發酵劑在乳品中應用的影響因素及協同發酵的研究進展
2021-06-22 01:09:18侯團偉段劍平吳曉紅
食品工業科技 2021年1期
關鍵詞:乳制品
侯團偉,段劍平,吳曉紅
(牧堡(上海)食品科技有限公司,上海201103)
乳品發酵劑是指在適宜的條件下,通過代謝作用產生乳酸、多肽、胞外多糖等物質的一類有益微生物,常稱作乳酸菌。乳酸菌通過自身酶系統的代謝作用,可將乳中乳糖轉化為乳酸,蛋白質降解為多肽或氨基酸,同時會伴隨有機酸、醇、酯、酮等風味成分的產生,進而賦予乳品特定的風味及豐富的營養物質,因而被廣泛應用于乳制品工業領域,例如酸奶、發酵黃油、發酵奶油、酪乳、干酪等食品[1-4]。乳酸菌的發酵特性對發酵乳制品品質的影響至關重要,而更加深入地了解乳酸菌發酵的影響因素及協同發酵作用,對于改善產品品質,滿足消費者的需求十分重要。
發酵乳品生產過程中,使用單一的發酵劑發酵,通常會存在一些缺陷[5-7]:如植物乳桿菌可以提升產品的風味及保存時間,然而其卻存在乳中生長不佳的問題;雙歧桿菌除了可以產生乳酸外,具有諸多益生作用,但其屬于嚴格厭氧型乳酸菌使其發酵受到一定限制;嗜熱鏈球菌單獨使用雖然可滿足產酸的需求,但是風味單一,難以滿足風味需求;同時,單一發酵劑的使用對抗生素、噬菌體的抵抗能力有限,發酵失敗風險較高。而發酵劑協同發酵的使用可以克服此類缺點,提升產品品質,降低發酵風險。因此,本文從發酵劑的種類及益生作用、影響發酵的因素與協同發酵作用三個方面對乳制品發酵劑進行概述,以期對發酵乳制品生產提供一定的指導。
1 乳制品發酵劑的種類與益生作用
1.1 發酵劑的種類
目前,我國批準可用于食品的菌種有35種。常用的乳酸菌主要來自乳桿菌屬(Lactobacillus)、鏈球菌屬(Strptococcus)、雙歧桿菌屬(Bifidobacterium)、明串珠菌屬(Leuconostoc)及乳球菌屬(Lactococcus)5個屬[8-9]。
其中,乳桿菌屬有202個種,常用作乳制品發酵劑的有8種,包括:嗜酸乳桿菌(L.acidophilus)、干酪乳桿菌(L.casei)、德氏乳桿菌(L.delbrueckii)、瑞士乳桿菌(L.helveticus)、約氏乳桿菌(L.johnsonii)、副干酪乳桿菌(L.paracasei)、鼠李糖乳桿菌(L.rhamnosus)及保加利亞乳桿菌(L.bulgaricus);此類乳酸菌主要來自人體消化道(腸道),主要為同型發酵型,以產生L-型乳酸為主,一般為嗜溫乳酸菌。鏈球菌屬有104個種,常用乳酸菌為嗜熱鏈球菌(S.thermophilus),其常見于乳品中,同型發酵,產生L-型乳酸,其具有較高的溫度耐受性。