Mechlppdk基因在木薯中的表達分析及RNA干擾載體構建和遺傳轉化
2021-07-09 09:06:30王海燕周新成王文泉
中國農學通報 2021年17期
王海燕,陳 新,周新成,沈 旭,孔 華,王文泉
(1中國熱帶農業科學院熱帶生物技術研究所,海口 571101;2南京農業大學生命科學院,南京 210095;3海南大學熱帶作物學院,海口 570228)
0 引言
木薯是具有高生物量的重要熱帶植物,起源于南美洲,是全球三大塊根作物之一,第六大糧食作物和新興能源作物,具有高效累積光合產物和抗旱、耐瘠薄等特性[1],廣泛種植于全球熱帶地區和中國兩廣,海南地區。栽培木薯是典型的C3-C4型熱帶植物,具有高光效高淀粉累積的特點。木薯塊根中積累貯藏的同化物僅僅是地上合成同化物的1/5,所以木薯葉片具有超強的光合碳固定能力。但是C4型碳固定對木薯葉片的這種超強光合碳固定的作用效率尚未見報道。
栽培木薯葉片PEPC酶的活性從8.3~80 mmol/(kg(Chl)·s),約是玉米和高粱的 15%~25%[2]。栽培木薯PEPCase活性遠遠高于田間大豆等C3植物的活性,與C3-C4中間型黃花菊屬幾個種的活性相當,比包含類似花環結構、C3-C4型黍屬的Panicum milioides中的活性高2~3倍[3],C4PEPC的存在通過免疫學方法得到進一步確證。利用來自玉米的PEPC、蘋果酸酶和蘋果酸脫氫酶的探針均能在栽培木薯葉片中檢測到蘋果酸酶和蘋果酸脫氫酶的存在。當前對栽培木薯光合作用屬于C3-C4中間型的認識僅限于栽培木薯葉片中有C4酶的存在,但是其中C4光合碳同化的效率,及對木薯生長發育的影響迄今尚未有報道。
丙酮酸磷酸雙激酶(PPDK)是C4光合作用的關鍵酶,催化C4光合作用原初CO2固定的受體磷酸烯醇式丙酮酸(PEP)的再生,繼而由磷酸烯醇式丙酮酸羧化酶(PEPC)催化PEP接受二氧化碳,合成草酰乙酸(OAA),至此完成二氧化碳的固定。玉米中C4循環是由葉肉細胞和維管束鞘細胞協作完成。同時發現玉米PPDK是C4循環的限速酶,一旦PPDK的活性被干擾,C4途徑就會被影響[4]。PPDK首先是在C4植物中發現[5],后面相繼在C3植物,原生動物和細菌都有發現。PPDK在C3和C4植物中是以同型四聚體形式存在,在細菌和原生動物是以二聚體形式存在[6],但他們的初級結構非常相似,表明ppdk基因在原核生物和真核生物分化前就存在了。迄今,科學家們已在多種植物中克隆到 ppdk基因,如小麥[7]、玉米[8]、水稻[9]、擬南芥[10]、甘蔗[11]、高粱[12]、籽粒莧[13]、家稗[14]等。
