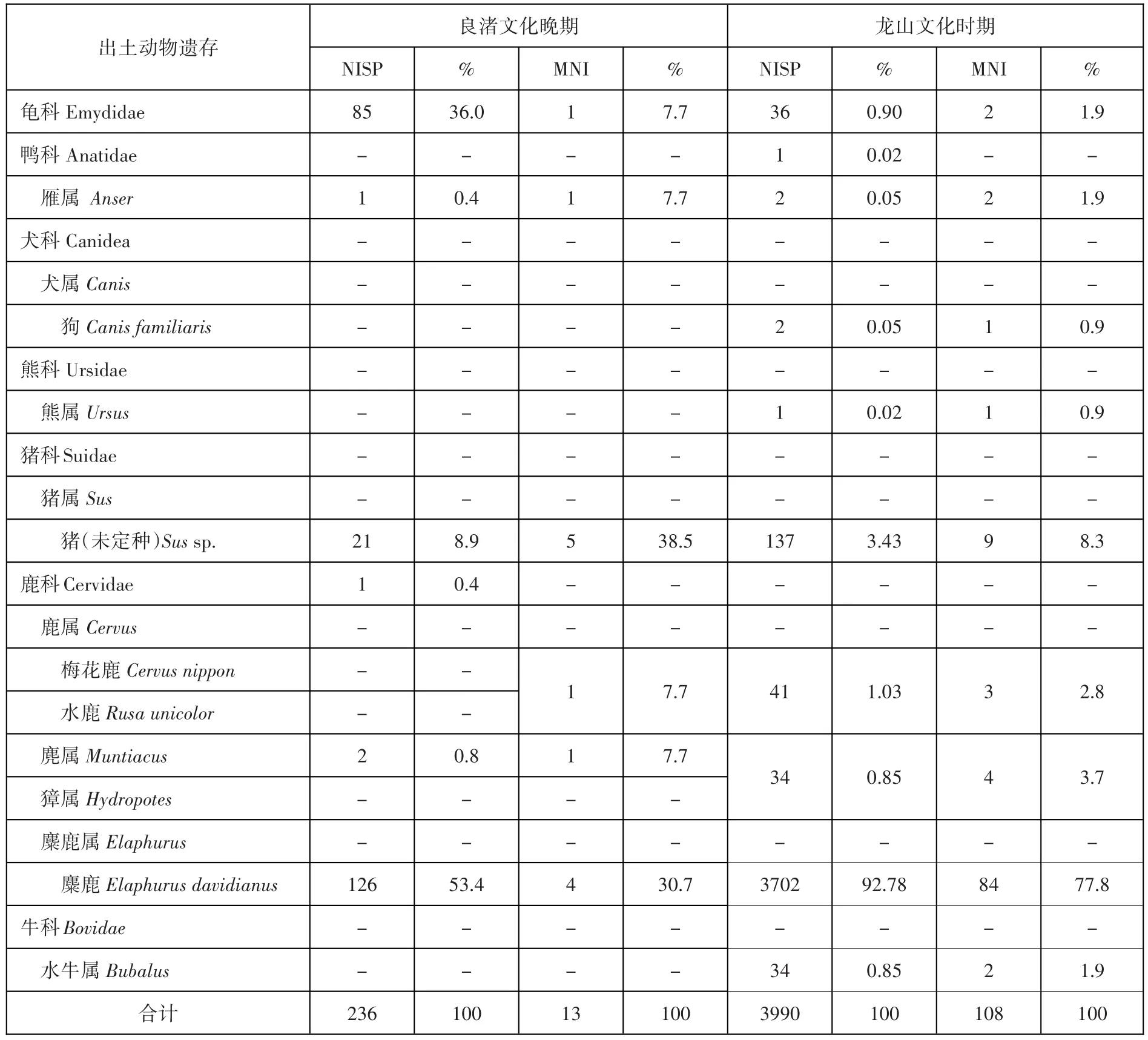
表一// 開莊遺址新石器時代動物骨骼可鑒定標本數(NISP)和最小個體數(MNI)統計表(單位:個)
二、材料與方法
開莊遺址2018—2019年度的發掘共發現動物遺存9384件,總重量約238965克。這些動物遺存絕大多數出土于新石器時代地層,包含良渚文化晚期和龍山文化時期兩個部分,其中又以龍山文化地層的堆積較為豐富。由于發掘區域較為集中,無法進一步界定同時期不同遺跡單位中動物遺存的獨立性(例如同屬良渚文化晚期的兩個灰坑,因其在空間上相距較近,很難判斷兩個灰坑中的動物骨骼是否來自于同一動物個體),因此本文將按照良渚文化和龍山文化兩個大的時期,對該遺址出土的動物遺存進行分類和統計。
種屬鑒定工作主要參照《動物骨骼圖譜》[3]、《考古遺址中的鳥骨鑒定手冊》[4]等動物骨骼圖譜及部分現生標本照片;動物的性別年齡鑒定主要參照《考古遺址中動物骨骼的年齡性別》[5];動物骨骼的尺寸數據測量參照《考古遺址出土動物骨骼測量指南》[6]等書中的標準。測量工具使用山東大學動物考古實驗室統一采買的電子數顯卡尺,稱重使用電子秤,并制作標簽。該遺址出土的動物種屬按照林奈生物分類法分類。
(一)良渚文化晚期動物遺存概況
該時期出土動物遺存349件,其中113件僅能鑒定為哺乳動物,根據其尺寸可大致分為大型哺乳動物(85件)和中型哺乳動物(28件)。其余可鑒定到目一級的動物遺存共計236件,包含龜鱉目、雁形目和偶蹄目。
1.龜鱉目(Testudinata)
龜科(Emydidae)腹甲殘片85件,至少代表1個個體。
2.雁形目(Anseriformes)
雁屬(Anser)僅有1件左側股骨近端。
3.偶蹄目(Artiodactyla)
(1)豬科(Suidae)
豬屬(Sus)共計21件,為上下頜及牙齒、枕骨、肩胛骨、肱骨、尺骨、髖骨等,至少代表不同年齡段的5個個體。
(2)鹿科(Cervidae)
麂屬(Muntiacus)共計2件標本,為下頜骨和角。
中型鹿,1件,為左側下頜游離M3。
麋鹿(Elaphurus davidianus)126件,為角、頭骨帶角、枕骨、上下頜及牙齒、樞椎、肩胛骨、肱骨、尺骨、橈骨、腕骨、掌骨、髖臼、股骨、髕骨、脛骨、距骨、跟骨、中央跗骨、跖骨、趾骨等。至少代表4個個體。
(二)龍山文化時期動物遺存概況
該時期出土動物遺存共計8999件。1532件標本由于過于破碎或缺少鑒定特征,記為碎骨或殘骨,其主要出于H13內;3476件標本僅能鑒定為哺乳動物,根據其尺寸可分為大型(2367件)、中型(1108件)和小型(1件)哺乳動物;其余可鑒定到目一級的動物遺存共計3991件,包含龜鱉目(36件)、雁形目(3件)、食肉目(4件)和偶蹄目(3948件)。
1.龜鱉目(Testudinata)
龜科(Emydidae)腹甲殘片36件,因花紋有明顯不同,至少代表2個個體。
