紫色紅曲霉磁受體IscA1的基因克隆及功能分析
2021-06-28 07:42:00黃勝男劉應保張佳蘭高夢祥
西北農業學報 2021年5期
關鍵詞:生物
黃勝男,劉應保,張佳蘭,高夢祥
(長江大學 生命科學學院,湖北荊州 434025)
磁場在保護地球免遭高能宇宙射線的損傷方面至關重要[1],同時,還能顯著調節地球生物的生長和代謝[1-3],即磁致生物學效應。過去的幾十年,大量的研究報道了動物、微生物等生物中存在磁場生物學效應[3-7]。然而,關于生物的磁感應機制的研究相對較少。
目前,主流的生物磁感應機制主要包括磁顆粒介導磁感應假說和隱花色素(cryptochrome,CRY)的化學自由基對磁感應假說[1-2]。前者認為生物磁感應主要由特定細胞中微小磁顆粒簇(Fe3O4等)介導實現的,比如趨磁細菌的磁小體,而且該裝置在基因組上有特定的基因編碼區,比如磁小體島。在高等生物中,雖然發現了相關的磁顆粒,但是一直未發現相關的編碼基因。隨后,隱花色素CRY的化學自由基對磁感應的假說被提出。該假說基于藍光受體CRY蛋白與輔因子FAD通過光誘導的自由基響應磁感應。大量的研究結果支持該假說。近10多年來,本課題組以絲狀真菌為研究對象,發現了磁場的各種生物學效應[8-15 ]。同時,通過分子生物學手段結合蛋白質組學,重點研究了藥食兩用的絲狀真菌紫色紅曲霉的磁感應機制。然而在大量差異表達蛋白中,未發現上述磁感應假說中的磁顆粒和CRY及其相關蛋白[16],而是發現其他蛋白,比如最關鍵的是鐵硫簇(iron sulfur cluster,ISC/Fe-S cluster)生物合成系統中的成員蛋白Isu1、IscS、IscA1、 Grx5等。其中,IscA1是最近在果蠅(Drosophilamelanogaster)中發現的新的磁受體MagR[17]。IscA1在不同生物中高度保守,除了作為磁受體,該蛋白還在能量代謝、輔因子、鐵硫儲備、基因表達調控等過程發揮重要作用[1]。最近,Zhou等[18]在紅色紅曲霉(MonascusruberM7)中克隆了IscA1基因,被認為是該菌的候選磁受體基因。由此推測, IscA1可能是紫色紅曲霉中的磁受體蛋白,該蛋白也是介導該菌磁感應過程中的關鍵蛋白。基于此,本研究采用同源克隆方法,經PCR擴增和DNA測序獲得紫色紅曲霉的磁受體基因IscA1的編碼序列,并借助生物信息學工具初步揭示其生物學功能。
1 材料與方法
1.1 菌種和引物
所用菌種為紫色紅曲霉(MonascuspurpureusSKY219),為長江大學生命科學學院微生物學實驗室保藏菌種。所用引物由武漢天一輝遠生物科技有限公司合成,具體序列見表1。
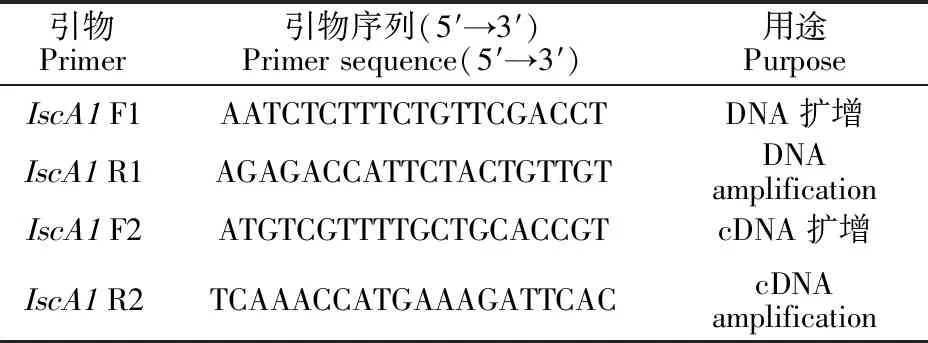
表1 所用引物Table 1 Primers used in this study
1.2 主要試劑和儀器
