VHL抑制劑對魚藤酮所致帕金森病秀麗隱桿線蟲模型的影響
2021-07-08 10:24:24高向東
中國藥科大學學報 2021年3期
關鍵詞:模型
查 倩,高向東,陳 松
(中國藥科大學生命科學與技術學院江蘇省生物藥物成藥性研究重點實驗室,南京211198)
帕金森病(Parkinson′s disease,PD)是一種與年齡相關的神經退行性疾病,目前全球有600多萬患者,隨著世界人口老齡化加劇,PD患者數量將逐年增加[1]。PD與線粒體質量控制途徑的異常調節密切相關,線粒體未折疊蛋白反應(mitochondrial unfolded protein response,mito-UPR)作為線粒體質量控制機制之一,激活的mito-UPR途徑能夠促進細胞存活和受損的細胞器恢復,但mito-UPR的長時間過度激活會導致缺陷的線粒體異常積累,從而破壞線粒體穩態平衡[2-3]。秀麗隱桿線蟲作為神經生物學研究的理想模式生物,其神經網絡簡單,僅有302個神經元,共4對多巴胺能神經元,其中頭部有2對CEPs(cephalic neurons)和1對ADEs(anterior deirid neurons),尾部有1對PDEs(posterior deirid neurons)。由于線蟲身體透明,熒光蛋白可以在活體線蟲中可視化,可直接觀察神經元中目的蛋白的表達情況[4]。同時,線蟲基因組是第一個被完全測序的多細胞生物的基因組,據統計,60%~80%的人類致病基因在線蟲基因組中具有直系同源物[5-7]。目前,已建立的線蟲帕金森病模型包括表達α-突觸核蛋白(α-synuclein,α-syn)或富含亮氨酸的重復激酶2(leucine-rich repeat kinase 2,LRRK2)的轉基因線蟲,以及PRKN/pdr-1,PINK1/pink-1,DJ-1/djr-1.1/djr-1.2和ATP13A2/catp-6缺失的線蟲模型等[8-9]。此外,有研究表明,神經毒素如魚藤酮、1-甲基-4-苯基吡啶離子(MPP+)和6-羥基多巴胺(6-OHDA)可誘導線蟲線粒體DNA損傷,導致線蟲多巴胺能神經元退化[10-11]。線蟲PD模型均能表現出多種表型缺陷,包括多巴胺能神經元的丟失,多巴胺依賴性行為異常等[12]。
本課題組于前期研究中建立了轉基因線蟲zcIs9;otIs181,發現線粒體復合體Ⅰ亞基NDUF7突變導致ADE多巴胺能神經元特異性退化,出現PD樣癥狀,同時基于該線蟲模型,進一步研究發現線蟲VHL-1功能缺失能顯著緩解ADE多巴胺能神經元退化情況[13]。在此研究基礎上,本研究采用魚藤酮建立PD線蟲模型,分析VHL抑制劑對PD線蟲模型神經元退化及行為學異常的影響。