2.雁形目(Anseriformes)
(1)大型鴨科(Anatidae)肢骨殘段1件。
(2)雁屬(Anser)左側脛骨遠端2件,至少代表2個個體。
3.食肉目(Carnivora)
橈骨近端1件,因無法確定其具體種屬,記為大型食肉動物,該個體可能存在病變。
(1)狗(Canis familiaris)2件,為尺骨和右側下頜帶牙齒,至少代表1個個體。
(2)熊屬(Ursus)僅有1件左側第五掌骨,代表1個個體。
4.偶蹄目(Artiodactyla)
(1)豬科(Suidae)
豬屬(Sus)137件,為頭骨(頂骨枕骨部)、上下頜及牙齒、寰椎、樞椎、肩胛骨、肱骨、尺骨、橈骨、髖骨、股骨、髕骨、脛骨、跟骨、距骨、掌骨、跖骨、趾骨等。至少代表9個個體。
(2)鹿科(Cervidae)
小型鹿34件,其中可以確定具體種屬的是獐屬(Hydropotes)犬齒1件和麂屬(Muntiacus)左側頭骨帶角柄1件,其余部位僅能歸為小型鹿,為下頜、肩胛骨、肱骨、橈骨、股骨、脛骨、掌骨、跖骨、近端趾骨等。至少代表4個個體。
中型鹿41件,其中可以確定種屬的是梅花鹿(Cervus nippon)角殘塊2件和水鹿(Rusa unicolor)自然脫落角環部2件,其余部位骨骼歸中型鹿,為頭骨帶角、角、枕骨、前頜骨、下頜及牙齒、肩胛骨、尺骨、脛骨、跟骨、距骨、掌骨、跖骨、末端趾骨等。至少代表3個個體,其中1個為幼年個體。
麋鹿(Elaphurus davidianus)3702件,為角、頭骨帶角、頭骨殘塊、前頜骨、枕骨、上下頜及牙齒、寰椎、樞椎、頸椎、胸椎、腰椎、薦椎、肩胛骨、肱骨、尺橈骨、尺骨、橈骨、腕骨、掌骨、坐骨、恥骨、髂骨、髖骨、股骨、髕骨、脛骨、距骨、跟骨、跗骨、跖骨、趾骨等。至少代表84個個體,其中70個為成年個體,14個為小于1個月的幼年個體。
(3)牛科(Bovidae)
水牛屬(Bubalus)34件,為上下頜游離臼齒、肋骨、肩胛骨、橈骨、腕骨、掌骨、股骨、距骨、跟骨、跗骨、跖骨、趾骨等。至少代表2個個體。
三、數據分析
開莊遺址2018—2019年度的考古發掘共發現動物遺存9384件,除去36件出自近現代地層和采集的標本,其余9348件動物骨骼均為新石器時代,這是本文的研究對象,下文中所有的“總標本數”“總數”等概念均指新石器時代動物標本總數。其中可鑒定標本數(NISP)為4226件,約占總標本數的45.21%。
從文化分期來看,新石器時代的動物遺存中,有8999件屬龍山文化時期,可鑒定標本數為3990(44.34%);良渚文化晚期僅出土標本349件,可鑒定標本數為236(67.62%)。
從遺址動物群構成來看,該遺址出土的動物骨骼有爬行動物、鳥類和哺乳動物。不論是良渚文化時期還是龍山文化時期,哺乳動物的數量都是最多的,分別占兩個時期標本總數的74.7%和82.0%。哺乳動物中又以大型哺乳動物占絕對主導地位,中型哺乳動物次之,小型哺乳動物僅在龍山文化地層發現1件(圖一、圖二)。
