微生物來(lái)源堿性蛋白酶活性提高策略的研究進(jìn)展
2021-06-23 07:55:42袁媛王蕾石亞偉
生物技術(shù)通報(bào) 2021年5期
袁媛 王蕾 石亞偉
(山西大學(xué)生物技術(shù)研究所 教育部化學(xué)生物學(xué)與分子工程重點(diǎn)實(shí)驗(yàn)室,太原 030006)
蛋白酶是指能夠?qū)⒌鞍踪|(zhì)降解成小肽和氨基酸的水解酶,占整個(gè)工業(yè)酶市場(chǎng)的60%。而堿性蛋白酶為在中性至堿性pH范圍內(nèi)具有活性的蛋白酶的統(tǒng)稱(chēng)。1945年,Jaag等[1]最早在地衣芽孢桿菌Bacillus licheniformis中發(fā)現(xiàn)了堿性蛋白酶。微生物蛋白酶大多屬于胞外酶[2],與動(dòng)植物相比,微生物來(lái)源的堿性蛋白酶具有分離純化過(guò)程簡(jiǎn)單、生產(chǎn)周期短且產(chǎn)率高的顯著特點(diǎn)[3]。堿性蛋白酶具有較強(qiáng)的耐堿、耐熱能力以及水解蛋白質(zhì)的活力[4],主要應(yīng)用在洗滌劑工業(yè)上,尤其是洗衣和餐具洗滌劑的生產(chǎn)中。堿性蛋白酶的生產(chǎn)菌種和研究對(duì)象主要是枯草芽孢桿菌(Bacillus subtilis)和地衣芽孢桿菌(Bacillus licheniformis)及短小芽孢桿菌(Bacillus pumilus)[5],如我國(guó)目前使用的地衣芽孢桿菌B.licheniformis2709,短小芽孢桿菌B.pumilus 289和209菌株等[6]。產(chǎn)堿性蛋白酶的放線菌較少,主要來(lái)源于鏈霉菌[7]。另外,某些霉菌也可以產(chǎn)生堿性蛋白酶。目前,我國(guó)堿性蛋白酶研究總體趨勢(shì)發(fā)展較好,但國(guó)內(nèi)企業(yè)仍不能實(shí)現(xiàn)高端堿性蛋白酶制劑的工業(yè)化生產(chǎn),主要原因在于國(guó)外專(zhuān)利技術(shù)的壟斷、國(guó)內(nèi)優(yōu)良的產(chǎn)酶菌株缺乏、酶的單位活力低、洗滌條件下酶的穩(wěn)定性較差等[8-9]。因此,如何提高堿性蛋白酶活力迫在眉睫。
1 產(chǎn)堿性蛋白酶的微生物菌株改造
1.1 傳統(tǒng)菌株誘變改造
這是早期微生物來(lái)源堿性蛋白酶研究工作中常用的一種策略,其原理是利用各種物理因素(常用紫外線和低能離子束)和化學(xué)試劑處理微生物細(xì)胞,使其發(fā)生基因突變,從而引起微生物菌株的遺傳性狀改變進(jìn)而有利于獲得高產(chǎn)堿性蛋白酶菌株[6]。Wang等[10]應(yīng)用物理因素紫外線、60Co-γ射線以及化學(xué)誘變劑NTG對(duì)短小芽孢桿菌B.pumilus BA06進(jìn)行誘變,得到產(chǎn)酶活力提高4倍的突變菌株SCU11。