1 材料
1.1 試劑與溶液
魚藤酮(純度:98.12%)、VHL抑制劑VH298(純度:99.83%,美國MCE公司);膽固醇(上海麥克林生化科技有限公司);瓊脂粉、酵母粉、蛋白胨(美國Oxoid公司);檸檬酸鉀(上海阿拉丁生化科技股份有限公司);次氯酸鈉溶液(國藥集團化學試劑有限公司);其他試劑均為市售分析純。
NGM培養基、M9溶液、S培養基、LB液體培養基、線蟲裂解液等溶液按照文獻[14]進行配制。
1.2 儀器
高速冷凍離心機、低溫培養箱(美國Thermo公司);體視顯微鏡、熒光體視顯微鏡(日本Nikon公司);倒置熒光顯微鏡(日本Olympus公司);高壓滅菌鍋(日本Panasonic公司)。
1.3 動物
線蟲品系:轉基因線蟲zcIs9;otIs181(本課題組前期構建[13])。
1.4 菌株
尿嘧啶缺陷型OP50大腸埃希菌(福建上源生物科學技術有限公司)。
2 方法
2.1 線蟲培養
將線蟲接種于涂布有尿嘧啶缺陷型OP50大腸埃希菌的線蟲固體生長培養基(NGM培養基)平板上,于20℃低溫培養箱中培養[15]。
2.2 線蟲同步化處理
使用M9溶液1 mL將處于產卵期的線蟲從直徑為6 cm的NGM平板上沖洗下來,加至1.5 mL EP管中,2 000 r/min離心30 s,吸除上清液700μL,再加入線蟲裂解液700μL,混勻5 min左右,4 000 r/min離心30 s,吸上清液至管內剩余液體約100μL,再加入M9溶液900μL,輕吹混勻,重復清洗3次后,4 000 r/min離心30 s,吸除上清液900μL,輕吹混勻剩下的蟲卵溶液,吸至未接種OP50大腸埃希菌的NGM平板上,放置20℃低溫培養箱培養,次日即可得到L1期線蟲幼蟲。
2.3 魚藤酮暴露試驗及線蟲死亡和多巴胺能神經元退化情況分析
將同步化處理得到的L1期線蟲接種至涂布有OP50大腸埃希菌的NGM培養基上,放置低溫培養箱生長1 d后進行實驗。魚藤酮誘導PD線蟲模型試驗采用96孔板進行,每孔溶液終體積為100μL,設置魚藤酮最終濃度為8.0,4.0,2.0,1.0,0.5,0μmol/L共6組,每組4個孔,用S培養基重懸OP50大腸埃希菌,保證每孔加入10 mg/μL OP50大腸埃希菌5μL,之后加入含有20~30只幼蟲的S培養基10μL,次日觀察并記錄各組線蟲死亡情況,并且觀察各組生長至L4期線蟲的多巴胺能神經元退化情況。