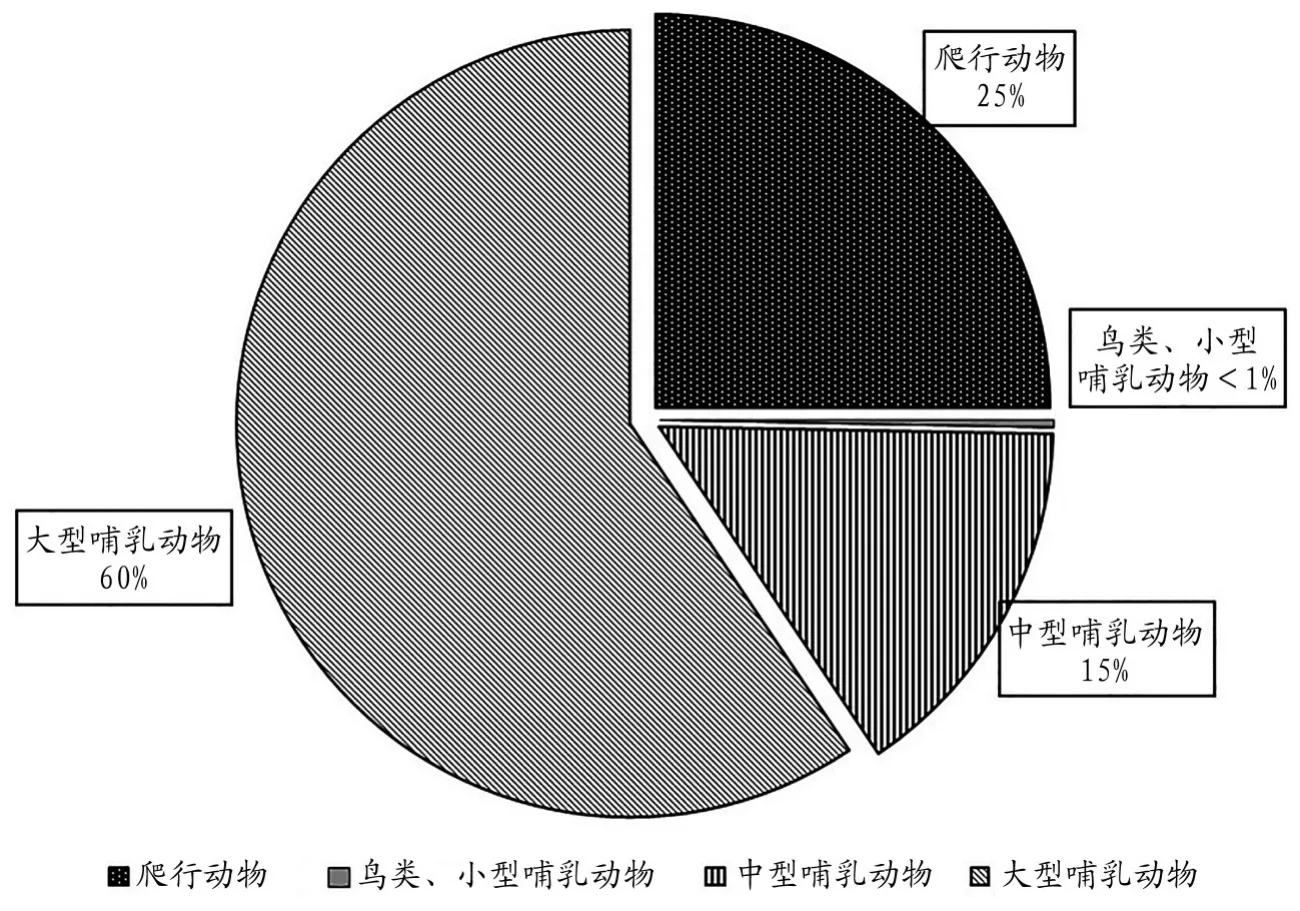
圖一// 良渚文化晚期動物群構成
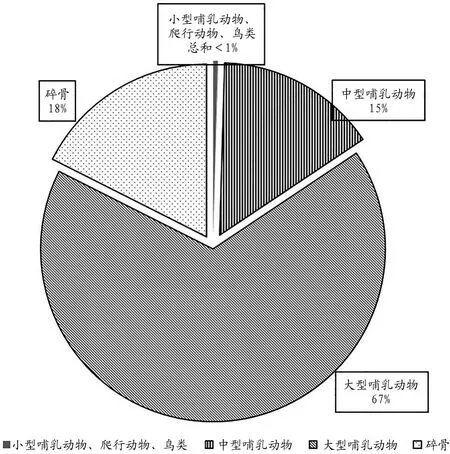
圖二// 龍山文化時期動物群構成
對動物群再細分,新石器時代可鑒定到“科”及以下的標本有龜科、雁屬、狗、熊屬、豬、梅花鹿、水鹿、麂屬、獐屬、麋鹿、水牛屬。良渚文化晚期,麋鹿的可鑒定標本數最大,龜科次之,豬再次之,雁屬、小型鹿、中型鹿的標本數量非常少,而最小個體數則是豬和麋鹿為最多(圖三)。由此可見,麋鹿和豬分別在兩個維度上數量顯著,占據著該時期標本構成的優勢地位,而龜科標本在兩個維度上的落差應該是由于龜腹甲殘塊較多造成的統計誤差。龍山文化時期,種屬之間的數量差異則十分明顯,麋鹿無論是在可鑒定標本數還是最小個體數上都占據著絕對優勢;豬次之;再就是梅花鹿、水鹿、麂屬等小、中型鹿類動物;其他種屬僅有零星的幾件(圖四)。
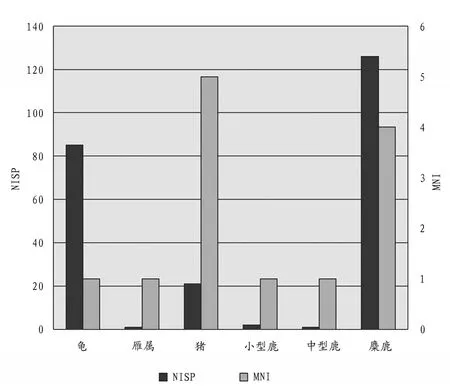
圖三// 良渚文化晚期動物骨骼NISP和MNI統計圖
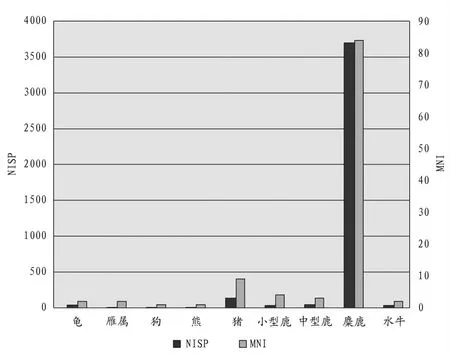
圖四// 龍山文化時期動物骨骼NISP和MNI統計圖
四、討論
(一)古環境復原
開莊遺址位于江淮東部,如今該地區水網密布,地質地貌上以沖積平原及濱海灘涂地為主,缺乏丘陵及山地地帶。
依據均變論原理,可以通過遺址中出現的動物種類來還原該遺址及周邊地區的古環境。龜科可半棲水,喜淡水河流、湖泊及沼澤地;雁屬多為候鳥,以魚、蝦、水草、種子為食,喜在淡水沿水岸水生植物或水岸植物稠密區覓食,淡水域為其活動范圍;野豬可棲息于多樣的環境,但喜歡棲居在茂密的灌叢、潮濕的闊葉林或草地[7];熊多棲息于闊葉林或混交林中,可于低丘活動;獐屬也稱河麂,沿江湖兩岸的濕地、苔草地、芒叢、蘆葦地以及臨近低山、丘陵坡地是獐比較適宜的生存環境,其主要靠蘆葦和高草叢隱蔽;梅花鹿的南方亞種喜棲息于植被以芒、草等草本植物和灌木叢為主的山丘;麋鹿喜溫暖濕潤氣候,其蹄寬大分裂,趾間有皮腱膜,適于在沼澤地行走,牙齒纖弱,適于采食鮮嫩水生植物[8]。
綜上,開莊遺址新石器時代的古氣候環境應該與現在江淮東部地區差不多,整體溫暖濕潤,沼澤廣布,水源豐富。遺址周圍河流、湖泊星羅棋布,從而孕育了大量的水岸水生植物,是喜水鳥類、鹿類動物的天堂。遺址地處平原地區,草地廣布,遠處可能有部分低丘灌木叢以及潮濕的闊葉林,又是野豬、熊、梅花鹿等動物的理想棲息地。
(二)生業模式的探討
通過對開莊遺址出土動物骨骼數量的比對分析,可以清楚地看到,麋鹿在良渚文化和龍山文化兩個時期都占據著優勢地位,應該是當時人們的主要肉食來源。麋鹿及各種體型的鹿類動物,均為野生,說明狩獵野生鹿類動物,尤其是麋鹿,是開莊先民生業經濟的重要組成部分。