王曉云等[11]通過(guò)對(duì)實(shí)驗(yàn)室保藏菌株枯草芽孢桿菌BC2進(jìn)行紫外誘變后,得到的突變菌株B38的產(chǎn)蛋白酶活性達(dá)到最高,相比出發(fā)菌株(27.68 U/mL)提高了3.14倍。王瑾等[12]對(duì)芽孢桿菌DL12菌株(酶活力為129.7 U/mL)進(jìn)行誘變處理,篩選出1株堿性蛋白酶產(chǎn)量高的菌株,酶活力為172.3 U/mL。傳統(tǒng)誘變改造的方法,雖然具有一定的優(yōu)點(diǎn),但它的弊端也很大,如費(fèi)時(shí)費(fèi)力、盲目性較大、效率低等。
1.2 太空誘變改造
隨著航天事業(yè)的飛速發(fā)展,太空誘變開(kāi)始慢慢興起,其原理是利用衛(wèi)星、飛船等將農(nóng)作物種子或生物菌種帶到太空,使產(chǎn)品因太空中特殊的環(huán)境條件而發(fā)生變異。劉雪等[13]以芝麻香細(xì)菌曲為誘變材料,搭乘“神舟十號(hào)”飛船進(jìn)行空間誘變,返回地面后對(duì)誘變后麩曲中的細(xì)菌進(jìn)行分離篩選,發(fā)現(xiàn)其中有一株產(chǎn)堿性蛋白酶能力最高的菌株,其酶活力是出發(fā)菌株的6.7倍。
1.3 原生質(zhì)體融合改造
該技術(shù)的方法是利用原生質(zhì)體的融合實(shí)現(xiàn)遺傳重組,從而選育高產(chǎn)堿性蛋白酶菌株。潘延云及其團(tuán)隊(duì)將含有堿性蛋白酶基因的pDW2/ B.subtilis BD105的枯草桿菌工程菌與該基因的出發(fā)菌株地衣芽孢桿菌B.licheniformis 2709進(jìn)行原生質(zhì)體融合,篩選獲得了酶活力最高達(dá)到30 778 U/mL的高產(chǎn)堿性蛋白酶的工程菌A16,比出發(fā)菌株高約50%-100%[14]。李宏[15]對(duì)芽孢桿菌 SD-142 的原生質(zhì)體進(jìn)行誘變處理,篩選到一株堿性蛋白酶酶活為2 759 U/mL的突變株。
1.4 基因工程改造
基因工程技術(shù),主要利用DNA重組的原理,將目的基因插入載體,然后轉(zhuǎn)入新的宿主細(xì)胞,構(gòu)建成基因重組工程菌。1985年,Jacobs等[16]首次成功克隆到芽孢桿菌的堿性蛋白酶基因后,利用基因工程技術(shù)構(gòu)建高產(chǎn)蛋白酶工程菌就開(kāi)始被廣大科學(xué)工作者應(yīng)用,為高產(chǎn)菌株的選育開(kāi)辟了新路徑[17]。