2018—2019年山東地區(qū)豬繁殖與呼吸綜合征病毒分子流行病學調(diào)查
2021-07-06 14:01:12梁俊超王宗升劉紅祥郝堯光王輝之
中國獸醫(yī)雜志 2021年2期
梁俊超 , 李 坤 , 袁 飛 , 王宗升 , 劉紅祥 , 郝堯光 , 王輝之 , 劉 東 , 于 錄
(1.吉林大學動物醫(yī)學學院 人獸共患病研究所 人獸共患病研究教育部重點實驗室 , 吉林 長春 130062 ; 2.青島易邦生物工程有限公司 動物基因工程疫苗國家重點實驗室 , 山東 青島 266000)
豬繁殖與呼吸綜合征(Porcine reproductive and respiratory syndrome,PRRS)是一種高度接觸性傳染病,豬繁殖與呼吸綜合征病毒(Porcine reproductive and respiratory syndrome virus,PRRSV)是PRRS病原,該病毒是一種有囊膜的單股正鏈RNA病毒,基因組大小15 kb左右,包含9個開放閱讀框。GP5蛋白由ORF5基因編碼,含有PRRSV主要的中和抗原表位,在誘導中和抗體及免疫交叉保護方面具有重要作用[1]。據(jù)ORF5基因序列分析[2],我國PRRSV流行毒株主要為基因Ⅱ型,且主要為譜系1、譜系3、譜系5和譜系8。
本試驗通過收集2018—2019年山東地區(qū)PRRSV臨床可疑樣品,對陽性樣品中PRRSV毒株進行ORF5基因測序分析,了解近年山東地區(qū)PRRSV的流行趨勢,旨為今后PRRSV的防控提供理論依據(jù)和參考。
1 材料與方法
1.1 病料收集處理 2018—2019年從山東多個地區(qū)(如青島、濰坊、德州、聊城等)采集493份疑似PRRSV感染的豬脾臟、肺臟等病料,病料經(jīng)研磨處理后,按照文獻[3]介紹的RT-PCR方法進行抗原檢測。檢測ORF5基因所用的引物:上游:5′-CATTTCATGACACCTGAGACCA-3′;下游:5′-AGAGCATATATCATCACTGGCG-3′。RT-PCR擴增條件:94 ℃預變性2 min;94 ℃變性15 s,54 ℃退火30 s,68 ℃延伸1 min,32個循環(huán);68 ℃延伸10 min。
1.2 主要試劑 RNA提取試劑盒,購自TaKaRa公司;高保真反轉(zhuǎn)錄試劑盒,購自羅氏公司;LA-Taq DNA聚合酶,購自寶生物工程(大連)有限公司。
1.3 PRRSVORF5基因測序 將PRRSV RT-PCR陽性產(chǎn)物序列送至生工生物工程(上海)股份有限公司進行測序。
1.4 核苷酸進化分析 通過軟件DNASTAR Lasergene對送檢測序PRRSVORF5基因進行序列同源性比對分析,利用MEGA 6.0軟件對所測PRRSVORF5基因序列進行遺傳進化分析。
1.5 氨基酸進化分析 通過軟件DNASTAR Lasergene對送檢測序PRRSV GP5蛋白氨基酸位點進行分析,并基于ORF5基因序列,通過NetNGlyc l.0 Sever在線軟件(http://www.cbs.dtu.dk/services/NetNGlyc/)預測并分析相應氨基酸序列的糖基化位點。
2 結(jié)果
2.1 RT-PCR檢測 將2018—2019年山東省多個地區(qū)送檢疑似PRRSV病料共493份進行RT-PCR檢測,目的條帶大小為603 bp,檢測結(jié)果若出現(xiàn)目的條帶,表明病料中PRRSV抗原檢測結(jié)果為陽性。部分RT-PCR檢測結(jié)果見圖1。送檢病料RT-PCR檢測結(jié)果見表1。從表1可知,2018年和2019年送檢病料中,PRRSV陽性率分別為20.79%和24.09%,其中保育仔豬病料PRRSV陽性率均為最高,分別為37.93%和29.89%。