遺址中還出土了一定量的豬骨,判斷其是家養還是野生對該遺址的生業經濟研究至關重要。筆者將分別對良渚文化晚期和龍山文化時期的豬骨進行討論。
1.龍山文化時期
因為龍山文化時期的豬骨性質更為清晰明了,所以先對其分析判斷。
從數量比例來看,該時期豬的可鑒定標本數和最小個體數的占比分別僅為3.4%和8.3%,與其他同時期出土家豬的安徽尉遲寺[9],山東尹家城[10]、莊里西[11]、丁公[12]等遺址相比較,其豬骨的數量比例相差甚遠。由于豬的牙齒尺寸會隨著馴化過程而逐漸縮小[13],因此下頜M3尺寸可以作為衡量家豬還是野豬的參考。該時期共發現3件下頜M3標本,分別為:TS1E3③層的游離下頜M31件,M3長44.18毫米;TS1E3⑤a層的左側下頜帶M1-M31件,M3長33.45毫米;TS1E2⑦的右側下頜帶P3-M31件,M3長44.66毫米(圖五)。這三件標本,尤其是兩件M3長度大于44毫米的標本具有極強的野生豬的形態特征,此外這3件標本M3長度的平均數據約為40.76毫米,也超出了一些學者提出的“下頜M3平均長度小于39毫米(即出現家豬)”[14]的標準。從年齡結構來看,該時期可以判斷具體年齡范圍的個體有7個(7—9個月1個;10—12個月1個;15—17個月1個;20—21個月1個;21—23個月2個;23—27個月1個),雖然這些個體的年齡大部分落在了2歲以下,似乎符合家豬飼養的年齡結構,但事實上,還有部分標本由于關鍵牙齒的缺失,只能鑒定到年齡下限,而無法判斷上限,導致有不少“大于18/22/25/29個月”(可能大于2歲的個體)的標本無法被統計在內,從而使得表面上死亡年齡偏小,在實際操作中,2歲以上的個體應該更多,而豬的整體年齡結構是偏大的。
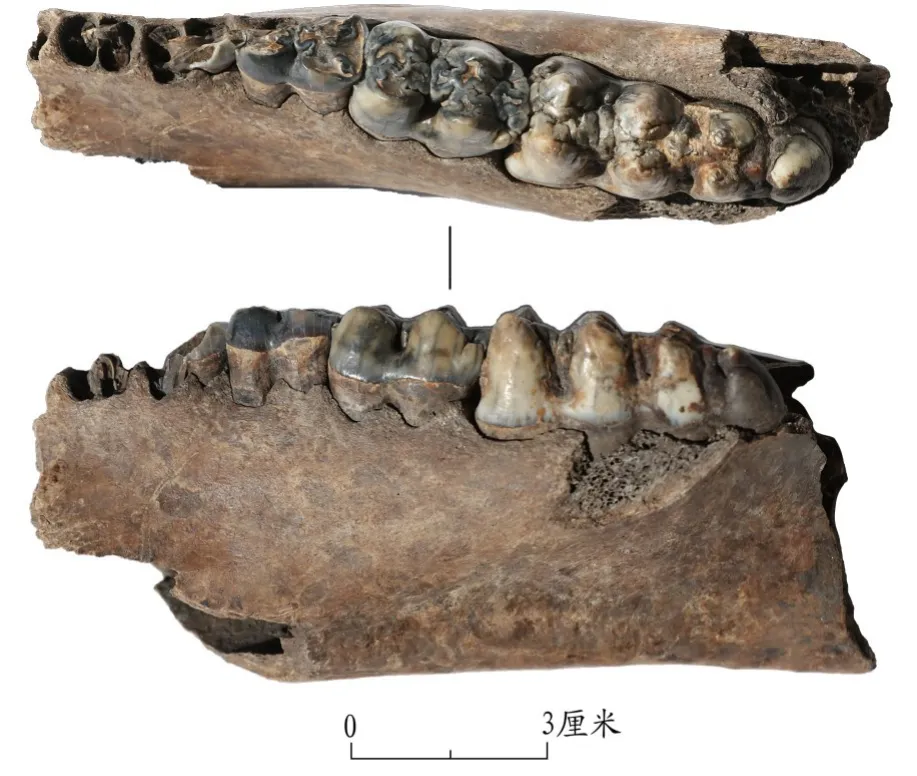
圖五// 龍山文化時期野豬下頜標本
綜上所述,龍山文化時期出土的豬骨應該大概率為野豬,但鑒于遺址年代偏晚,不排除有家豬飼養的可能性。
2.良渚文化晚期
相較于龍山文化時期,良渚文化晚期的動物骨骼雖數量遠不及龍山文化時期,但豬骨的成分更為復雜。相對數量比例方面,該時期豬的最小個體數是最高的,高于麋鹿的比例,占總最小個體數的41.7%,雖然豬骨的可鑒定標本數僅占約8.9%,但這個情況與龜腹甲的統計誤差是息息相關的,由于龜腹甲較為破碎,導致其統計出的可鑒定標本數較大,這樣在一定程度上就會拉低豬骨標本所占的比例。“一個考古遺址出土的主要哺乳動物群中,若豬所占的相對比例(NISP比例、MNI比例或者MW比例都可以)超過30%以上,我們就基本可以肯定該遺址豬群中很可能已經出現了家豬。”[15]基于此,良渚文化晚期的豬骨標本中應該存在家豬。牙齒形態方面,該時期并未發現可以作為牙齒形態參考的下頜M3標本。年齡結構方面,有5個個體可判斷具體年齡,其中7—10個月2個,16—17個月2個,21—23個月1個,全部是小于2歲的個體,符合出現家豬飼養的死亡年齡構成。與龍山文化時期情況不同的是,這是該時期全部的牙齒標本,不存在有潛在的個體被忽略的情況,因此基本可以代表該階段豬的死亡年齡結構。