DL5000 DNA Marker(MD102-02),DL2000 DNA Marker(MD101-02),2 × Phanta Master Mix(P511-01),DNA純化試劑盒(DC301-1),總RNA提取試劑(RNA isolater Total RNA Extraction Reagent,R401-01),反轉錄試劑盒(HiScript Ⅱ Q RT SuperMix for qPCR,R223-1)購自南京諾唯贊生物科技有限公司;十六烷基三甲基溴化銨(Hexadecyl trimethyl ammonium Bromide,CTAB),氯仿,無水乙醇,異丙醇,苯酚購自國藥集團化學試劑有限公司。
ZF-208凝膠成像系統為上海嘉鵬科技有限公司產品;C1000 TouchTM型PCR儀,PowerPac Basic型電泳儀,GelDocTM XR型凝膠成像儀為美國Bio-rad公司產品。
1.3 方法
1.3.1 紫色紅曲霉(M.purpureusSKY219)磁受體IscA1基因擴增引物設計 根據近緣種紫色紅曲霉(M.purpureusNRRL 1596)的IscA1基因序列(Monpu1_501839)[18]設計引物(表1),擴增本研究所用紫色紅曲霉(M.purpureusSKY219)中IscA1的DNA和編碼區序列,預期擴增的長度分別為1 331和735 bp。
1.3.2IscA1的DNA擴增 基因組提取參照文獻[14]方法。以M.purpureusSKY219基因組為模板,擴增IscA1的DNA序列。PCR反應體系為:基因組模板為1 μg,引物(IscA1F1/IscA1R1)各2 μL,2 × Phanta Master Mix 25 μL,用去離子水補足50 μL。PCR反應條件為:95 ℃ 3 min;95 ℃ 15 s、50 ℃ 15 s、72 ℃ 45 s,30個循環; 72 ℃ 5 min。PCR產物經1% 瓊脂糖凝膠電泳檢測,并對目的DNA片段純化,然后送至武漢天一輝遠生物科技有限公司進行DNA 測序。
1.3.3IscA1的cDNA擴增 首先提取M.purpureusSKY219的總RNA,然后逆轉錄得到的cDNA進行擴增IscA1的編碼區序列。總RNA提取方法如下:收集培養6 d的菌體,在液氮預冷的研缽中速凍并充分研磨后,取適量的粉末放于含1 mL RNA提取試劑的1.5 mL無酶離心管中,充分混勻后,加入1/5體積的氯仿,混勻,靜置5 min,4 ℃、10 000 r/min離心10 min,取上清加入等體積預冷的異丙醇沉淀30 min,離心 10 min,棄上清,沉淀用75%乙醇洗滌兩次,離心 10 min,棄上清,沉淀晾干,用DEPC水溶解,經瓊脂糖凝膠電泳檢測。得到總RNA之后,用反轉錄試劑盒得到cDNA。接著以cDNA為模板擴增IscA1的編碼區,反應體系為:cDNA 1 μg,引物各2 μL,2 × Phanta Master Mix 25 μL,用去離子水補足50 μL。PCR反應條件為:95 ℃ 3 min,95 ℃ 15 s、58 ℃ 15 s、72 ℃ 30 s,30個循環, 72 ℃ 5 min。PCR產物經瓊脂糖凝膠電泳檢測后,純化目的DNA,并進行DNA測序。