Tang等[18]將來(lái)自地衣芽孢桿菌B.licheniformis 2709的堿性蛋白酶基因apr克隆到芽孢桿菌穿梭表達(dá)載體pHL中,最終在B.subtilis WB600中表達(dá)得到了高表達(dá)菌株BW-016,表達(dá)量增加了65%。Sareen等[19]將地衣芽孢桿菌B.licheniformis RSP-09-37的堿性蛋白酶基因在大腸桿菌E.coli JM109中進(jìn)行表達(dá)。Lin等[20]成功將地衣芽孢桿菌2709的堿性蛋白酶基因Apr 克隆到表達(dá)載體pET-28b(+)中,得到重組質(zhì)粒pET-28b(+)-Apr。黃磊等[21]將篩選獲得一種高酶活的堿性蛋白酶的編碼基因aprE209克隆入表達(dá)質(zhì)粒pHY-WZX,再轉(zhuǎn)入枯草桿菌B.subtilis WB600中,得到的重組酶的酶活力達(dá)到400 U/mL。利用該種方式構(gòu)建工程菌的堿性蛋白酶的高產(chǎn)菌株,具有極強(qiáng)的目的性,克服了傳統(tǒng)誘變的盲目性[22],具有很大的應(yīng)用潛力。
除了構(gòu)建基因工程菌的方法,還可以通過(guò)基因敲除技術(shù)改造表達(dá)菌株。傳統(tǒng)的基因編輯方法有同源重組整合載體法、反向篩選標(biāo)記法和Cre/loxP定點(diǎn)重組法,其中利用同源重組的原理進(jìn)行靶基因的基因敲除的原理圖,如圖1所示。基因敲除是選擇合適的質(zhì)粒,將靶基因兩邊的同源序列克隆至載體上,轉(zhuǎn)入宿主菌后利用同源重組的原理實(shí)現(xiàn)對(duì)靶基因的置換敲除[23]。現(xiàn)今,CRISPR/Cas9基因編輯方法由于其具有周期短,操作簡(jiǎn)單的優(yōu)點(diǎn),得到人們的青睞。韓登蘭等[24]成功構(gòu)建了芽孢萌發(fā)缺陷型工程菌枯草芽孢桿菌B.subtilis 168ΔsleBΔcwlJ。劉洋等[25]利用pk18質(zhì)粒作為自殺質(zhì)粒并通過(guò)同源重組敲除的方式對(duì)其進(jìn)行敲除,證明了NRPS-A與抗菌肽BBL的合成具有直接關(guān)系。趙銀娟等[26]利用自殺質(zhì)粒pMAD,最終獲得不帶抗性且ClpQY完全敲除的枯草芽孢桿菌B.subtilis 3610菌株。Zhou等[27]通過(guò)單獨(dú)敲除參與孢子形成的調(diào)節(jié)基因(spo0A,sigF和sigE),最終發(fā)現(xiàn)ΔsigF的突變體的堿性蛋白酶酶活力相比野生型菌株高出了約19.7%。Zhou等[28]對(duì)地衣芽孢桿菌2709的lchAC基因和胞外粘多糖編碼的eps簇敲除,通過(guò)篩選不同模塊質(zhì)粒和基因組位點(diǎn)之間最有效的表達(dá)系統(tǒng),進(jìn)一步優(yōu)化了堿性蛋白酶基因(aprE)的表達(dá)。