綜上,良渚文化晚期存在家豬,也可能存在部分狩獵來的野生個體,但應該是以家豬為主。
此外,我們還可以通過對比來考察兩個時期中豬的性質以及生業模式的不同。從良渚文化晚期的豬(NISP占比8.9%,MNI占比41.7%)到龍山文化時期的豬(NISP占比3.4%,MNI占比8.3%),兩個指標均有所下降,而野生麋鹿的數據則直線上升(圖六);龍山文化時期豬的年齡構成整體上比良渚文化晚期的要大,該時期野生形態特征也表現得更為明顯。盡管兩個時期的豬骨性質并不能單純地以野生或家養來定義,但肉食資源性質從良渚文化時期表現出“家養與野生摻雜”到龍山文化時期“野生占主導”,這種變化本身也可以說明問題,且更突出了二者的不同。此外,植物方面的材料也顯示出了相似的規律,與家豬飼養相輔相成的稻作農業,在開莊遺址良渚文化晚期階段延續著江淮東部的傳統并保持著一定強度,而到了龍山文化時期,稻作農業似乎未表現出持續強化的發展態勢[16]。從良渚文化晚期到龍山文化時期,開莊遺址的野生動物資源占比迅速增加,野生植物資源也趨于強勢,是什么原因導致了該地區肉食資源獲取方式的“逆發展”?這值得今后更多地探討。
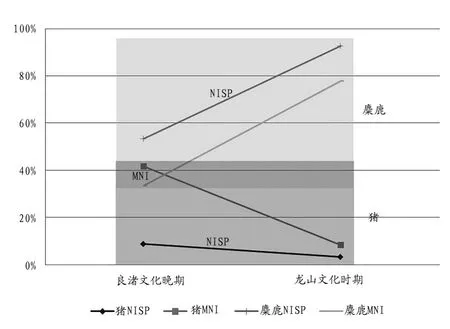
圖六// 開莊遺址良渚晚期到龍山時期豬和麋鹿的數量比例變化
開莊遺址地處水網密布的地勢平緩地區,自然資源極為豐富,資源域范圍很大。動物方面,良渚文化晚期的肉食獲取方式可能是以小規模飼養家豬與狩獵相結合的模式,在飼養家豬的同時會捕獲一些野生動物以補充肉食資源;而龍山文化時期的先民主要用狩獵的方式來獲取資源,以麋鹿為主的大型野生哺乳動物是其主要肉食來源,輔之以水牛、野豬、中小型鹿類動物等。植物方面,采集植物是重要來源,農業在聚落生產中不占絕對優勢。稻作是最重要的農耕生產內容,良渚文化晚期保持著一定強度的稻作農業傳統,而龍山文化時期并未發現持續強化的態勢[17]。骨角器制作方面,人們會將食用后的麋鹿長骨加工成骨鏃、骨鑿等骨器,獐牙制成牙刀,牛肩胛骨制成骨耜,采集或狩獵到的鹿角加工制作為角器。
(三)H13性質的討論
H13是龍山文化時期一個非常特殊的灰坑,僅發掘了北半部。在距開口深約21厘米以下部分,堆填了大量未成年哺乳動物的細碎骨骼[18]。這些動物骨骼共約3000件,除34件屬于成年大、中型哺乳動物之外,其余均屬于未成年個體。整體來看,這些骨骼非常細碎且脆弱,所有部位的骨骺均未愈合(圖七),沒有發現鹿角。綜合各個部位的尺寸和特征,判斷這些骨骼應屬于大型鹿,再結合遺址中出土了大量麋鹿的信息,認為其應該為麋鹿。另外,發現了27件(左14、右13)保存較好的距骨,基于此,H13中的未成年骨骼遺存至少代表了14個未成年麋鹿個體。