雙歧桿菌屬有64個種,常用乳酸菌為長雙歧桿菌(B.longum)、短雙歧桿菌(B.breve)、青春雙歧桿菌(B.adolescentis)、雙歧雙歧桿菌(B.bifidum)及嬰兒雙歧桿菌(B.infantis)[9];雙歧桿菌是一類革蘭氏陽性、厭氧型,一般存在于哺乳動物及某些昆蟲的胃腸道之中[6]。除來自昆蟲的雙歧桿菌,其他的雙歧桿菌不具有呼吸作用途徑,僅能通過發酵產生能量。大多數腸道微生物通過糖酵解途徑或戊糖磷酸途徑代謝碳水化合物,而雙歧桿菌則通過果糖-6-磷酸磷酸酮酶途徑進行碳水化合物代謝,也稱雙歧途徑[10]。明串珠菌屬有14個種,常用于發酵乳制品的包括乳酸明串珠菌(L.lactis)、腸膜明串珠菌(L.mesenteroides)、酒明串珠菌(L.oeni)及乳脂明串珠菌(L.cremoris),其是一類革蘭氏陽性菌,兼性厭氧型,異型發酵,主要為嗜溫乳酸菌[11-12]。乳球菌屬有15個種,常用乳制品中的為乳酸乳球菌,同型發酵,產生L-型乳酸,為嗜溫乳酸菌。常見發酵乳品中乳酸菌的生化及代謝特性見表1[13-22]。
1.2 乳酸菌的益生作用
幾乎所有發酵劑均具有益生作用,乳酸菌可通過在人體消化道中定植或其發酵制品中含有某些代謝產物來間接發揮其益生作用。
1.2.1 可定植于腸道的乳酸菌 雙歧桿菌、干酪乳桿菌、保加利亞乳桿菌、瑞士乳桿菌、嗜酸乳桿菌、鼠李糖乳桿菌及嗜熱鏈球菌通常定植于人體腸道中,通過生長繁殖(營養競爭)及代謝產物(活性成分)起到調節腸道菌群及促進人體健康的作用。
雙歧桿菌在體內的變化:新生嬰兒腸道內是無菌的,但嬰兒出生后很多細菌開始繁殖;1~2 d,可在糞便中檢測出腸球菌、大腸桿菌、梭菌及乳酸菌;3~4 d,雙歧桿菌開始繁殖,第5 d,雙歧桿菌成為優勢菌群[23];采用母乳喂養的嬰兒,腸道內的雙歧桿菌數量高于人工喂養嬰兒,且前者中的雙歧桿菌數量占總菌數的91%;人體內的雙歧桿菌的數量將隨年齡的增長而減少,且種類由短雙歧桿菌及嬰兒雙歧桿菌為主轉變成以長雙歧桿菌及青春雙歧桿菌為主要菌群[22,24-25]。雙歧桿菌已被證實具有多種益生作用,如抑菌、防止心血管疾病、提升機體免疫力等作用。雙歧桿菌主要通過產生抑菌物質(有機酸、細菌素、類細菌素等)、競爭營養和粘附位點等作用,起到抑菌作用[26-27];目前,雙歧桿菌降低血液膽固醇的假說主要有:a其代謝產物丙酸等有機酸抑制了膽固醇的合成;b酸性條件下,膽固醇與游離膽鹽發生沉降作用;c雙歧桿菌對膽固醇的吸附作用[22,28]。雙歧桿菌主要通過激活腸道粘膜的免疫系統,使機體產生細胞因子、抗體,進而提升免疫力,Francesca等[29]研究發現,雙歧桿菌可通過誘生細胞因子,促進免疫B細胞的增殖來調節免疫力;Wang等[30]發現,動物雙歧桿菌通過激活小鼠巨噬細胞,增強IL-1活性,同時產生IL-6。
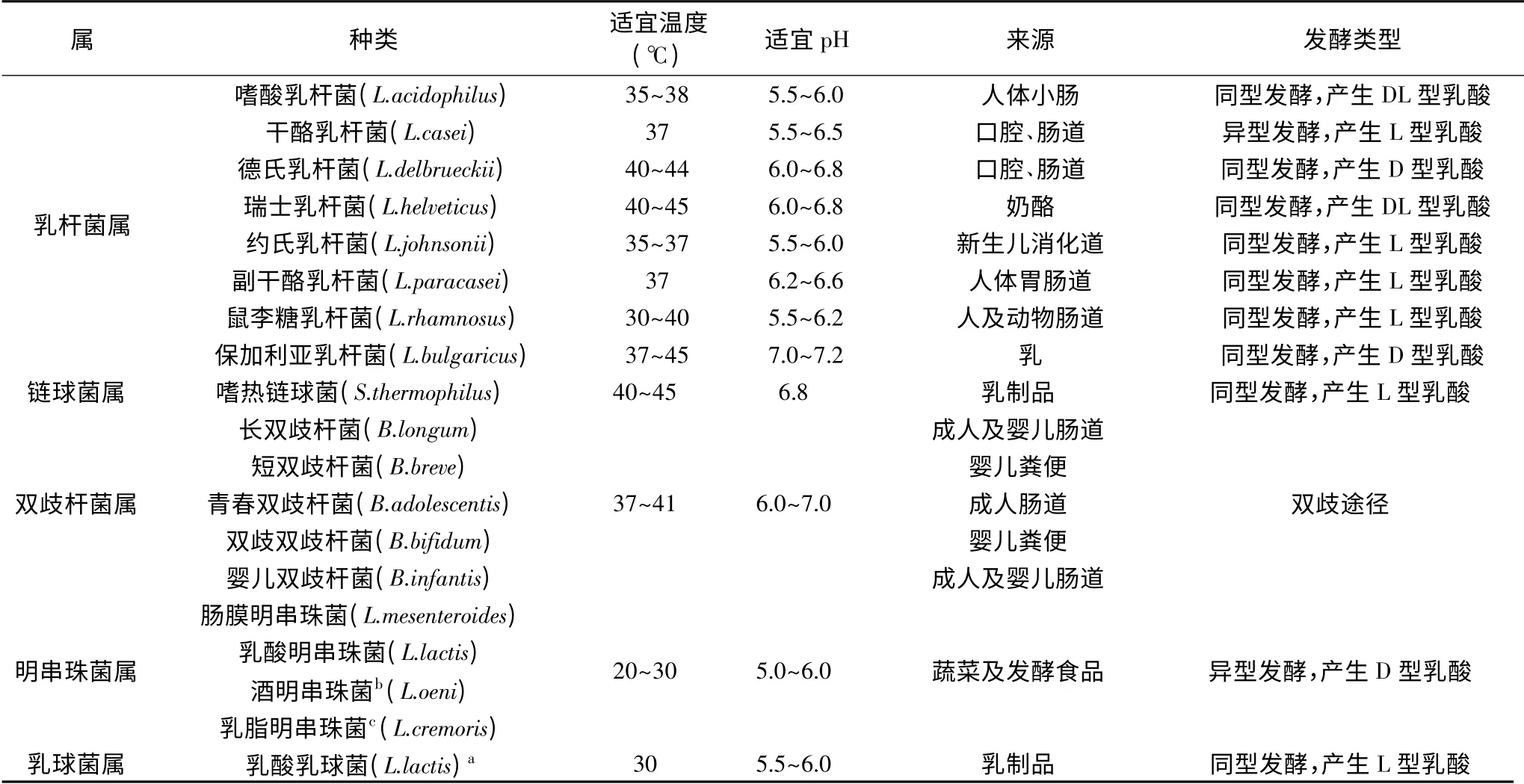