對于判斷線蟲是否死亡,在體視顯微鏡下觀察,當線蟲身體呈現僵直狀態且無運動行為時,用鉑絲針刺激其身體,無反應的線蟲則視為死亡。對于觀察線蟲多巴胺能神經元退化情況或mito-UPR情況,滴25 mmol/L左旋咪唑溶液5 μL于含有3%瓊脂糖墊的載玻片上,將處理后的線蟲放置溶液中進行麻醉,之后將蓋玻片輕輕壓住線蟲,于熒光體視顯微鏡或熒光倒置顯微鏡下觀察頭部ADE多巴胺能神經元退化情況或mito-UPR激活情況。對于判斷線蟲ADE多巴胺能神經元是否退化,在正常情況下,線蟲頭部ADE多巴胺能神經元mCherry紅色熒光斑點清晰可見,而當暴露于一定濃度的魚藤酮后,線蟲ADE多巴胺能神經元mCherry紅色熒光斑點缺失則視為ADE多巴胺能神經元退化。在正常情況下,線蟲蟲體GFP綠色熒光微弱,而當mito-UPR激活后,線蟲蟲體GFP綠色熒光明顯增強。
2.4 抑制劑VH298干預PD線蟲模型試驗
抑制劑干預試驗采用96孔板進行,每孔溶液終體積100μL,每孔20~30只L1期幼蟲,設置抑制劑VH298終濃度為400,200,100μmol/L進行提前干預,每組4個孔,1 d后使用M9溶液清洗線蟲3次,再將線蟲加入新鮮配制的含有魚藤酮和不同濃度VH298的96孔板中進行共孵育,1 d后使用M9溶液清洗線蟲3次,將線蟲轉移至NGM培養基上進行各項指標分析。觀察線蟲死亡和多巴胺能神經元退化情況的方法,參見“2.3”項。
2.5 線蟲運動行為指標的測定[16-17]
2.5.1 頭部擺動頻率分析 抑制劑VH298干預PD線蟲模型參見“2.4”項。在未接種OP50大腸埃希菌的NGM培養基上滴加M9溶液80μL,用鉑絲針挑取處理后的線蟲置于M9溶液中,待其恢復1 min后,在體視顯微鏡下觀察并記錄線蟲在1 min內頭部擺動的次數(線蟲頭部從一側擺向另一側又擺回來的次數)。
2.5.2 身體彎曲頻率分析 抑制劑VH298干預PD線蟲模型見“2.4”項。用鉑絲針挑取處理后的線蟲于未接種OP50大腸埃希菌的NGM培養基上,待其恢復1 min后,在體視顯微鏡下觀察并記錄其在30 s內身體彎曲的次數(線蟲在爬行過程中沿著身體長軸方向上1個波長變化作為1次身體彎曲)。
2.6 線蟲覓食行為的分析[16]