由于淀粉酶和幾丁質(zhì)酶是兩種在宿主菌株中分泌量相對(duì)較高的天然細(xì)胞外蛋白,可阻礙堿性蛋白酶的分泌和表達(dá),所以Zhou等[29]基于CRISPR/Cas9開(kāi)發(fā)了一種高效的地衣芽孢桿菌基因組編輯系統(tǒng)將以上兩種酶的基因敲除,結(jié)果表明,在兩個(gè)基因均缺失的突變體中有效地提高了堿性蛋白酶的產(chǎn)量。近幾年,CRISPR/Cas9在實(shí)驗(yàn)中的應(yīng)用越來(lái)越廣泛,很多課題組也在以此為基礎(chǔ)不斷的創(chuàng)新,相信未來(lái)的發(fā)展前景會(huì)更好。
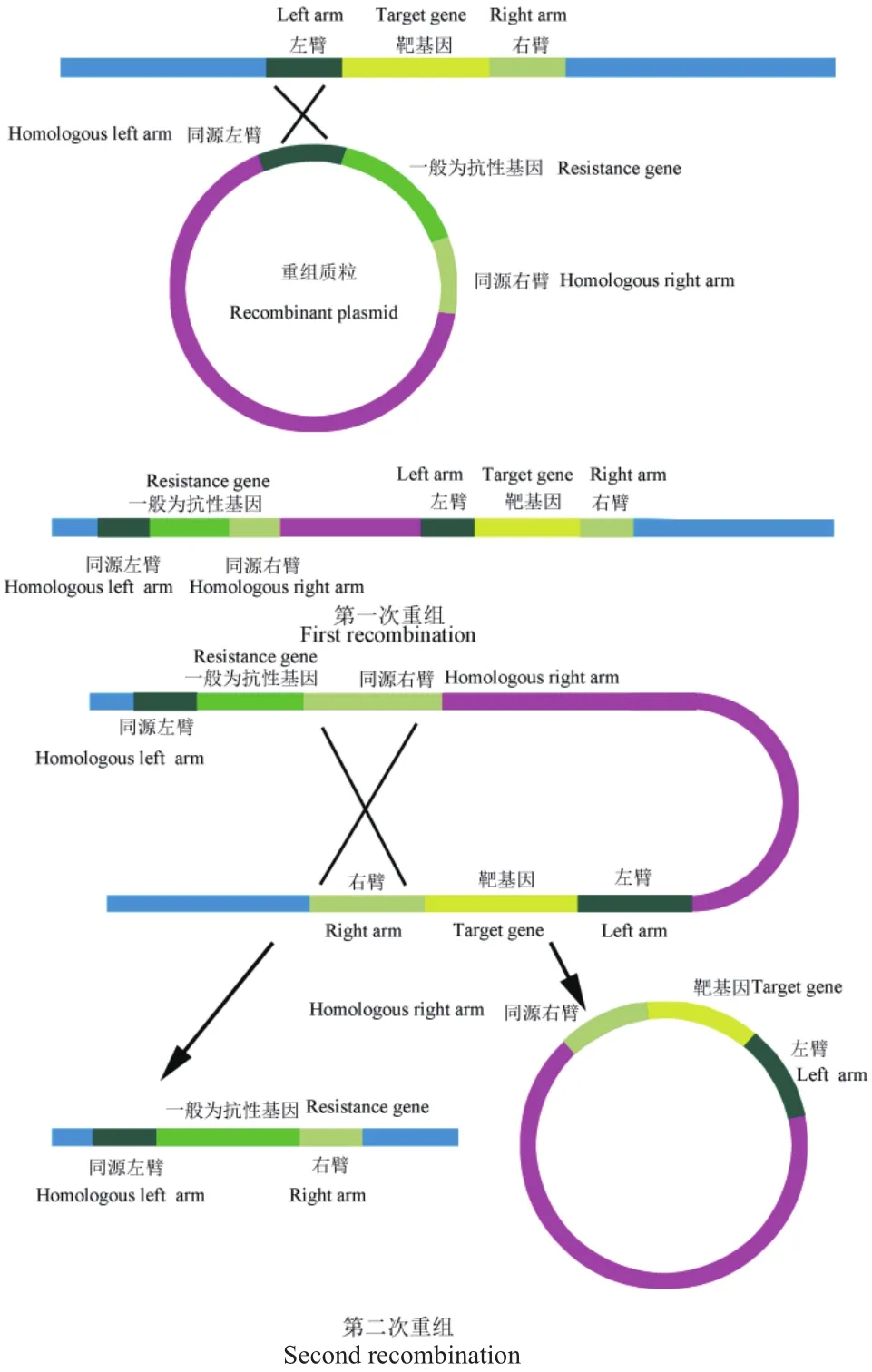
圖1 同源重組法基因敲除原理圖Fig.1 Schematic mechanisms of gene knockout using homologous recombination
2 利用蛋白質(zhì)工程提升堿性蛋白酶活性
在過(guò)去的30年中,蛋白質(zhì)工程已經(jīng)成為開(kāi)發(fā)提升酶活性的重要工具。當(dāng)前利用蛋白質(zhì)工程技術(shù)對(duì)堿性蛋白酶的改造主要集中在提高其熱穩(wěn)定性、抗氧化性以及專(zhuān)一性等的研究中。Jaouadi等[30]通過(guò)蛋白質(zhì)工程發(fā)現(xiàn)5個(gè)氨基酸:Leu31、Thr33、Asn99、Phe159和Gly182對(duì)短小芽孢桿菌B.pumilus 的SAPB的酶活性有重要影響,通過(guò)定點(diǎn)誘變構(gòu)建了12個(gè)突變體,結(jié)果發(fā)現(xiàn),三重突變體L31I/T33S/N99Y的比活最高,大約為野生型酶的2倍。Li等[31]通過(guò)對(duì)其N(xiāo)末端序列-1和-2處位置進(jìn)行定點(diǎn)誘變,以提高鏈霉菌角蛋白酶Sfp2的表達(dá)水平,最終發(fā)現(xiàn)L(-1)F突變體(48 935 U/mg)的比活性是野生型Sfp2的9倍。石亞偉等[32]利用蛋白質(zhì)工程技術(shù),對(duì)PB92枯草桿菌蛋白酶進(jìn)行改造,獲得A188P+V262I堿性蛋白酶突變體,相比野生型,其熱穩(wěn)定性以及耐堿性均有明顯提高,在兩種洗滌劑體系(STPP體系和MGDA體系)進(jìn)行評(píng)價(jià),發(fā)現(xiàn)在不加任何保護(hù)劑和穩(wěn)定劑的前提下,突變體相比親本酶具有更優(yōu)良的穩(wěn)定性和洗滌性能。蛋白質(zhì)工程技術(shù)出現(xiàn)至今已近40年,各項(xiàng)技術(shù)手段日漸成熟,結(jié)合大量的蛋白酶空間結(jié)構(gòu)的揭示,進(jìn)一步為利用在基因水平上通過(guò)氨基酸突變,提升蛋白酶的活性成為提升蛋白酶活力的重要手段。