表1 常用于發酵乳品中乳酸菌的種類及特性Table 1 Types and metabolic characteristics of lactic acid bacteria in fermented dairy
常用的乳桿菌屬乳酸菌:干酪乳桿菌存在于干酪、牛奶及人的口腔、腸道中,屬于人體原籍菌[31],研究證明,其除了具有抑菌作用、提高免疫力及抗腫瘤外,還具有降低血壓的作用,這主要是通過其細胞外蛋白酶的作用,產生的肽類物質對血管緊張素抑制來實現降血壓作用,目前,其主要應用于酸乳、干酪及乳酸菌飲料的生產[16,32];1905年,保加利亞乳桿菌是科學家第一次從酸奶中分離,其具有調節腸道菌群平衡、提高免疫力等作用,主要用于酸奶、發酵奶油及干酪的生產[33];瑞士乳桿菌除具有抑菌、調節機體免疫力等作用外,還具有優于其他乳桿菌的抗高血壓的作用,其主要通過產生活性酶(ACE)、抗高血壓肽Val-Pro-Pro及Ile-Pro-Pro等物質,抑制血管緊張素轉化加以實現[34-36];嗜酸乳桿菌存在于人體口腔、腸道中,具有一般乳桿菌的益生作用,主要用于普通酸奶的生產;鼠李糖乳桿菌存在于人及動物腸道內,不能利用乳糖,可以發酵麥芽糖、阿拉伯糖等;其除具有一般乳桿菌益生作用外,還具有預防幽門螺桿菌定植的作用[37],主要用于酸奶制作。
嗜熱鏈球菌存在于人體及乳制品中,其產生的細胞外多糖具有促進雙歧桿菌增殖及調節免疫力的功效,其主要用于硬質干酪及酸奶的生產[38-39]。
1.2.2 不可定植于腸道的乳酸菌 乳酸乳球菌及明串珠菌主要通過代謝產物促進其他益生菌生長間接發揮益生作用。
乳酸乳球菌是一種革蘭氏陽性菌,兼性厭氧型,同型發酵;不能在胃腸道定植,但其可產生乳酸及細菌素(多肽或蛋白)抑制其他有害微生物的生長,進而促進其他有益發酵菌的生長,其主要用于干酪、酪乳、酸奶油生產[40-41]。
明串珠菌是一類革蘭氏陽性菌,兼性厭氧型,異型發酵;存在于發酵食品中,其不能在腸道內定植,僅能通過代謝產生的有益成分(α-葡萄糖-低聚糖)促進其他有益菌的生長,其主要用于生產干酪(如埃門塔爾干酪、高達干酪等)、黃油、酪乳、酸奶油[11-12]。
乳品發酵劑通常具有益生功能,但乳酸菌的益生作用一般認為,只有當消費的食品中存活的益生菌數達到106CFU/g以上[42]才能實現;由于干酪在酸性條件下具有緩沖作用,在消化道及胃中對益生菌具有保護而被認為是益生菌最為有效的載體;而在不影響干酪品質的條件下,如何保證益生菌在干酪制作過程中的存活量將是益生菌干酪制作的另一個挑戰,益生菌干酪制作過程中常見問題及可能的解決方案見表2[42-44]。
2 乳制品發酵的影響因素
乳制品的發酵受多種因素的影響,如原料乳、培養條件、發酵菌種、噬菌體、抗生素等,原料乳需要保證有足夠的乳糖、蛋白質等成分可供乳酸菌發酵使用;培養條件,即需要確保適宜的發酵溫度及pH;前兩者相對較易實現,而發酵菌種的選用不當、噬菌體及抗生素的污染將大大增加乳制品發酵失敗的風險。