1.3.4IscA1的生物信息學分析 將IscA1的編碼序列進行多序列對位排列分析搜索其保守結構域,并構建進化樹;預測其蛋白一級、二級、三級結構及蛋白互作網絡等,具體工具和使用方法見表2。
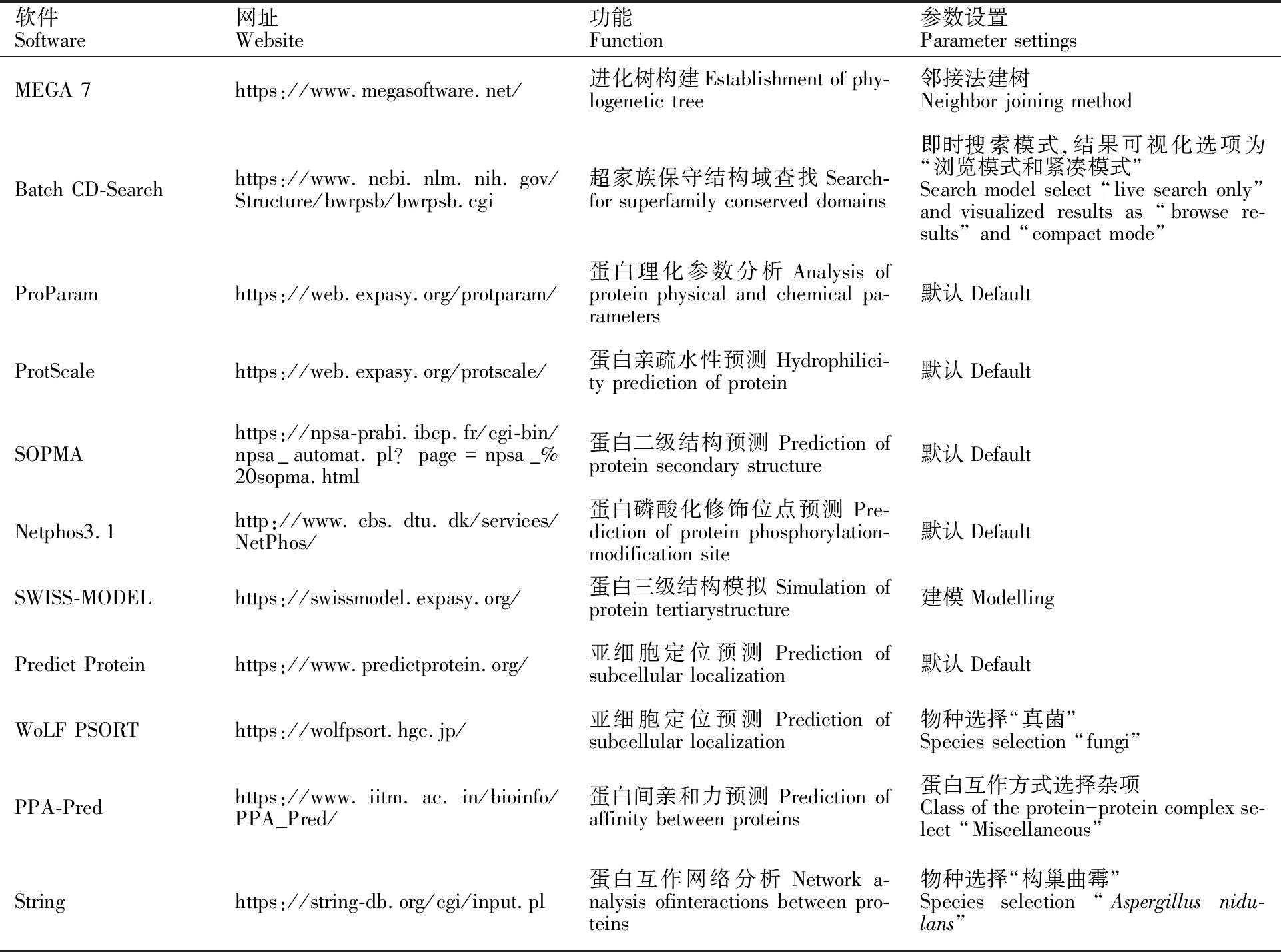
表2 所用到的分析工具和方法Table 2 Analysis tools and methods used in this study
2 結果與分析
2.1 IscA1的DNA擴增
根據紫色紅曲霉近緣種的IscA1基因設計引物,以M.purpureusSKY219基因組(圖1-a)為模板進行PCR擴增,擴增產物的電泳結果(圖1-b)顯示,在電泳膠的第2泳道的1 500 bp附近有1條特異性條帶,與IscA1的預期的DNA序列大小(1 331 bp)相符,即目的DNA擴增成功。DNA測序結果發現,M.purpureusSKY219的IscA1基因的DNA序列與M.purpureusNRRL 1596的IscA1的基因序列的相似度為100%(覆蓋度為100%),即兩者IscA1的DNA序列完全相同。
2.2 IscA1的cDNA擴增
以M.purpureusSKY219的總RNA(圖2-a)反轉錄得到的cDNA為模板進行擴增,結果顯示(圖2-b),在第2泳道上,750 bp附近有1條特異性條帶,與預期大小(735 bp)相符。IscA1編碼區測序與M.purpureusNRRL 1596的IscA1的編碼區完全相同,該序列經NCBI ORFfinder分析發現,IscA1共編碼244氨基酸(圖3)。
2.3 IscA1保守結構域和蛋白家族歸屬分析
將M.purpureusSKY219的 IscA1與腹黑果蠅(D.melanogaster)、紅色紅曲霉(M.ruber)、釀酒酵母(Saccharomycescerevisiae)、大腸桿菌(Escherichiacoli)、枯草芽孢桿菌(Bacillussubtilis)、人(Homosapiens)、小鼠(Musmusculus)、原鴿(Columbalivia)、斑馬魚(Daniorerio)、秀麗隱桿線蟲(Caenorhabditiselegans)、擬南芥(Arabidopsiathaliana)、大豆(Glycinemax)等物種的 IscA1進行多序列對位排列分析,結果發現,M.purpureusSKY219的 IscA1與其他物種的磁受體蛋白在序列上高度相似,尤其是第172、236和238位的半胱氨酸Cys(C,星號“*”標識)最為保守,為鐵硫簇結合位點(圖4)。