3 通過(guò)優(yōu)化啟動(dòng)子或信號(hào)肽的方式提升堿性蛋白酶的表達(dá)量
啟動(dòng)子作為轉(zhuǎn)錄水平調(diào)控中的關(guān)鍵元件,而信號(hào)肽是翻譯階段的重要元件,在這二者的基礎(chǔ)上進(jìn)行改造,對(duì)于蛋白酶表達(dá)調(diào)控是一種很好的策略。如Liu等[33]使用最佳表達(dá)系統(tǒng)(信號(hào)肽DacBSP和雙啟動(dòng)子PBsamy-PBaamy)的重組枯草芽孢桿菌B.subtilis WB600在5 L發(fā)酵罐中培養(yǎng)以評(píng)估堿性蛋白酶的表達(dá)水平發(fā)現(xiàn),在56 h的峰值為27 860 U/mL。Guan等[34]通過(guò)應(yīng)用半理性啟動(dòng)子工程方法,構(gòu)建了包含單啟動(dòng)子和雙啟動(dòng)子的一系列表達(dá)質(zhì)粒,發(fā)現(xiàn)雙啟動(dòng)子PgsiB-PHpaII表現(xiàn)最佳。
啟動(dòng)子改造方面主要有通過(guò)啟動(dòng)子替換、串聯(lián)啟動(dòng)子或不同啟動(dòng)子組合調(diào)控蛋白酶的表達(dá),另外也可通過(guò)對(duì)啟動(dòng)子進(jìn)行定點(diǎn)突變來(lái)調(diào)控蛋白酶的表達(dá)。史超碩等[35]分析解淀粉芽孢桿菌的 α -淀粉酶啟動(dòng)子P1、枯草芽孢桿菌的 α-淀粉酶啟動(dòng)子P2及啟動(dòng)子組合P-1-2、P-2-1對(duì)克勞氏芽孢桿菌的堿性蛋白酶aprE表達(dá)的影響,結(jié)果表明,發(fā)酵48 h后,雙啟動(dòng)子重組菌枯草芽孢桿菌B.subtilis WB600/P-2-1-aprE表達(dá)的酶活性最高,達(dá)到了6 125 U/mL。楊春暉等[36]先通過(guò)基因克隆技術(shù)得到了短小芽孢桿菌B.pumilus的堿性蛋白酶基因的啟動(dòng)子片段,將其插入到穿梭質(zhì)粒載體pSUGV4中,分別轉(zhuǎn)入枯草芽孢桿菌B.subtilis和短小芽孢桿菌B.pumilus中進(jìn)行表達(dá),得到的堿性蛋白酶活性分別為466.5 U/mL和3 060 U/mL。