抑制劑VH298干預PD線蟲模型見“2.4”項。在直徑為9 cm的未接種OP50大腸埃希菌的NGM培養基中央滴大腸埃希菌OP50菌液5μL,形成直徑為1 cm的圓形菌苔,將線蟲放入距離培養皿中心位置4 cm處,每個培養皿放置20條。分別記錄4,12,24 h后接觸菌落的線蟲數,計算接觸菌落線蟲占線蟲總數的比例。
2.7 統計分析
實驗數據采用GraphPad Prism 8.0統計軟件進行分析,各項指標以xˉ±s表示,采用Two-Way ANOVA檢驗對線蟲覓食行為數據進行分析,采用One-Way ANOVA檢驗對其他數據進行分析,P<0.05時表明差異有統計學意義。
3 結 果
3.1 不同濃度魚藤酮導致轉基因線蟲zcIs9;otIs181死亡率變化
本研究選用帶有zcIs9和otIs181兩個遺傳標記的轉基因線蟲,其中zcIs9[hsp-60p∷GFP]用于監測線蟲mito-UPR,otIs181[dat-1p∷mCherry+ttx-3p∷mCherry]用于標記4對多巴胺能神經元和1對非多巴胺能神經元AIY。轉基因線蟲zcIs9;otIs181同步化處理后生長2 d,采用不同濃度的魚藤酮損傷線蟲,次日在體視顯微鏡下觀察并記錄各組線蟲死亡情況。結果如圖1所示,低劑量魚藤酮處理線蟲1 d即可導致線蟲死亡,當魚藤酮濃度上升至4.0和8.0μmol/L時,線蟲死亡率分別為49.24%和88.14%。以上結果說明,魚藤酮能導致轉基因線蟲zcIs9;otIs181死亡,并且呈現一定的濃度依賴性。
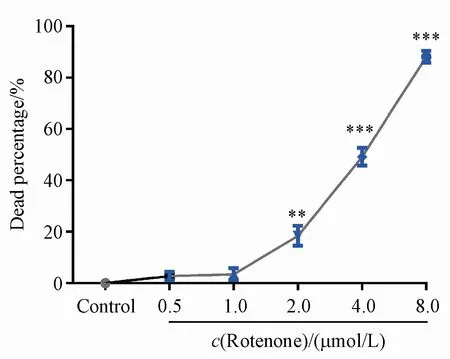
Figure 1 Evaluation of death in C.elegans strain with zcIs9;otIs181 with exposure to different concentrations of rotenone(xˉ±s,n=20-30)
3.2 不同濃度魚藤酮導致轉基因線蟲zcIs9;otIs181多巴胺能神經元退化情況
轉基因線蟲zcIs9;otIs181同步化處理后生長2 d,采用不同濃度的魚藤酮損傷線蟲,1 d后在熒光體視顯微鏡下觀察并記錄各組L4期線蟲多巴胺能神經元退化和mito-UPR激活情況。結果如圖2所示,與對照組相比,0.5,1.0和2.0μmol/L的魚藤酮損傷組線蟲ADE退化率分別為27.87%,29.17%和39.71%,魚藤酮濃度上升至4.0和8.0 μmol/L時,線蟲ADE退化率分別為84.17%和93.16%。以上結果說明,魚藤酮能導致轉基因線蟲zcIs9;otIs181的多巴胺能神經元退化,且與對照組相比,魚藤酮損傷組線蟲mito-UPR均被激活,說明魚藤酮可能通過誘導轉基因線蟲zcIs9;otIs181線粒體損傷,破壞線粒體穩態,進而導致多巴胺能神經元退化及線蟲死亡。