2.1 發酵菌種的選擇
發酵底物的不同,需要選擇適宜的發酵菌種或混合菌種,否則將會導致發酵失敗或得不到滿意的發酵效果。不同發酵乳制品中,適宜選用的常用乳酸菌見表3[16,45-47]。
干酪作為發酵乳制品占比較高的一種,世界產量約1900萬噸每年(約占牛奶總產量的35%)。干酪也是發酵乳制品中最為復雜的,存在著復雜的微生物關系。如何進行菌種的有效結合使用將直接決定干酪的品質。依據乳酸菌在干酪的制作過程中的主要作用,可將其分為發酵乳酸菌及非發酵乳酸菌;發酵乳酸菌主要是產酸以達到凝乳的目的,而非發酵乳酸菌則主要作用于干酪的成熟期,可賦予奶酪特殊的風味及質構。干酪發酵及熟化過程中,發酵乳酸菌與非發酵乳酸菌相互作用見圖3[48]。
最常用的嗜溫發酵乳酸菌包括明串珠菌、乳酸乳球菌及乳脂鏈球菌;嗜熱鏈球菌為嗜熱乳酸菌中常用于發酵制作硬質及半硬質干酪的菌種,如瑞士及意大利干酪,乳酸菌通過糖酵解產生乳酸,使得體系酸度降低,在實現凝乳的同時,也抑制了致病菌的生長;德氏乳桿菌及瑞士乳桿菌在干酪生產中被廣泛用作產生特殊風味的附屬發酵劑,干酪乳桿菌、植物乳桿菌、明串珠菌可以通過檸檬酸鹽代謝為干酪貢獻風味成分[48-51]。

表2 益生菌干酪制作過程中常見問題Table 2 The common problems in process of probiotic cheese

表3 常用于發酵乳制品中的乳酸菌種類Table 3 The types of lactic acid bacteria commonly used in fermented dairy products
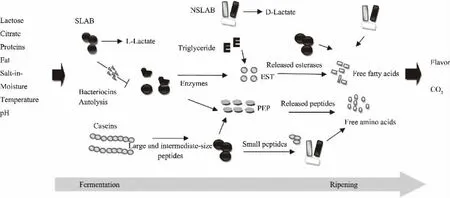
圖1 干酪加工過程中,發酵乳酸菌與非發酵乳酸菌的相互作用Fig.1 The interaction between SLAB and NSLAB during the process of cheese
Lynch等[52]研究表明,副干酪乳桿菌與植物乳桿菌的添加可以促進切達干酪風味的形成,進而縮短了成熟期。Daniel等[53]在瑞士干酪的生產加工中,通過添加干酪乳桿菌來抑制丙酸菌的生長,減少產氣過多,對干酪品質的影響。黃宜等[54]通過研究得出,將乳酸乳球菌與腸膜明串珠菌用于制作高達干酪模型,可以產生良好的風味,具備制作高達干酪潛力。
結合具體發酵乳制品底物及發酵劑特點(產酸或產風味成分),選擇適宜的菌種或巧妙地使用菌種組合發酵才能實現發酵目的。
2.2 噬菌體
噬菌體是一類侵害細菌的病毒的統稱,其只有存在于細菌細胞內才能繁殖,且宿主的專一性強。對于發酵乳制品而言,若被噬菌體污染,將會導致發酵時間延長,或者造成菌體裂解,導致發酵失敗,引起產品風味及口感變差的后果[55]。據統計,60%~70%奶酪生產的失敗是由噬菌體污染所導致。