圖七// H13未成年麋鹿骨骼愈合狀況
對這14個未成年麋鹿個體的年齡判斷是進行深入研究的關鍵。參考唐納德·查普曼(D.I.Chapman)等對黃麂年齡判斷的指標[19],日本學者Hiroko Koike和Noriyuki Ohtaishi關于梅花鹿牙齒萌出時間的標準[20],以及針對馬鹿[21]的牙齒萌出時間研究等,這14個未成年麋鹿個體的年齡應該處于圍產期,即出生前后,它們可能是剛出生約1周的新生兒,或者是仍未出生的胎兒。最能說明這個問題的是中央跗骨標本,隨著動物體重的增加,跗骨愈合生長的速度非常快,但如圖七∶3所示,中央跗骨兩半并未開始愈合且形狀非常小,因此這些未成年小鹿的年齡確實非常年輕。而下頜牙齒的萌出情況也證實了這一點(圖八)。

圖八// H13未成年麋鹿牙齒萌出狀況
這些麋鹿新生兒/胎兒似乎并沒有被人們所利用,而是被直接丟棄在了H13之中。綜合整個遺址的動物骨骼情況來看,僅在H13中發現了圍產期麋鹿骨骼的集中堆積,除了H39中發現的1件小于1個月的麋鹿下頜,其余遺跡單位并未發現,試想如果人們將麋鹿胎兒分而食之或者作他用,必定會在別處發現相似的骨骼,然而事實上并非如此。從常識來說,這些個體發育不完全,肉量及骨髓少,無法提供人們想要獲取的肉食資源。可與H13對比的有丹麥中石器時代的采集狩獵遺址——林克洛斯特(Ringkloster)遺址,該遺址出土的動物骨骼中28.4%的馬鹿和18.6%的歐洲狍為新生兒或胎兒。但是后者因掌、跖骨遠端發現有一圈切割痕跡,作者推測這些幼鹿可能是為了剝皮而被宰殺的[22],這顯然與H13不同。
還有一個值得注意的問題,既然是直接將小鹿丟棄至灰坑中,那為什么在發掘現場沒有發現完整的骨架?這一點可以從骨骼狀態本身和埋藏學角度來分析:首先,這種骨壁薄、骨質松的胎兒骨骼是極為脆弱的,在肌肉、皮毛組織腐爛之后,其很難在漫長的埋藏歲月中保持原本的狀態;其次,根據對H13內容物的描述,這些骨骼發現于灰坑開口21厘米以下,說明在人們集中丟棄了一批麋鹿胎兒之后,這個灰坑可能長期暴露在外,受到了外界的擾動。
民族學材料表明每年農歷的二、三月份即所謂的“鹿胎期”是母鹿懷孕的季節,最適合獵鹿[23]。獵獲的鹿胎自母體取出后,一般并不食用,多被直接丟棄。
綜上,筆者推測H13是一處專門用來丟棄麋鹿胎兒的灰坑,在人們將獵殺的懷孕麋鹿帶回營地之后,處理過程中將其腹中的胎兒直接丟進了H13,并未對其進一步地利用。
(四)麋鹿各部位出現頻率的探討
麋鹿作為遺址中出現頻率最高的動物,是人們的主要肉食來源,其各部位的出現頻率可以在一定程度上幫我們還原先民的行為。本文采用MNE(Minimum numbers of elements,即某種動物用于鑒定分類時的解剖學部位的最小數量)和MAU( Minimum animal unit,其用于 MNE 頻率之間的對比)兩個概念來對解剖學部位的出現頻率進行量化分析。賓福德認為對于采集狩獵遺址來說,人們會對動物肢解和搬運,動物某一個部位的出現并不代表其整個個體在遺址中的出現,因此MNI存在著很大的誤差,而相較于MNI,MNE和MAU可以更清晰地表現出某個部位在遺址中出現的頻率,而不受性別、年齡的影響[24]。