RNA干擾(RNA interference,RNAi)是指細胞中導入與內源性mRNA編碼區某段序列同源的雙鏈RNA(double-stranded RNA,dsRNA),可致該mRNA發生特異性降解從而導致基因表達沉默的現象[15]。該技術能夠干擾特定基因,高效、特異地阻斷體內基因表達,誘使細胞表現出特定基因缺失的表型,具有高效性、擴大性、特異性及可遺傳性等特點。經過多年實踐,RNA干擾技術日趨成熟,現已廣泛應用于植物基因功能研究[16-17]。本研究擬利用QPCR確定Mechlppdk在木薯不同組織器官的表達譜,構建木薯Mechlppdk的RNA干擾載體,并轉化木薯,獲得Mechlppdk的干擾突變體,為下一步木薯丙酮酸磷酸雙激酶基因Mechlppdk功能的研究提供研究材料。
1 材料與方法
1.1 實驗材料和試劑
ppdk基因表達譜分析木薯材料葉片,莖稈,塊根均采自生物所試驗地田間種植180天木薯植株(Mahnihot esculenta var.Arg7)。木薯遺傳轉化用材料是模式木薯TMS60444。大腸桿菌(Escherichia coli)DH5α系本實驗室留存,農桿菌(Agrobacterium tumefaciens)LBA4404是上海生命科學研究院張鵬研究員饋贈,RNA干擾中間載體pHANNIBAL和pART27表達載體是本研究所郭安平課題組饋贈;Taq DNA polymerase、pMD18-T、T4 DNA連接酶購自大連寶生物公司,DNA凝膠回收試劑盒購自OMEGA公司,其他生化試劑為進口或國產分析純試劑。引物合成和DNA測序是由生工生物工程(上海)有限公司完成。
1.2 方法
1.2.1 木薯葉片RNA提取和cDNA的合成 參照李正緒方法進行[18]。
1.2.2 Mechlppdk QPCR引物設計 通過比對GeneBank上公布的木薯Mechlppdk的cDNA序列,在轉運肽區設計定量引物,如下QF:5’-GCAGGGGTATAGAGGAT CGC-3’;QR:5’-GGCGCCGTCCTAATTAACAT-3’,木薯的β-actin基因作為管家基因,actinF:5’-CAAGGGCAACATATGCAAGC-3’,actinR:5’-CCTT CGTCTGGACCTTGCTG-3’。熱循環儀是Rotor-gene 6000,擴增程序如下:95℃預變性 1 min;95℃ 10 s,59℃ 15 s,72℃ 20 s,40個循環。每個樣品設置3個重復,取其平均值用于表達分析。采用2-ΔΔCT進行數據分析,并使用IBM SPSS Statistics 19.0進行差異顯著性分析。
1.2.3 Mechlppdk RNAi的引物設計 通過找到Mechlppdk的特異區段,分別設計2對引物,即在引物序列前分別加兩個不同的酶切位點,形成帶有不同酶切位點的兩個反向互補片段,具體引物序列如下:senseF:5’-GAATTCATGTCGTCGGCG-3’;senseR:5’-AGGAGCAAGAGACTCGAG-3’;antisenseF:5’-ATC GATTCTCTTGCTCCT-3’;antisenseR:5’-CGCCG ACGACATTCTAGA-3’。同時把中間載體的內含子序列加到反向互補片段之間,預測其二級結構。