噬菌體通常分為兩種,即溫和噬菌體及烈性噬菌體,前者可將自身DNA整合到相應宿主染色體上且可以穩定遺傳;后者感染后,迅速在宿主體內繁殖并最后釋放出子代噬菌體,再去感染其他細胞[55]。研究發現,噬菌體從侵染乳酸菌到釋放新的噬菌體,該周期僅需40~45 min,且數量由1增殖到100或300,乳酸桿菌及乳球菌被噬菌體感染后,在感染初期,活菌數目將降至低于感染前的十分之一,感染25 d后,活菌數量將低于感染前的千分之一[55-56]。馬成杰等[57]研究了嗜熱鏈球菌噬菌體對酸奶發酵的影響,結果顯示,噬菌體的存在不僅影響產酸速率,還會導致產品口感變差、后酸化嚴重和乳清析出等問題。Silvina等[58]從有質量缺陷的藍紋干酪中分離出了9種明串珠菌噬菌體,研究發現,此類噬菌體具有較高的耐熱性(80℃,30 min或90℃,2 min才能滅活)及乙醇耐受性(75%乙醇濃度,30 min才能滅活)。因此為確保發酵劑安全,通常牛奶消毒條件為85℃下,加熱20~30 min。
學者們對乳酸菌噬菌體基因組及與宿主的相互作用機理進行了研究,常見的乳酸菌及相應噬菌體的特異性結合方式見表4。
通過上述乳酸菌噬菌體的研究發現,乳酸菌若被噬菌體感染,則需要通過與之對應的特異性的結合物質才能夠實現,因此,一方面可以借助噬菌體的特異性,采用乳酸菌混合或輪換發酵,降低由于噬菌體侵染帶來的發酵失敗的風險,此菌種需要滿足對相同噬菌體的侵染敏感度不同及具有相似的發酵條件和速率;另一方面也可采用抗噬菌體菌株的選育(借助自然突變、物理誘導及化學誘變的方法)為阻斷其宿主菌的裂解作用提供可能,從菌種選育的角度防御噬菌體對發酵劑的影響。

表4 常見的乳酸菌及相應噬菌體的特異性結合方式Table 4 The specific binding of common lactic acid bacteria and the corresponding phage

表5 常見乳酸菌及耐抗生素相關基因Table 5 Common lactic acid bacteria and antibiotic-resistant genes
趙慧穎[66]從異常發酵液中分離選育了植物乳桿菌抗噬菌體菌株并對噬菌體滅活進行了研究,研究表明抗噬菌體菌株產酸及生長狀況與原菌株無顯著差異,且其可通過抑制噬菌體的吸附或減少了噬菌體可利用的營養物質(ATP、嘧啶)來抑制噬菌體遺傳物質的復制來實現抗噬菌體作用。王慕華等[67]研究了制備保加利亞乳桿菌與嗜熱鏈球菌抗噬菌體菌株的原生質體融合,結果表明融合后的各項指標優良,可適用酸奶生產。
此外,由于噬菌體廣泛存在于發酵設備、車間等環境中,應做好設備的消毒及操作人員、車間衛生工作。