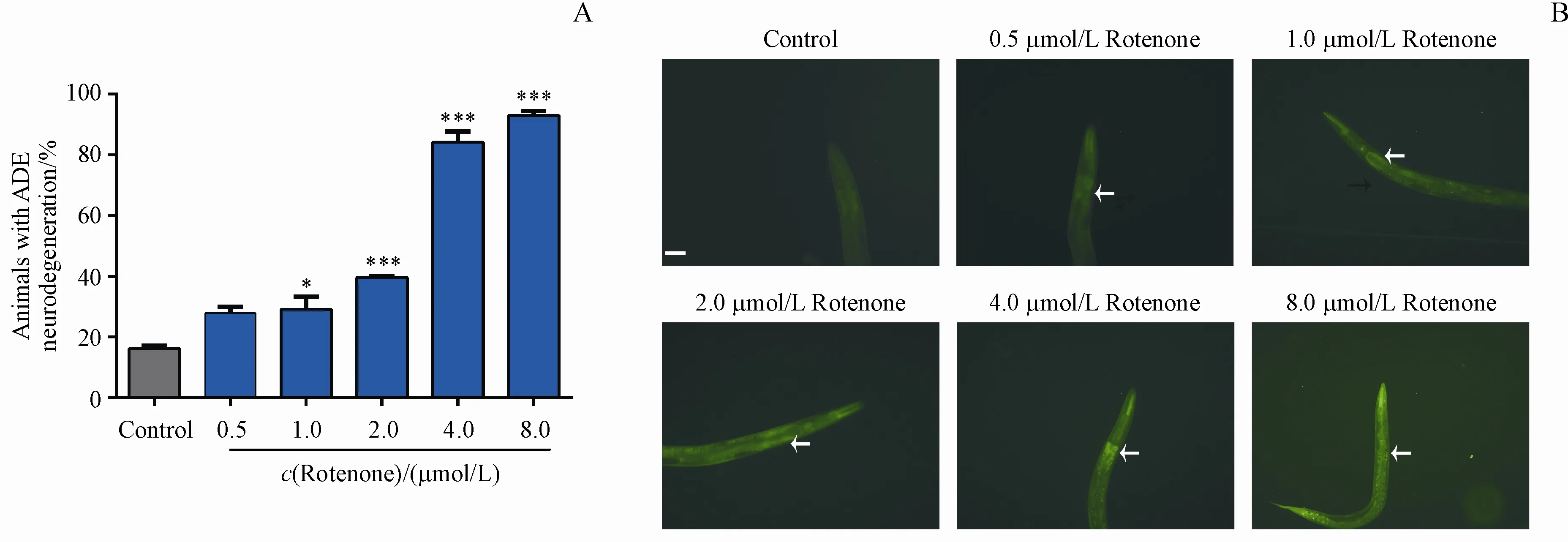
Figure2 Evaluation of neuronsand mito-UPRin C.elegans strain with zcIs9;otIs181 with exposuretodifferent concentrationsof rotenone(xˉ±s,n=20-30)
3.3 VHL抑制劑對PD線蟲模型存活情況的影響
采用不同濃度的VHL抑制劑VH298(400,200和100μmol/L)進行提前干預,次日采用終濃度為2.0μmol/L魚藤酮與不同濃度的VH298共同孵育線蟲,1 d后在體視顯微鏡下觀察并記錄各組線蟲死亡情況。結果如圖3所示,與模型組相比,100,200和400μmol/L的VH298均能在一定程度上降低2.0μmol/L魚藤酮誘導的線蟲死亡率,說明VHL抑制劑能夠緩解魚藤酮導致線蟲模型的死亡。
3.4 VHL抑制劑對PD線蟲模型多巴胺能神經元退化情況的影響
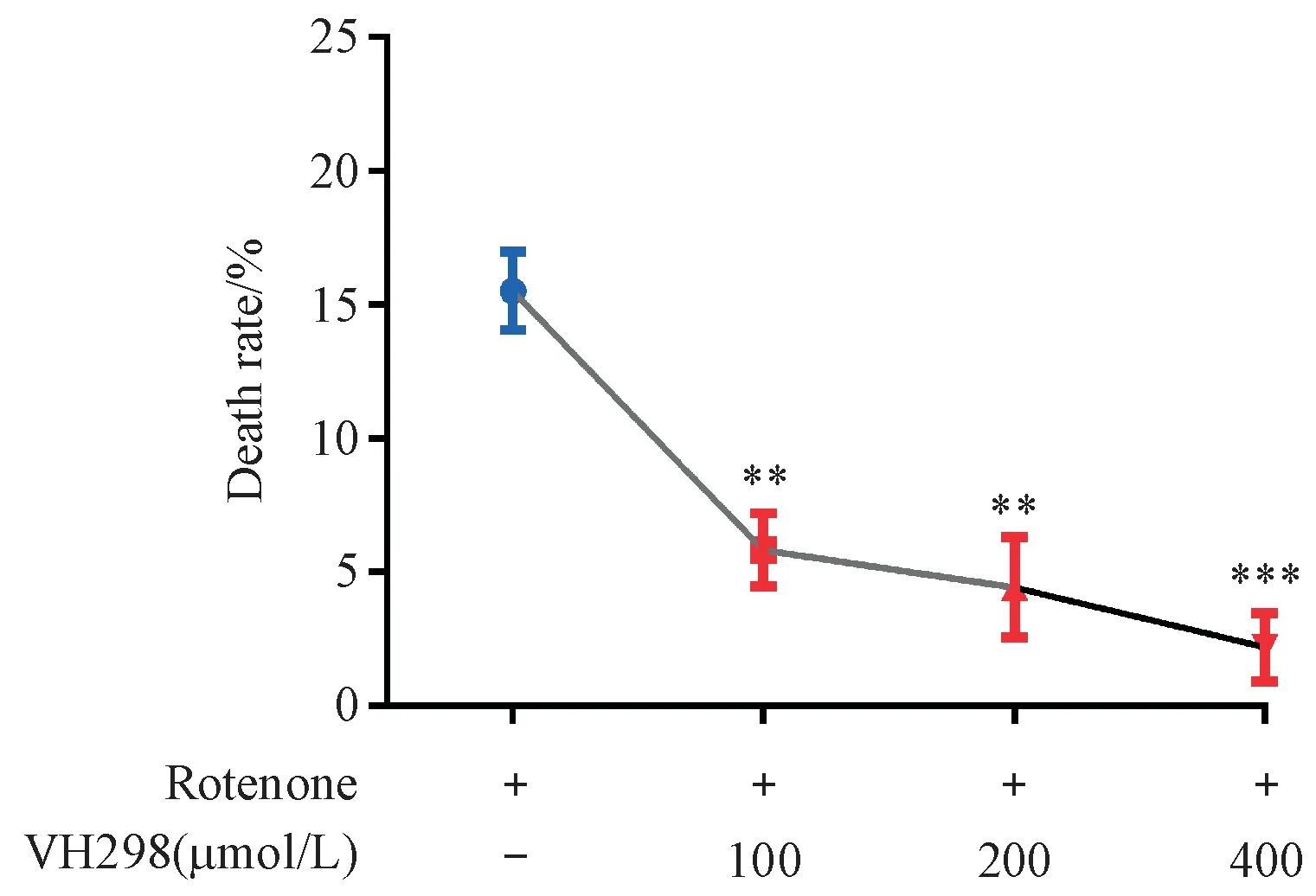
Figure 3 Effects of VHL inhibitor VH298 on death in C.elegans strain with zcIs9;otIs181 with exposure to 2.0μmol/L rotenone(xˉ±
首先采用不同濃度的VHL抑制劑VH298(400,200和100μmol/L)進行提前干預,次日采用終濃度為2.0μmol/L魚藤酮與不同濃度的VH298共同孵育線蟲,1 d后在體視顯微鏡下觀察并記錄各組L4期線蟲多巴胺能神經元退化情況。結果如圖4所示,與模型組相比,400μmol/L的VH298有效緩解魚藤酮誘導的ADE神經元退化情況,說明VHL抑制劑對魚藤酮所致線蟲模型具有一定的多巴胺能神經元保護作用。
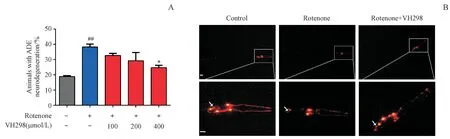
Figure4 Effects of VHL inhibitor on neurons in C.elegans strain with zcIs9;otIs181 with exposure to 2.0μmol/L rotenone(xˉ±s,n=20-30)