1.2.4 木薯Mechlppdk RNAi片段的PCR擴增及RNA干擾中間載體的構建 以1.2.2中合成的cDNA為模板,利用上述設計的引物,擴增Mechlppdk的特異片段,電泳,切膠、回收后連接PMD18-T載體,轉化感受態細胞(E.coli DH5α)后,提取質粒并測序。根據測序結果選取與所設計干擾片段序列一致的兩個質粒分別命名為p Mechlppdki-XE和p Mechlppdki-CX,分別用Xho I/EcoR I和Cla I/Xba I進行雙酶切,電泳,切膠回收目的片段分別命名為Mechlppdki-XE和Mechlppdki-CX。同時用Xho I/EcoR I雙酶切中間載體p HANNIBAL,膠回收大片段后用T4DNA連接酶將其與回收的Mechlppdki-EX連接,轉化感受態細胞(E.coli DH5α)后用氨芐(終濃度為50 μg/L)篩選陽性克隆,提取質粒并酶切檢測Mechlppdki-XE是否與pHANNIBAL載體連接,連接上Mechlppdki-XE的pHANNIBAL質粒命名為pHANNIBAL-MechlppdkiXE,將pHANNIBALMechlppdkiXE進行Cla I/Xba I的雙酶切,切膠,回收目的片段后用T4DNA連接酶將其與回收的Mechlppdki-CX連接,轉化感受態細胞(E.coli DH5α)后用氨芐(終濃度為50 μg/L)篩選陽性克隆,提取質粒并酶切鑒定,獲得陽性克隆,命名為pHANNIBAL-MechlppdkiXECX。
1.2.5 RNA干擾表達載體的構建 根據實驗室干擾載體pART27的質粒圖譜,使用Not I對pHANNIBALMechlppdkiXE-CX和pART27分別進行Not I酶切,電泳,切膠回收,將回收后的MechlppdkiXE-CX大片段和pART27的大片段進行連接,轉化感受態細胞(E.coli DH5α)后用卡那霉素(終濃度為50 μg/L)篩選陽性克隆Mechlppdk RNAi表達載體,具體流程詳見圖1。

圖1 Mechlppdk RNAi干擾載體構建流程圖
1.2.6 RNA干擾載體轉化木薯及nptII的PCR驗證及QPCR檢測 將1.2.4中獲得的干擾載體通過電擊轉化的方法轉化農桿菌LBA4404,獲得含有干擾載體的農桿菌菌株,然后侵染木薯TMS60444脆性胚愈傷,通過共培養,洗菌,誘導,最后獲得具有卡那霉素抗性的轉基因木薯苗。具體方案參照文獻[19-20]。將生根篩選出的轉基因木薯苗提取基因組DNA,通過PCR的方法進行nptII的鑒定。利用Mechlppdk QPCR引物對獲得的轉基因株系進行Mechlppdk的表達分析。