2.3 抗生素
1929年第一種抗生素青霉素被發現以來,目前已知的抗生素達9000多種。在牛奶生產中,抗生素是一種治療乳腺炎的有效方法,但牛奶中抗生素的殘留將可能導致發酵的延遲或失敗。
為更好地認知乳酸菌耐藥性,研究者對其可能存在的耐藥性基因進行了研究,常見乳酸菌種與耐藥性相關基因見表5[68-70]。一種耐藥基因可能具有多重抗生素耐受性。
乳酸菌種類不同,對抗生素種類呈現不同的耐受性。李平蘭等[71]以嗜酸乳桿菌、長雙歧桿菌、干酪乳桿菌、植物乳桿菌及腸球菌為實驗乳酸菌,研究了8類抗生素對其生長的影響,結果表明:上述乳酸菌對大環內酯類抗生素、呋喃妥因、氯霉素及利福平抗生素敏感,對其他抗生素則呈現出不同的敏感度。潘琳等[72]對乳球菌屬對抗生素的最小抑制量進行了研究分析,結果表明:明串珠菌對福利平、紅霉素及萬古霉素敏感,最小抑制量為1.25μg/mL,對鏈霉素、新霉素最大耐受量為1280μg/mL;嗜熱鏈球菌對紅霉素、福利平、四環素敏感,最小抑制量為0.625μg/mL,對卡那霉素、鏈霉素的最大耐受量為160和80μg/mL;乳球菌對紅霉素、福利平及四環素敏感,最小抑制量為<0.625μg/mL,對新霉素最大耐受量可達640μg/mL。通過認知乳酸菌的抗生素敏感度,進行混合發酵,能夠降低發酵失敗的風險。
因此,為保證發酵乳制品的成功發酵,一方面可以通過降低乳中抗生素的使用及強化管理,另一方面可以通過篩選具有較好存活性的乳酸菌或進行不同抗生素敏感程度的乳酸菌組合來實現發酵目的。
3 發酵劑間的協同發酵
微生物群體間的生長關系包括共生關系、競爭關系、偏利關系、偏害關系等[5]。協同發酵即是利用微生物間的互利共生關系、偏利關系達到更好的發酵效果。
3.1 共生關系協同發酵
保加利亞乳桿菌與嗜熱鏈球菌作為發酵劑被廣泛地用于發酵乳制品中。兩種菌種對乳糖的代謝方式均為同型乳酸發酵[73]。大量研究表明,二者存在互惠共生關系。田輝等[39]認為,嗜熱鏈球菌在發酵牛奶時,由于產酸快的特點,創造了一個適宜另一種乳酸菌生長的環境;李志成等[74]研究表明,嗜熱鏈球菌與保加利亞乳桿菌的混合發酵時產酸及產生黏性的能力均優于單一菌種;一些學者認為它們相互作用的機制是兩種菌種代謝產物相互促進的結果,即在發酵過程中,保加利亞乳桿菌不含有甲酸及丙酮酸裂解酶,進而導致生長缺乏嘌呤及甲酸,而嗜熱鏈球菌可以通過糖酵解等途徑為其提供需要的甲酸、葉酸及丙酮酸等進而促進保加利亞乳桿菌的生長;保加利亞乳桿菌產生的氨基酸(如甲硫氨酸、脯氨酸、組氨酸)及多肽類的物質則促進了嗜熱鏈球菌的生長,相互作用關系見圖2[75-78]。
但Mchiouer等[79]研究認為,保加利亞乳桿菌與嗜熱鏈球菌混合發酵的產酸效果((101±3.2)°D)優于單獨發酵產酸((92±2.7)°D);且保加利亞乳桿菌的發酵濾液對嗜熱鏈球菌產酸具有較好的促進作用,而嗜熱鏈球菌的發酵濾液對保加利亞乳桿菌的產酸無促進效果。