圖1 ORF 5基因的PCR擴增Fig.1 Amplification of ORF 5 gene by PCRM:DL2 000 DNA相對分子質(zhì)量標準; 1~12:送檢樣本ORF5基因片段; 13:陰性對照; 14:陽性對照;15~18:送檢樣本ORF5基因片段M:DL2 000 DNA marker; 1-12:ORF5 gene fragment of samples; 13:Negative control; 14:Positive control; 15-18:ORF5 gene fragment of samples

表1 2018—2019年不同送檢病料PRRSV陽性率Table 1 Positive rate of PRRSV in different samples from 2018 to 2019 (%)
2.2 PRRSVORF5基因同源性比對分析 從PRRSV陽性樣品中挑選34份PRRSV的PCR產(chǎn)物送檢進行ORF5基因測序,經(jīng)比對后確認得到34個603 bp的樣品PRRSVORF5基因序列。樣品PRRSVORF5基因序列按照“送檢年月-樣品編號-送檢市區(qū)縮寫”進行編號。34個樣品PRRSVORF5基因序列與相應譜系標準毒株序列通過DNASTAR軟件進行比對,結(jié)果見表2和圖2。從圖2可知,34個樣品PRRSVORF5基因序列之間同源性為81.3%~100%,與基因Ⅱ型PRRSV同源性高于基因Ⅰ型PRRSV,與基因Ⅱ型PRRSV的4個譜系代表毒株同源性為81.7%~99.3%。

表2 34個PRRSV ORF 5基因序列與標準毒株同源性對比Table 2 The homology of 34 PRRSV ORF 5 gene sequences compared with the standard strains (%)

圖2 34個PRRSV ORF 5基因核苷酸同源性分析Fig.2 Nucleotide homology analysis of 34 PRRSV ORF 5 gene
2.3 PRRSVORF5基因遺傳進化分析 34個樣品PRRSV的ORF5基因序列通過MEG 6.0軟件繪制遺傳進化樹,見圖3。從圖3和表3中可知,2018—2019年山東地區(qū)34個樣品PRRSV主要分布在譜系1和譜系8,占比分別為55.88%和41.18%。從表3中可知,2018年譜系1和譜系8中樣品PRRSV分別占比為50.00%和41.67%;2019年譜系1和譜系8中樣品PRRSV分別占比為59.10%和40.90%。

圖3 34個PRRSV ORF 5基因序列進化樹分析Fig.3 Evolutionary tree analysis of 34 PRRSV ORF 5 gene sequences?:譜系1、3、5和譜系8的PRRSV標準毒株?:PRRSV standard strain of lineage 1,3,5 and lineage 8

表3 2018—2019年34個PRRSV在不同譜系占比Table 3 The proportion of 34 PRRSV in different lineages from 2018 to 2019
2.4 PRRSV GP5蛋白氨基酸位點變異分析 通過DNASTAR軟件將34個樣品PRRSV GP5蛋白氨基酸序列與標準毒株進行比對,結(jié)果見圖4。從圖4可知,34個樣品PRRSV GP5蛋白氨基酸以點突變?yōu)橹鳎匆姴迦胪蛔儯?個樣品PRRSV在36aa處存在缺失突變。信號肽區(qū)域aa1-25、aa24-39及aa57-61三個區(qū)域氨基酸點突變集中程度較高。

圖4 34個PRRSV GP5蛋白氨基酸同源性分析Fig.4 Amino acid homology analysis of 34 PRRSV GP5 proteins3個方框: 3個PRRSV在36aa處存在缺失突變3 boxes: Deletion mutation of 3 PRRSV was found at 36aa
aa27-30(V27LVN30)是非中和抗原表位,又稱誘餌表位,aa37-45(S37HLQLIYNL45)是中和抗原表位(Protein neutralization epitope,PNE),其中識別位點被認為是H38和 I42YN44,而結(jié)合位點被認為是L39QL41[4-7]。從圖4可知,在誘餌表位區(qū)域,不同譜系樣品PRRSV氨基酸位點突變分別為V27→A27、L28→F28、L28→P28、N30→S30和N30→D30。在中和表位區(qū)域,不同譜系樣品PRRSV氨基酸位點突變分別為H38→N38、H38→Y38、L39→S39、L39→T39、L39→I39和N44→S44。據(jù)相關(guān)研究報道,R13和R151是PRRSV潛在的毒力位點,與病毒毒力密切相關(guān)[8-9]。從圖4可知,不同譜系樣品PRRSV的R13和R151位點發(fā)生突變,分別為R13→H13、R13→Q13、R151→K151和R151→T151。
從表4可知,2018—2019年34個樣品PRRSV的潛在毒力位點突變率最高,為82.35%。2018—2019年不同譜系的樣品PRRSV潛在毒力位點、中和表位位點和誘餌表位位點均發(fā)生不同程度突變。