堿性蛋白酶生產(chǎn)菌株大多使用一般分泌(Sec)途徑將蛋白質(zhì)分泌到培養(yǎng)基中。所有分泌蛋白的一個(gè)共同特征就是它們的N端含有信號(hào)肽(signal peptide,SP),它們是之所以被分泌的重要識(shí)別位點(diǎn)。Degering等[37]構(gòu)建了由220種地衣芽孢桿菌B.licheniformis的信號(hào)肽(稱(chēng)為異源SP)和173種枯草芽孢桿菌B.subtilis的信號(hào)肽(稱(chēng)為同源SP)組成的信號(hào)肽文庫(kù)發(fā)現(xiàn),來(lái)源于地衣芽孢桿菌B.licheniformis幾丁質(zhì)酶的信號(hào)肽dBli00338可以使堿性蛋白酶在枯草芽孢桿菌B.subtilis TEB1020中活性提高6-7倍。
在啟動(dòng)子和信號(hào)肽這些調(diào)控元件水平上對(duì)微生物堿性蛋白酶進(jìn)行研究,也是目前在微生物堿性蛋白酶領(lǐng)域的熱門(mén)研究方向。通過(guò)單獨(dú)替換啟動(dòng)子、信號(hào)肽或?qū)烧哌M(jìn)行組合,比較分析不同啟動(dòng)子或信號(hào)肽對(duì)堿性蛋白酶的表達(dá)情況,選出最優(yōu)方案,從而提高了微生物堿性蛋白酶的酶活力。
4 展望
隨著對(duì)環(huán)境保護(hù)的日益重視,生物催化劑的使用越來(lái)越受到關(guān)注。微生物來(lái)源堿性蛋白酶在洗滌劑行業(yè)廣泛應(yīng)用是減少化學(xué)洗滌劑的不二選擇。目前堿性蛋白酶仍然不能滿足工業(yè)對(duì)多品種生產(chǎn)的增長(zhǎng)需求,特別是在液體洗滌市場(chǎng),我們需要面對(duì)的挑戰(zhàn)包括酶產(chǎn)量低、酶活力低、穩(wěn)定性差以及生產(chǎn)成本高等問(wèn)題。除此之外,還需對(duì)堿性蛋白酶生產(chǎn)過(guò)程精確控制,特別是簡(jiǎn)化堿性蛋白酶的下游純化工藝尤為重要。近年來(lái)合成生物學(xué)成為學(xué)科發(fā)展的一個(gè)新的熱點(diǎn),通過(guò)人工染色體構(gòu)建、地盤(pán)細(xì)胞設(shè)計(jì)、全局轉(zhuǎn)錄調(diào)控等有望應(yīng)用于堿性蛋白酶生產(chǎn)相關(guān)的多基因表達(dá)的調(diào)控[38],加之開(kāi)發(fā)更多便捷高效的芽孢桿菌基因編輯技術(shù)等有望獲得人們理想的蛋白酶。但蛋白酶產(chǎn)品的實(shí)現(xiàn),在酶制劑的發(fā)酵、分離純化、穩(wěn)定性方面仍然有賴于經(jīng)典的蛋白類(lèi)產(chǎn)品的發(fā)酵和提純工藝的進(jìn)步[39]。如何保持酶的高濃度、高純度、高穩(wěn)定性等,特別是在洗滌劑中的穩(wěn)定,都是堿性蛋白酶活力提升繞不開(kāi)的技術(shù)難題。不論采用何種途徑提高堿性蛋白酶的酶活力,都有望擴(kuò)大其在生物技術(shù)領(lǐng)域和工業(yè)領(lǐng)域中的應(yīng)用。