盧兆蕓等[80]將嗜熱鏈球菌與保加利亞乳桿菌用于酸奶發酵,成功保留純酸牛奶的口感與芳香,同時賦予了產品紫紅薯的色澤與營養;胡穎等[81]將嗜熱鏈球菌與保加利亞乳桿菌混合發酵制作出了濃郁奶香味及酸味清爽的夸克產品。
3.2 偏利關系協同發酵
3.2.1 嗜熱鏈球菌與乳酸乳球菌 乳酸乳球菌及嗜熱鏈球菌均屬于革蘭氏陽性菌,兼性厭氧型,同型發酵。文獻報道,乳酸乳球菌除產生乳酸抑制其他微生物生長外,還可通過核糖體產生一類生物活性的多肽、蛋白抑菌物質,此類抑菌素具有良好的酸、熱穩定性,對革蘭氏陽性及陰性菌均有較強的抑制作用[82-83],因此二者共存時可能會帶來負面關系。
然而,Claude等[84]認為乳酸乳球菌與嗜熱鏈球菌間存在偏利共生關系。其模擬在切達干酪的生產的條件下,研究了單一(嗜熱鏈球菌或乳酸乳球菌)或混合菌種(嗜熱鏈球菌與乳酸乳球菌)的生長狀況,研究結果表明,嗜熱鏈球菌的存在可以較好地促進乳酸乳球菌的生長,但是乳酸乳球菌的存在對嗜熱鏈球菌的生長無顯著影響。Pega等[85]采用嗜熱鏈球菌及乳酸乳球菌作為發酵劑,成功制作了含有生育酚及植物甾醇生物活性成分的干酪。乳酸乳球菌及嗜熱鏈球菌具體相互作用的機理還有待進一步闡明。
3.2.2 植物乳桿菌與瑞士乳桿菌 植物乳桿菌是一類在植物、土壤、發酵食品中發現的革蘭氏陽性菌,兼性厭氧型乳酸菌。Ralitsa等[86]研究發現,分離的8株植物乳桿菌具有高活性氨基肽酶,但是水解蛋白能力卻非常有限;另有學者研究發現,植物乳桿菌中存在肽轉運系統,但缺少編碼細胞壁蛋白酶系統,即單獨存在可以通過寡肽及二三肽運輸體實現胞外肽的轉運,在胞內多肽酶的作用下,實現肽的進一步降解[87]。
瑞士乳桿菌由于可以降解苦味肽,同時可產生獨特的風味,因此常用于干酪、發酵乳等乳制品中。瑞士乳桿菌屬于多氨基酸營養缺陷型乳酸菌,但其卻能在游離氨基酸稀少的乳中良好生殖;這主要是由于瑞士乳桿菌中,編碼細胞壁蛋白酶的基因PrtM存在兩種,其通常含有兩種細胞壁蛋白酶,因此使得其具有強大的蛋白質水解能力[88]。
通過植物乳桿菌與瑞士乳桿菌的協同發酵,可以 獲 得 良 好 的 產 品。Carina等[89]研 究 表 明,L.plantarum I91可利用L.helveticus 138降解干酪的產物生長,同時研究發現L.plantarum I91對蛋白質的初級水解無作用,而對次級水解有影響。施小玉[90]將植物乳桿菌、瑞士乳桿菌、發酵乳桿菌、干酪乳桿菌及乳酸克魯維酵母用于制作出了風味獨特且自由基清除率優良的功能性羊乳開菲爾。姜梅等[91]借助植物乳桿菌和瑞士乳桿菌混合發酵,獲得了特色雙菌雙蛋白發酵乳。