2 結果與分析
2.1 Mechlppdk在木薯不同組織器官的表達分析
如圖2,通過對180天的木薯根、莖和葉3個器官中Mechlppdk基因的表達分析,發現Mechlppdk在葉片中表達量最高,達到內參基因的50%,莖中只有可見的少量表達,根中沒有表達。說明Mechlppdk主要在葉片中起作用。
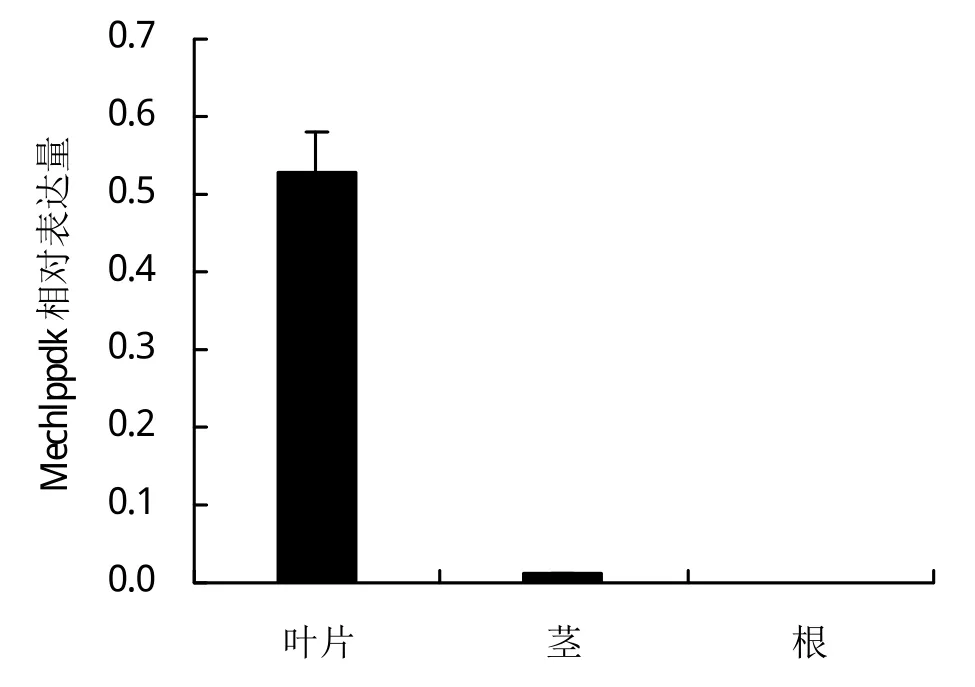
圖2 Mechlppdk在根、莖、葉中的相對表達量
2.2 Mechlppdk干擾載體特異序列的選擇
將JGI網站公布的Mechlppdk(Manes.03G188300),經BLAST,選擇轉運肽區一段保守序列,自第1個堿基至165個堿基,共165個堿基。具體序列信息如圖3A。將正反序列中間連接pHANNIBAL的內含子,預測二級結構,如圖3B,形成典型的發夾結構。
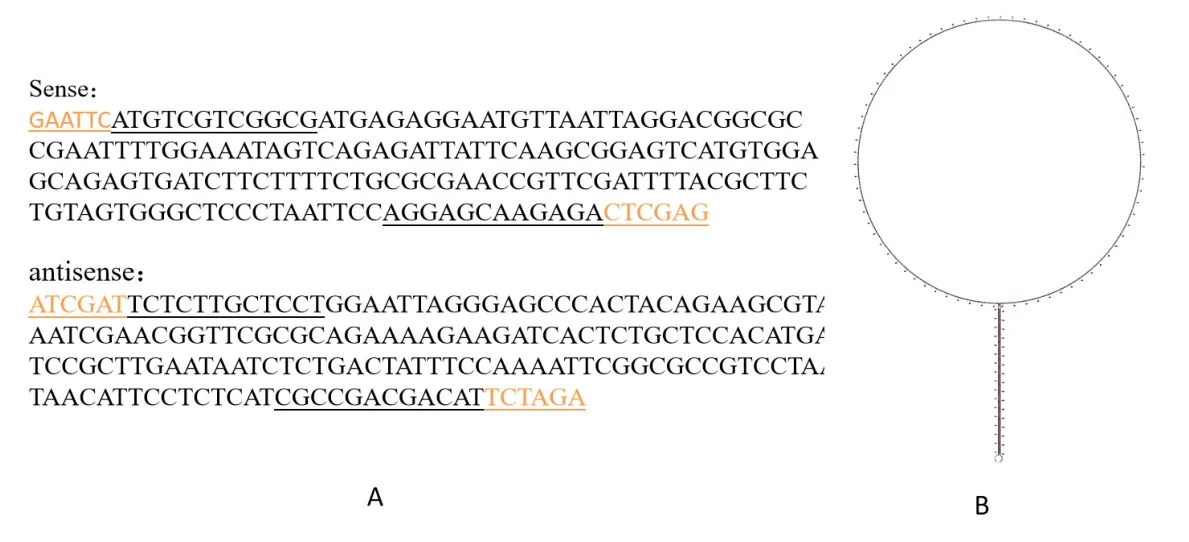
圖3 木薯Mechlppdk siRNA設計
2.3 Mechlppdk RNA干擾片段的克隆
通過RT-PCR擴增得到175 bp左右的目的片段(包含酶切位點序列),見圖4A,將目的片段連接至pMD-18T克隆載體上,通過化學轉化法轉至大腸桿菌DH5α中,經氨芐篩選獲得陽性菌落,通過PCR鑒定,結果顯示得到的小片段與預期大小一致(圖4 B)。將陽性克隆送測序,選擇與Mechlppdk選定序列完全一致的克隆,提取質粒,分別命名為pMechlppdkiXE和pMechlppdkiCX,用于下步構建。
2.4 Mechlppdk RNA干擾中間載體的構建
首先用EcoR I/Xho I分別酶切pMechlppdkiXE和pHANNIBAL,電泳,分別膠回收小,大片段,并用T4DNA連接酶將目的片段和載體連接,轉化大腸桿菌DH5α,在帶有氨芐的培養基上篩選陽性克隆,提取質粒酶切鑒定,結果見圖5A,經EcoR I/Xho I雙酶切后,獲得與圖4中PCR擴增大小一致的條帶,將該陽性質粒命名為pHANNIBAL-MechlPPDKiXE。繼而用Cla I/XbaI雙酶切pHANNIBAL-MechlppdkiXE和pMechlppdkiCX,電泳,膠回收目的片段,用T4DNA連接酶連接載體和小片段,轉化大腸桿菌DH5α,在篩選培養基上篩選陽性克隆,PCR擴增正確的克隆進行質粒提取和酶切鑒定,結果見圖5B,經Cla I/Xba I雙酶切,在100 bp和250 bp的條帶之間有一條約170 bp的條帶,與圖4中PCR擴增的條帶大小一致,酶切正確的陽性質粒命名為pHANNIBAL-MechlppdkiXE-CX.