在線工具Batch CD-Search搜索保守結構域的結果發現,M.purpureusSKY219的 IscA1與其他物種的磁受體蛋白都具有典型的Fe-S簇生物合成超家族(TIGR00049)結構域(圖5)。
上述蛋白構建的進化樹圖顯示,M.purpureusSKY219的 IscA1與M.ruber、S.cerevisiae真菌的磁受體 IscA1聚在一個進化枝上,同時與細菌和植物聚在另一個大的進化枝上(圖6)。
2.4 IscA1的理化性質分析
ProParam分析的結果表明, IscA1為堿性蛋白(pI為10.05),分子質量為27 106 u,不穩定系數為59.99(>40),為不穩定蛋白。其中絲氨酸(Ser)、精氨酸(Arg)和丙氨酸(Ala)含量最為豐富,分別占比為12.3%、9.0%和8.2%。親水性指數為負值(-0.575),即為親水蛋白。
2.5 IscA1的親疏水性預測
Protscale的親疏水性分析發現,IscA1蛋白的絕大部分氨基酸的親水性數值均為負值(圖7),與上述理化性質預測結果一致,即為親水 蛋白。
2.6 IscA1的二級結構預測
二級結構預測的結果發現,IscA1的二級結構元件α-螺旋、β-轉角、無規則卷曲和延伸鏈的比例分別為22.13%、2.05%、62.70%、13.11%,以無規則卷曲為主(圖8)。
2.7 IscA1的磷酸化修飾預測
Netphos 3.1預測的結果發現,IscA1在Ser、Thr和Tyr3個位點被磷酸化修飾的個數分別為26、21、13個(圖9)。表明紫色紅曲霉中的IscA1可能被其他激酶磷酸化修飾。
2.8 IscA1的三級結構模擬
軟件SWISS-MODEL模擬的IscA1的三級結構如圖10所示。IscA1以細長嗜熱聚球藻(ThermosynechococcuselongatusBP-1)為模板,二者的相似度為37%,覆蓋度為43%。模擬結果顯示,三維結構以無規則卷曲和β-折疊為主,還有比較典型的α-螺旋(圖10)。
2.9 IscA1的亞細胞定位分析
Predict Protein預測的結果顯示,IscA1主要定位于線粒體(圖11)。同時WoLF PSORT預測的亞細胞定位發現,IscA1也是定位于線粒體,二者預測結果一致。
2.10 IscA1的互作蛋白預測
String預測的互作蛋白的結果顯示(圖12),IscA1可以與Fe-S簇裝配輔助蛋白IscA2(ANIA_05953)和谷氧還蛋白Grx 5(AN4304.2)發生非特異性結合。而且,IscA1和Grx可以被分子伴侶DnaJ蛋白激活(AN4304.2)。Grx 5、DnaJ和IscA1共同參與Fe-S簇的生物合成和傳遞過程,因此,在Fe-S簇組裝和傳遞過程中會發生不同的互作[18]。