3.2.3 雙歧桿菌與嗜酸乳桿菌 雙歧桿菌中含有多種代謝相關的酶類[92-93]:a.糖代謝相關酶類:雙歧桿菌中均含有活力高于其他腸道菌的α、β-半乳糖苷酶,其中β-半乳糖苷酶為乳酸菌種常見乳糖酶,而較少見的α-半乳糖苷酶則可降解α-D-半乳糖苷寡糖;此外,雙歧桿菌中還有α-葡萄糖苷酶、甘露糖苷酶、D-木糖苷酶、β-呋喃果糖苷酶等水解寡糖的酶;b.蛋白代謝相關酶:磷蛋白磷酸酶、三肽酶、二肽酶、羧肽酶等。這也使得雙歧桿菌具有較強的生存能力。嗜酸乳桿菌耐酸性強,可利用乳糖、果糖、蔗糖及葡萄糖同型發酵產生乳酸,但其分解蛋白能力弱,單獨發酵,產酸緩慢。
利用雙歧桿菌與嗜酸乳桿菌存在偏利共生關系進行協同發酵,可以提升產品生產效率及提升產品品質。張香美等[94]研究了短雙歧桿菌與嗜酸乳桿菌混合培養時微生態關系,結果表明混合培養加快了產酸,嗜酸乳桿菌對短雙歧桿菌無明顯的促進或抑制作用,而短雙歧桿菌則促進了嗜酸乳桿菌的生長,這可能由于嗜酸乳桿菌在發酵過程中需要維生素B2、葉酸、醋酸鹽等生長因子,而雙歧桿菌在生長中可產生醋酸、葉酸、維生素B2等多種維生素,進而對其產生促進作用[95-96]。曲秀偉[97]研究雙歧桿菌和嗜酸乳桿菌對干酪產生抗氧化肽及穩定性的研究表明,當二者以1∶1比例混合培養時,產酸加速且活菌數顯著高于單獨培養,并成功制作了具有抗氧化作用的益生菌干酪。田其英等[98]采用雙歧桿菌與嗜酸乳桿菌混合發酵成功制備了口感細膩、奶香純厚、高活菌數的凝固型發酵乳。
4 展望
隨著生物技術的不斷發展,將有更多益生性發酵微生物被分離及進行安全性評估。影響發酵的因素中,具有良好噬菌體耐受性及可賦予產品良好品質的乳酸菌的篩選與組合需要系統研究。此外,發酵劑的抗生素耐受性基因是否存在安全風險也值得關注與研究。單獨使用一種發酵劑難以獲得滿意的發酵效果,利用具有共生、偏利關系及對噬菌體或抗生素的敏感程度不同的乳酸菌進行科學的組合使用,可以實現不同發酵劑的優勢互補,提高發酵效率,提升發酵乳制品品質,降低發酵失敗的風險,同時某些乳酸菌的相互作用機制仍需進一步的闡明。
猜你喜歡
今日畜牧獸醫(2021年9期)2021-03-28 16:15:36
中國乳業(2018年3期)2018-04-13 01:05:08
食品與機械(2017年4期)2017-07-05 14:46:17
農產品市場周刊(2017年16期)2017-05-27 12:04:19
中國乳業(2017年2期)2017-01-18 14:37:36
中國乳業(2016年4期)2016-11-07 09:50:28
現代食品(2016年24期)2016-04-28 08:12:06
工業設計(2016年12期)2016-04-16 02:51:53
食品科學(2013年14期)2013-03-11 18:25:10
食品安全導刊(2011年6期)2011-04-12 00:00:00