對開莊遺址出土的麋鹿骨骼的MNE、MAU和%MAU[25]進行統計(表二),各部位出現頻率(%MAU)見圖九。
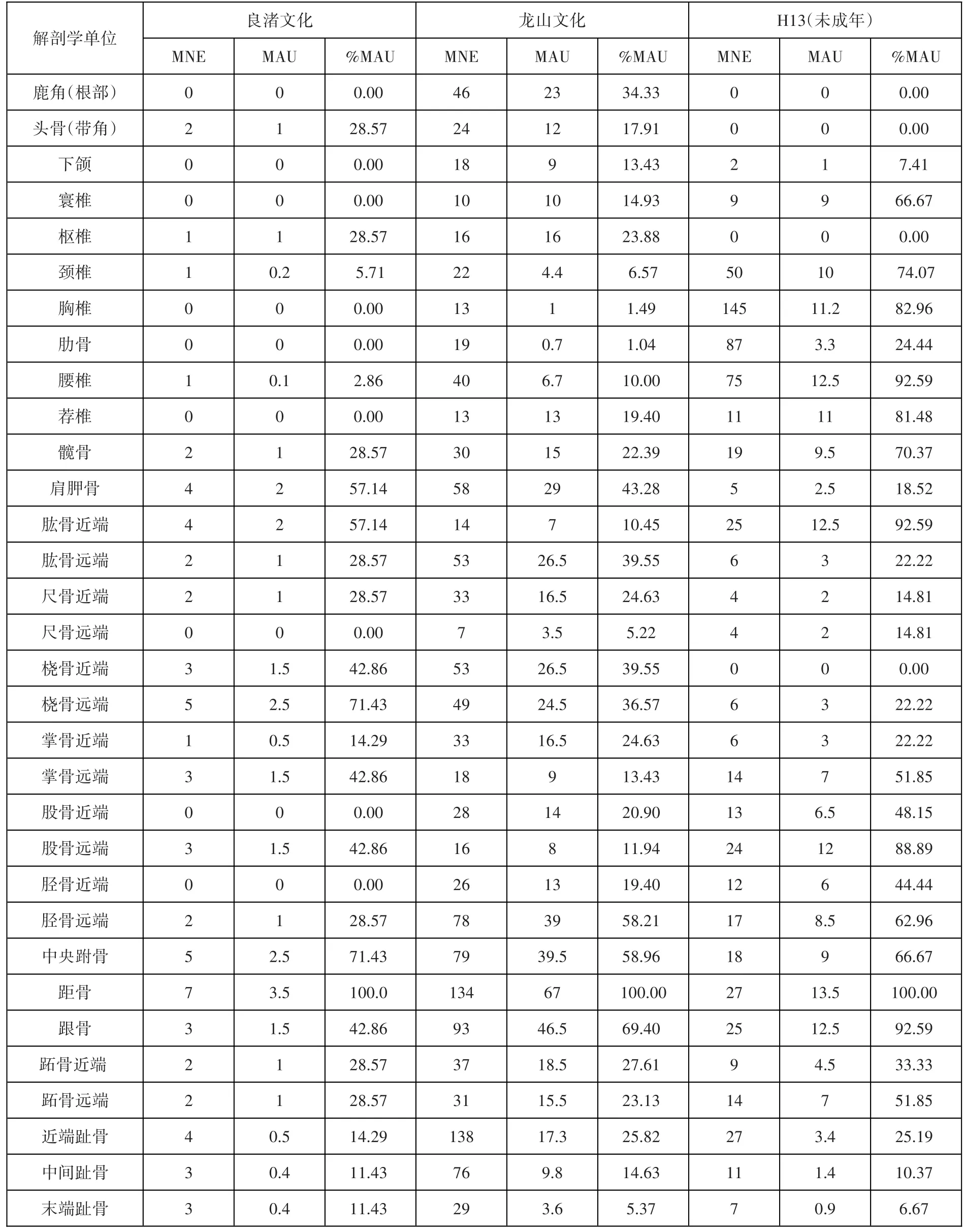
表二// 開莊遺址麋鹿解剖學單位MNE、MAU及%MAU統計表(單位:個)
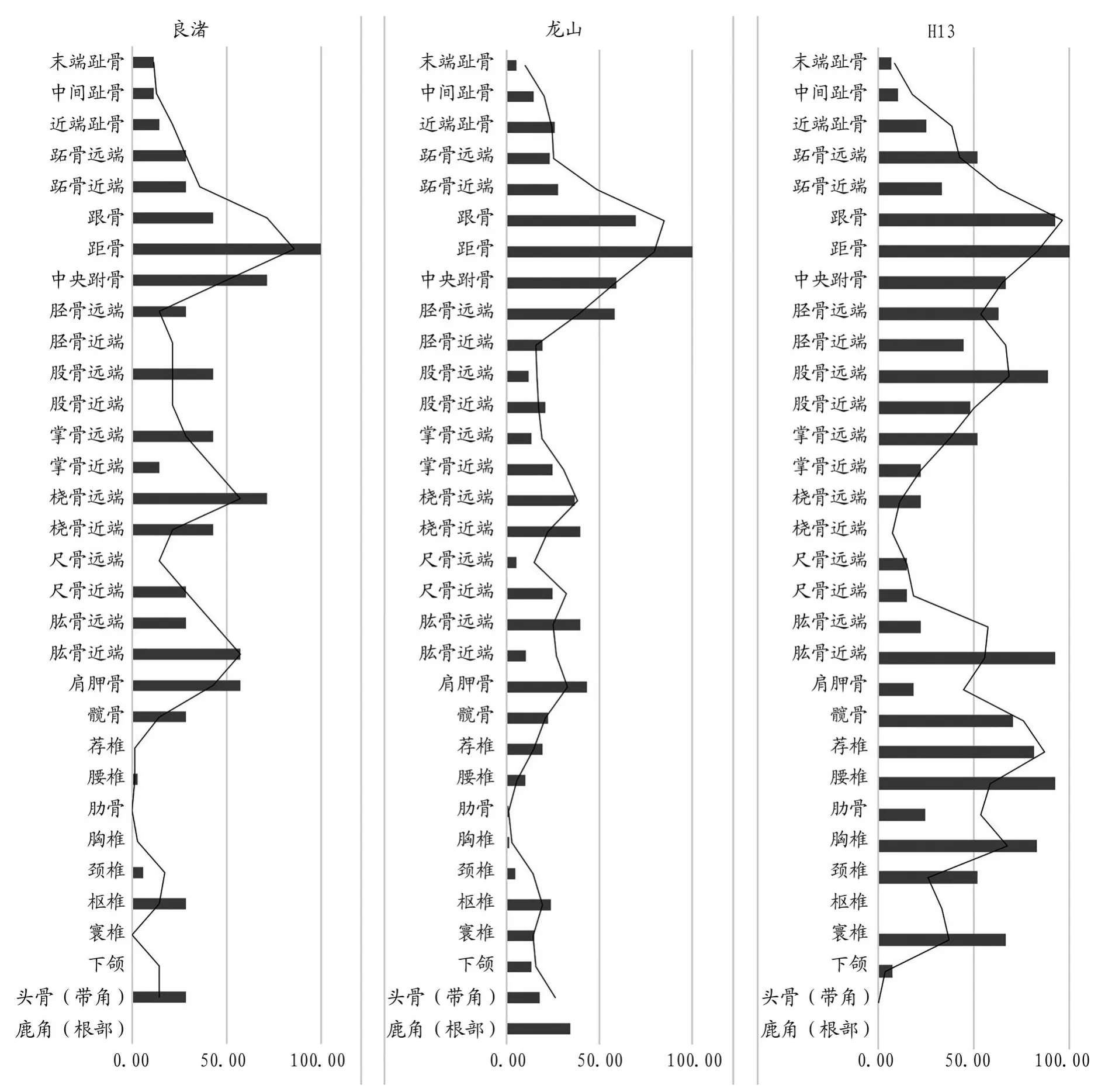
圖九// 開莊遺址良渚文化、龍山文化、H13(胎兒)麋鹿各部位頻率圖
由于H13內的胎兒骨骼很可能均為完整個體,因此將H13內的胎兒標本單獨進行了計算,一定程度上可以將其作為一個參照標準,但同時也有較大的弊端:H13中的麋鹿個體為骨骼未發育完全的新生兒或胎兒,鑒定過程較為困難,很多具有鑒定特征的部位缺失、未愈合或者是未成形,因此可能會造成某些解剖部位的數據偏差及缺失,如圖九H13中的鹿角、頭骨和橈骨近端。除去因材料本身限制導致的鑒定結果誤差,H13中其余部位的頻率分布還是比較均衡的,中軸骨(寰椎、樞椎、頸椎、胸椎、肋骨、腰椎、薦椎、髖骨)、前肢(肩胛骨、肱骨、尺骨、橈骨、掌骨)、后肢(股骨、脛骨、中央跗骨、根骨、距骨、跖骨)以及末端骨骼(趾骨)的豐度都比較高。
由于良渚文化晚期的標本量較小,所以會有部分部位缺失的情況,但良渚文化晚期和龍山文化時期的部位頻率分布與走勢較為相似,均整體呈現四肢骨多,中軸骨少的規律。具體表現為:后肢關節部(中央跗骨、根骨、距骨)最高,這與3個骨骼部位的形狀有關,其不易因受到外力而破碎,更容易在遺址中保存下來,也更容易被辨認;前肢、后肢和頭部(包括頭骨和角)的頻率分布較為平均;而中軸骨出現頻率較低(圖一〇[26])。成年個體與H13中的未成年個體所表現出的部位頻率是明顯不同的。
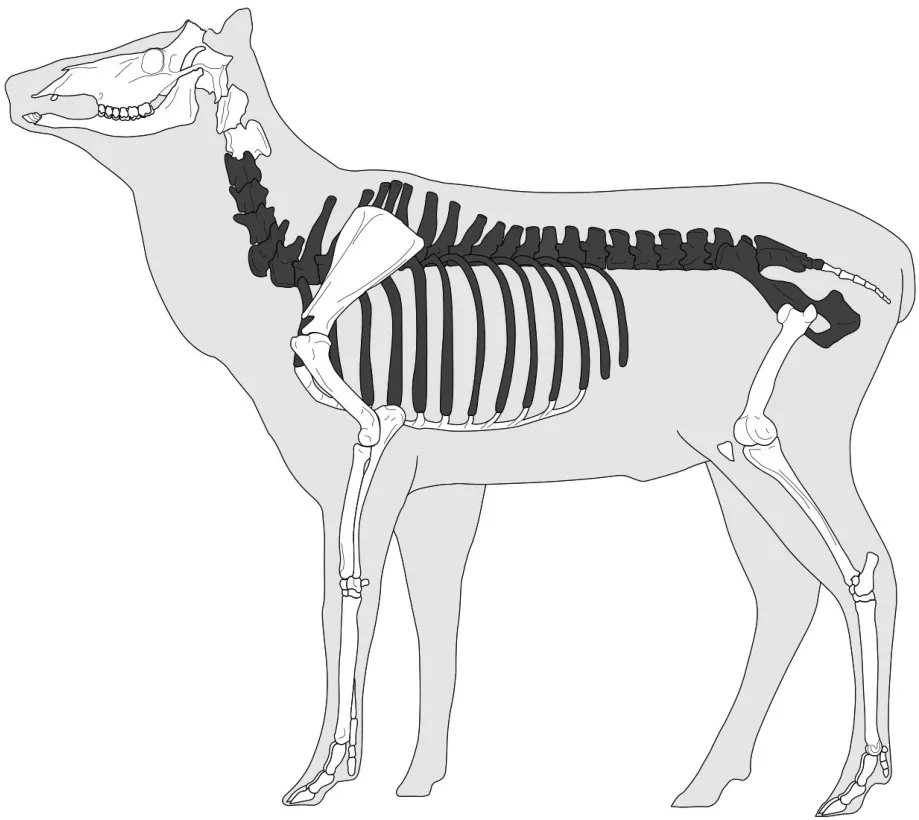
圖一〇// 開莊遺址成年麋鹿相對缺失(出現頻率低)部位示意圖(黑色部分)
在對比成年和未成年個體的部位頻率差異之前,先要解釋兩個現象的存在:第一,良渚文化和龍山文化兩個時期的成年麋鹿的下頜及牙齒數量都不多,而相較于馬家浜遺址[27]等出土了大量麋鹿牙齒、下頜標本的史前遺址來說,開莊遺址的麋鹿牙齒及下頜出現頻率是非常低的;第二,開莊遺址發現的26件(良渚文化晚期2件、龍山文化時期24件)保存相對完整的頂骨都帶角(包含角柄、角環和角),且大多發現有加工痕跡。由于雄性麋鹿長角而雌性不長角,因此這些標本都屬于雄性個體。開莊并未發現屬于雌性個體的不帶角的頂骨標本。出現上述牙齒下頜部頻率低和僅發現有雄性頭骨的兩個現象的原因,可能是由于在狩獵過程中,人們為了減少路途上的負擔,而直接將沉重的頭骨及下頜部位丟棄,很少將其帶回營地,但為了提供制作角器的原材料,帶角的雄性頭骨會被保留。