PPA-Pred預測的蛋白間親和力的結果顯示,IscA1與D.melanogaster的藍光受體蛋白CRY之間的親和力較強(自由能△G=-45.99 kJ/mol,解離常數Kd=1.03e-8mol),而與絲狀真菌構巢曲霉的藍光受體LreA、LreB均有更強的親和力(IscA1/LreA:△G= -55.52 kJ/mol,Kd=1.9e-10mol;IscA1/LreB:△G=-42.91 kJ/mol,Kd=3.04e-8mol),這些結果說明磁受體IscA1與不同的藍光受體蛋白(CRY、LreA、LreB)均可以互作,與新近報道的磁受體IscA1-藍光受體CRY復合物共同感應磁場的理論相符[16]。
3 結論與討論
IscA1是Fe-S簇生物合成系統中的必需蛋白,在各個物種中高度保守[1]。本課題組前期研究發現,該蛋白是紫色紅曲霉磁感應中的關鍵蛋白。最新的研究揭示,IscA1是新的磁受體[16]。因此,本研究基于同源克隆原理,通過PCR擴增和DNA測序,獲得M.purpureusSKY219的IscA1編碼區序列,該基因編碼的蛋白共有244個氨基酸。進一步的生物信息學分析發現,IscA1與真菌的同源蛋白親緣關系較近。該蛋白具有3個最保守的Cys(172、236和238位),并具有典型的Fe-S簇生物合成超家族的保守結構域,這些都是已報道的磁受體IscA1的典型特征[1]。IscA1為堿性蛋白和親水蛋白,二級結構組成與已報道的M.ruberM7的IscA1蛋白的二級結構高度相似[18],均以不規則卷曲為主,其次是α-螺旋、β-轉角。模擬的IscA1三級結構則與解析的T.elongatus的IscA1的晶體結構具有較高的相似度,具有典型的α-螺旋、β-轉角[19]。IscA1具有60個可能的磷酸化修飾位點,主要定位于線粒體。蛋白互作的結果發現,IscA1可被DnaJ激活,并和IscA2、Grx5存在互作關系。在線粒體Fe-S簇生物合成過程中,Fe2+和S原子在支架蛋白IscU上組裝成初生的[2Fe-2S]簇,然后在DnaJ、HSP等蛋白的協助下,將簇傳遞給載體Grx5,然后在IscA1、IscA、IBA57等蛋白復合物的幫助下,將Fe-S “交付”給脫輔基蛋白[19-20]。因此,在Fe-S的組裝和傳遞過程中,IscA1會和上述蛋白發生相互作用。最新的研究提出,磁受體IscA1和藍光受體CRY形成蛋白復合物賦予果蠅等生物磁感應能力[17],本研究也發現,IscA1與不同藍光受體(CRY、LreA、LreB)之間均存在較強的親和力,表明IscA1在紫色紅曲霉中可能與未知的藍光受體蛋白形成復合物賦予該菌磁感應能力。研究結果為全面揭示磁受體IscA1在紫色紅曲霉磁感應機制中的功能奠定基礎。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