表4 2018—2019年34個PRRSV GP5蛋白氨基酸位點變異分析Table 4 Amino acid variation analysis of 34 PRRSV GP5 proteins from 2018 to 2019
2.5 PRRSV GP5蛋白潛在N-糖基化位點變異分析 通過NetNGlyc l.0 Sever在線軟件,對34個樣品PRRSV GP5蛋白氨基酸序列的N-糖基化位點進行預測分析,結(jié)果見表5。從表5可知,與NADC 30相比,譜系1中2018年和2019年P(guān)RRSV糖基化位點增加1個的樣品占比分別為50.00%(3/6)和53.85%(7/13),2019年糖基化位點增加2個的樣品占比為23.08%(3/13);與JXA1相比,譜系8中2018年和2019年P(guān)RRSV糖基化位點減少1個的樣品占比分別為40.00%(2/5)和44.44%(4/9)。
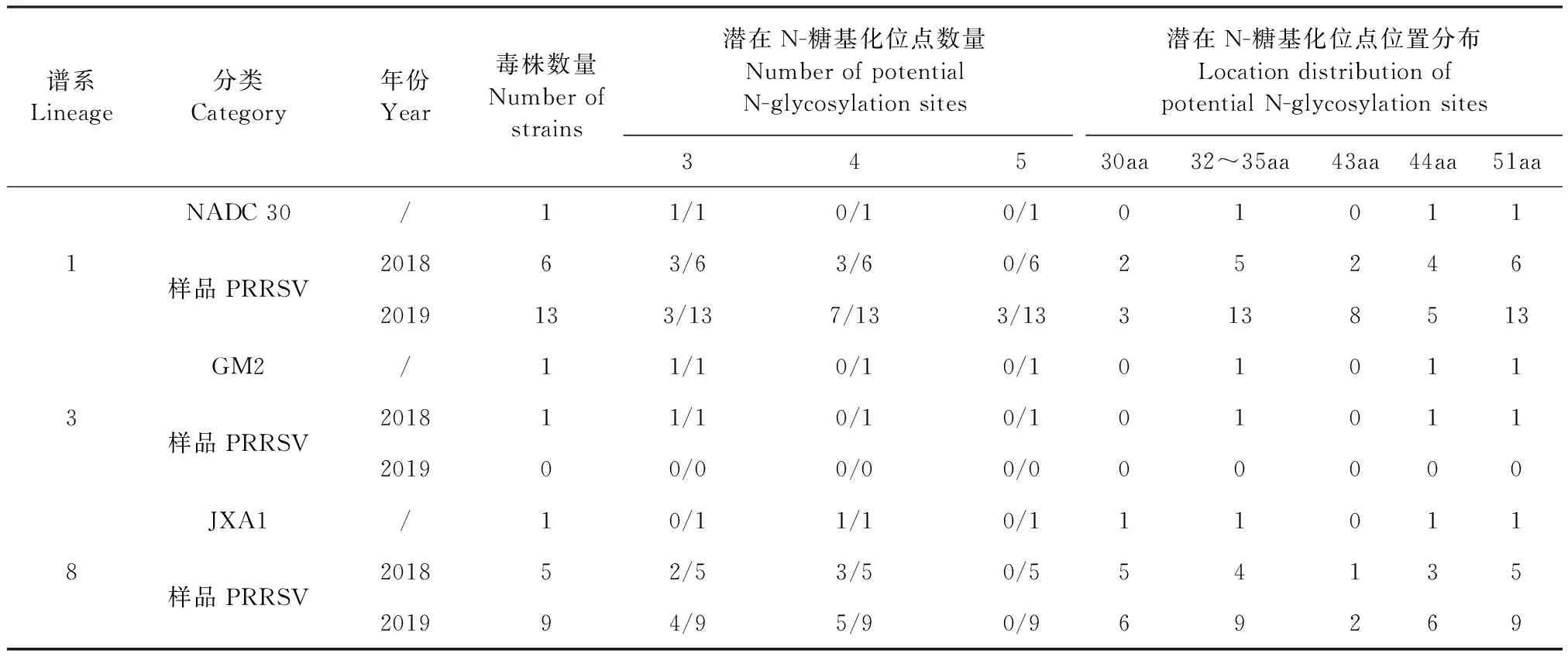
表5 2018—2019年34個PRRSV的GP5蛋白潛在N-糖基化位點數(shù)量及位置分布Table 5 Number and location of potential N-glycosylation sites of 34 PRRSV GP5 proteins from 2018 to 2019
3 討論
目前我國PRRSV感染仍十分普遍,且已成為豬場的常在疾病[10]。從RT-PCR檢測結(jié)果可知,2018年和2019年全年P(guān)RRSV抗原陽性率較往年報道相比有所下降[11],但感染率仍較高,其中保育仔豬病料PRRSV陽性率最高,與姚亞輝統(tǒng)計結(jié)果相符[12]。
2018—2019年山東地區(qū)共測序34個PRRSV樣品,經(jīng)ORF5基因同源性比對分析發(fā)現(xiàn),34個PRRSV樣品與經(jīng)典疫苗毒株VR2332、MLV及歐洲株LV同源性相對較低,與譜系8中HP PRRSV JXA1、HuN4、TJ及譜系1中的NADC 30 PRRSV的同源性相對較高。據(jù)報道,PRRSV同源性高低影響PRRS疫苗毒株的田間保護效果[13]。從遺傳進化分析結(jié)果可知,譜系1中樣品PRRSV即NADC30 like PRRSV或已成為山東地區(qū)田間流行優(yōu)勢野毒毒株,這與劉艷艷報道結(jié)果相符[11]。