回到上一個問題,用以食用的成年個體與未被利用的未成年個體之間的部位頻率的差異反映了人們對成年麋鹿的特殊處置。那是什么行為造成這樣的現象呢?筆者認為有兩種可能:第一,營地間的搬運行為。獵人們將獵來的麋鹿帶回營地,并對其進行簡單肢解,從肩胛骨部剖解下前肢,從股骨部剖解下后肢,四肢留在此處進行食用,而將中軸骨部尤其是胸椎腰椎帶肉部位搬運到其他營地,由此推測開莊遺址可能是一個獲取營地或是一個對獵物進行前處理的場所。第二,此處為骨、角器取料加工場所。開莊遺址出土了骨器、角器、牙器,以及制作骨、角器的原料、坯料、廢料等共計291件[28]。其中很明顯有利用麋鹿長骨,尤其是掌、跖骨制作骨鏃、骨鑿等工具的現象,也出現大量麋鹿角上殘留有劈砍、切鋸等取料和加工的痕跡,由此推斷開莊遺址可能是一處對骨、角器前處理或加工的場所,因此會保留有更多的四肢骨。
上述兩種可能并不對立,開莊遺址可能既是一處獲取營地,也是一處骨、角器加工地。人們于臨近沼澤處狩獵麋鹿,后將其帶回營地進行初步肢解,四肢部留在原地直接食用,之后再對其進行簡單處理,加工制成工具,而一部分人將麋鹿中軸帶肉部搬運到其他的營地或聚居地。
五、結語
長江中下游地區河網密布,自然資源豐富,孕育了眾多史前文化。相較于寧紹平原、太湖平原,江淮東部地區開展過系統的動植物研究的遺址還相對較少,開莊遺址的發掘以及后續進行的分支研究,為重建該地區史前社會提供了寶貴的資料。
開莊遺址2018—2019年度的發掘中,動物遺存包含龜科、大雁、狗、熊、豬、梅花鹿、水鹿、麂、獐、麋鹿、水牛,數量以野生哺乳動物為主,尤以麋鹿為甚。良渚文化晚期是以飼養家豬和狩獵野生動物相結合的生業經濟模式,而龍山文化時期的肉食來源以狩獵野生鹿類動物為主。通過對H13性質以及麋鹿各部位出現頻率的討論,開莊遺址可能為一處獲取營地,是作為獵人們對獵物進行前處理以及制作骨、角器的場所,各營地間存在搬運行為。
值得注意的一點是,開莊遺址的兩個階段——良渚文化晚期和龍山文化時期,處于新石器時代晚期甚至末期,當時均已經出現了農業種植和家畜飼養的技術。距今5300—4300年的良渚文化時期,農業經濟達到全盛,形成了以水稻生產與家豬飼養相結合的生業經濟模式[29],而龍山文化時期的眾多遺址也多形成了以食用家豬為主的肉食結構[30]。在這種大趨勢下,開莊先民反其道行之,選擇了狩獵(或者是半家養半狩獵)的肉食獲取方式,動物構成以野生大型鹿為主。這個情況在江淮下游地區并不是個例,良渚文化時期的上海廣富林遺址[31]、江蘇蔣莊遺址[32],馬橋文化時期的上海馬橋遺址[33]、江蘇綽墩遺址[34]的動物遺存也均顯示出了以野生動物為主的現象。這些遺址所顯示出的共同現象,為我們今后研究良渚文化與后良渚時期的社會經濟與社會進程,以及該時期南北方文化的交流融合提供了思路和資料。