3.5 VHL抑制劑對PD線蟲模型運動能力的影響
采 用VHL抑 制 劑VH298(400,200和100 μmol/L)和魚藤酮(2.0μmol/L)處理線蟲,體視顯微鏡下觀察L4期線蟲頭部擺動、身體彎曲情況。結果如圖5所示,VH298能在一定程度上緩解魚藤酮導致的線蟲頭部擺動頻率和身體彎曲頻率異常情況,說明VHL抑制劑能夠減少魚藤酮所致PD線蟲模型的行為學異常。
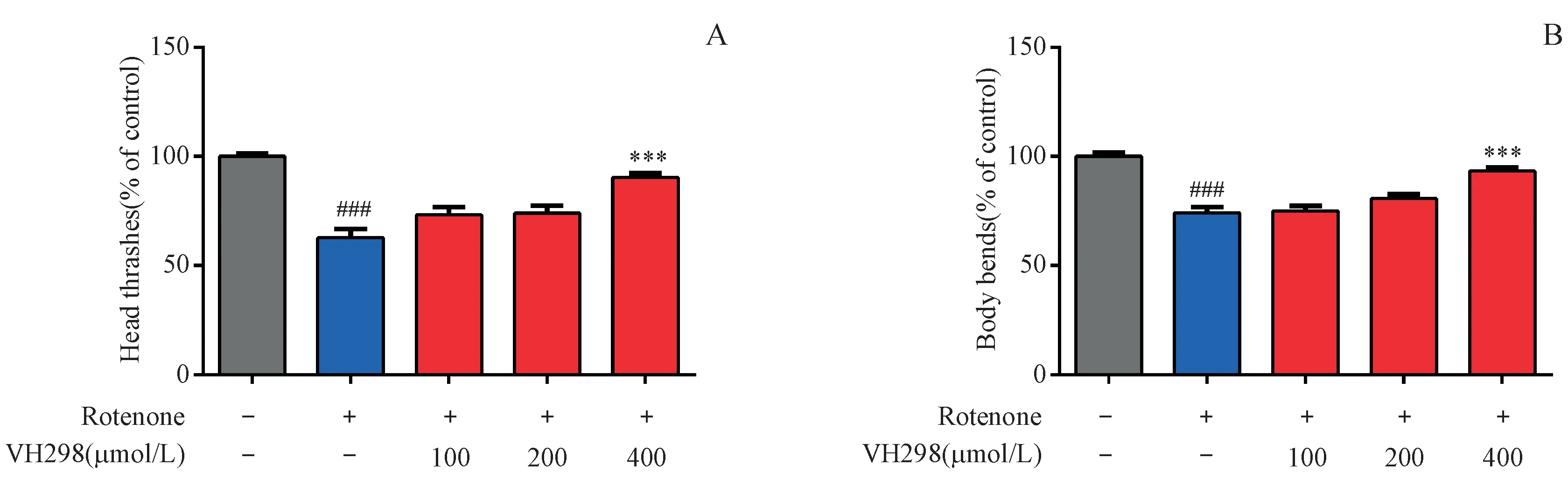
Figure 5 Effects of VHL inhibitor VH298 on head thrashes and body bends in C.elegans strain with zcIs9;otIs181 with exposure to 2.0μmol/L rotenone(xˉ±s,n=20-30)
3.6 VHL抑制劑對PD線蟲模型覓食行為的影響
采 用VHL抑 制 劑VH298(400,200和100 μmol/L)和魚藤酮(2.0μmol/L)處理線蟲,體視顯微鏡下觀察線蟲覓食情況。結果如圖6所示,與模型組相比,VH298處理后線蟲的菌落接觸率在4,12和24 h均有一定程度的增加,其中400μmol/L VH298處理后線蟲的菌落接觸率在12和24 h時顯著增加。以上結果說明,VHL抑制劑能改善魚藤酮所致PD線蟲模型的覓食行為異常情況。
4 討 論
本研究采用神經毒性劑魚藤酮處理轉基因線蟲zcIs9;otIs181,該轉基因線蟲出現異常死亡、神經元異常退化以及異常激活mito-UPR的情況,模擬了類PD樣癥狀如多巴胺能神經元丟失和運動障礙;通過VHL抑制劑VH298干預PD線蟲模型,發現VH298能促進PD線蟲模型存活,緩解ADE多巴能神經元退化情況,并且改善PD線蟲模型運動障礙如頭部擺動頻率和身體彎曲頻率以及多巴胺依賴性行為如覓食行為異常等情況。綜上,魚藤酮可能通過引起轉基因線蟲zcIs9;otIs181線粒體損傷,導致線粒體穩態失衡,進而誘發線蟲異常死亡及神經元退化;VHL抑制劑VH298能減少魚藤酮導致的轉基因線蟲zcIs9;otIs181異常死亡;可通過保護多巴胺能神經元進而改善由多巴胺能神經元丟失所導致的運動行為異常和覓食行為異常。本研究為VHL抑制劑治療帕金森病奠定了一定的理論基礎。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