據(jù)報道,PRRSV的誘餌表位與中和表位在中和抗體產(chǎn)生的過程中發(fā)揮關(guān)鍵作用,尤其在感染早期,可抑制免疫系統(tǒng)識別中和表位,導致中和抗體產(chǎn)生延遲[14]。此外,PRRSV重要抗原表位氨基酸位點發(fā)生突變,將影響抗原抗體結(jié)合,有利于病毒免疫逃避,導致PRRSV疫苗株的免疫效果不佳甚至免疫失敗[15-17]。從氨基酸位點變異分析結(jié)果可知,2019年與2018年相比,譜系1中樣品PRRSV誘餌表位及中和表位氨基酸位點突變率呈上升趨勢,潛在毒力位點突變率高,由此推測PRRSV重要抗原位點突變可能更有利于病毒的免疫逃避,而譜系8中樣品PRRSV誘餌表位及中和表位氨基酸位點突變率呈下降趨勢,潛在毒力位點突變率呈上升趨勢,由此推測可能由于疫情、存欄影響導致該PRRSV逃避機體免疫的壓力下降,使其突變率下降,但其重要抗原位點仍在不斷變異,增加防控難度。
研究發(fā)現(xiàn),中和表位附近N-糖基化位點具有屏蔽作用,且該位點缺失能增強免疫原性、敏感性及中和抗體的中和能力,不利PRRSV的免疫逃避[4,8,14],此外,PRRSV的N-糖基化位點數(shù)目、位置的變化影響抗體對病毒的中和能力及病毒對宿主細胞的侵染力[18]。從GP5蛋白潛在糖基化位點變異分析結(jié)果可知,2019年與2018年相比,譜系1中樣品PRRSV相比NADC 30 PRRSV的糖基化位點數(shù)量增加的比例明顯升高,由此推測這可能更利于PRRSV逃避機體的免疫,增加PRRSV的防控難度;譜系8中樣品PRRSV相比JXA1的糖基化位點數(shù)量減少的比例略有升高,由此推測這可能更有利于機體免疫識別,PRRSV免疫防控更需重視。
4 結(jié)論
山東地區(qū)現(xiàn)階段PRRSV呈現(xiàn)多樣化、復雜化,其中NADC 30 like PRRSV或已成為田間流行的優(yōu)勢野毒毒株,這或?qū)е翽RRSV疫苗保護效果下降。結(jié)合臨床生產(chǎn),PRRS防控不能單純依賴疫苗,更需提高豬場生物安全水平和飼養(yǎng)管理水平,豬群健康對PRRS防控至關(guān)重要。綜上所述,PRRSV防控需綜合防控。