圖4 Mechlppdk RNA干擾片段的擴增及克隆鑒定

圖5 pHANNIBAL-MechlppdkiXE-CX中間載體酶切鑒定
2.5 Mechlppdk RNA干擾表達載體的構建并轉化農桿菌LBA4404
用Not I分別酶切pART27和pHANNIBALMechlppdkiXE-CX,電泳,膠回收目的片段和線性載體,用T4DNA連接酶將載體和目的片段連接,轉化大腸桿菌DH5α,在帶有卡那霉素的LB培養基上篩選陽性克隆,經菌液PCR(圖6A),將菌液PCR陽性的克隆進一步提取質粒,Not I酶切,有8個克隆可切出與預期大小一致條帶,結果見圖6B,此時即獲得pART27-MechlppdkRNAi。將 pART27-MechlppdkRNAi轉化農桿菌LBA4404的感受態細胞,篩選培養基上獲得陽性克隆,并經PCR和質粒酶切檢測為陽性。
圖6 pART27-MechlppdkRNAi的分子鑒定
2.6 pART27-MechlppdkRNAi轉基因木薯苗的獲得
將獲得的帶有干擾載體的LBA4404侵染木薯品種TMS60444的脆性胚愈傷,經過誘導,篩選,獲得具有卡那霉素抗性苗。轉基因結果如圖7。抗性苗放置在添加有50 μg/L卡那霉素的MS培養基上進行生根篩選,如圖7D,7~9天部分轉基因植株在抗性培養基上生根,野生型沒有生根。繼而選擇生根的株系進行卡那霉素Npt II的PCR鑒定,成功擴增到目的條帶,如圖7E。最終獲得7個Mechlppdk的干擾株系。

圖7 轉pART27-MechlppdkRNAi木薯苗的誘導及鑒定
對上述獲得的7個轉基因木薯株系考察Mechlppdk基因的表達情況,結果如圖8。7個株系中有4個株系(1,2,3,7)的Mechlppdk的表達量大幅下調,其中最大的有7號株系,Mechlppdk的表達量下調到野生型的9.7%。有2個株系的Mechlppdk的表達量有輕微下調,表達量分別下調到野生型的87%和74%。
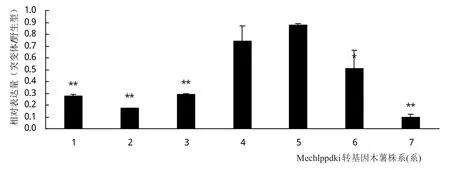
圖8 Mechlppdk干擾株系Mechlppdk的相對表達分析
3 討論與結論
RNAi是由雙鏈RNA分子在細胞內干擾同源基因表達下調或沉默的技術[21]。RNA干擾最早追溯到1990年,Napoli等[22]在將查耳酮合酶基因(chs)轉入牽牛花中意外發現的。RNAi能夠定向關閉生物體內的某一基因,使其不發揮作用,同時不影響其他基因,方便人們對特定基因功能的研究[23]。當前廣為應用的還有crisp技術,是在DNA水平敲除目的基因,效率高,但對于重要的功能基因,完全敲除后,往往具有植株致死性。而RNAi技術可以不同程度的干擾目的基因的表達,對于研究重要功能基因具有優勢。但是RNAi的特異性是相對的,在某些情況下,siRNA能在翻譯水平抑制與之不完全相關的基因表達即脫靶效應。為了應對此類脫靶現象,應該針對同一基因多設計幾個靶點,以確保靶基因表達的有效抑制[24]。本研究共獲得7個Mehlppdk的干擾株系,其中3個株系Mechlppdk的表達量顯著下調,說明本次對Mechlppdk干擾操作有效。
雙子葉植物中,ppdk由單基因編碼[25],5’有兩個不同的啟動子分別產生細胞質和質體定位的轉錄本[26]。木薯ppdk同樣有2個啟動子,分別產生葉綠體定位的Mechlppdk和細胞質定位的Mecyppdk。前期研究發現Mecyppdk完全嵌套在Mechlppdk里面,所不同的是Mechlppdk的5’多一轉運肽序列,具有組織特異性,主要在葉綠體表達[27]。本研究為更準確的干擾Mechlppdk基因,選擇了該基因的5’轉運肽區進行干擾載體的構建,最大限度的減少對Mecyppdk基因的干擾。目前尚未看到關于木薯Mechlppdk基因功能的研究報道。Zhang等[28]獲得了chlppdk功能缺失的玉米突變體,該材料的葉片在種植108 h后出現白化,且消耗完種子內存活的營養物質就會死亡。表明chlPPDK對于玉米的生存至關重要。接下來將利用獲得的干擾突變體展開生理生化和分子生物學研究,以期闡明chlPPDK對木薯生長發育的影響。
本研究通過QPCR發現Mechlppdk在木薯葉片中具有較高的表達,達到內參基因的50%;通過構建木薯Mechlppdk的干擾載體轉化木薯,獲得7個陽性轉基因株系,其中3個株系較大幅度的降低了Mechlppdk的表達,最低的僅是本底chlppdk的9%。這些轉基因干擾株系將為Mechlppdk功能研究提供非常有利的實驗材料,同時能夠部分闡述光合C4固定對木薯生長發育的影響。
